
Mlig-SKP1 Gene Is Required for Spermatogenesis in the Flatworm Macrostomum lignano
 , and
, and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Knockdown of Mlig-SKP1 Gene Does Not Affect Regeneration but Causes Abnormal Testes Morphology
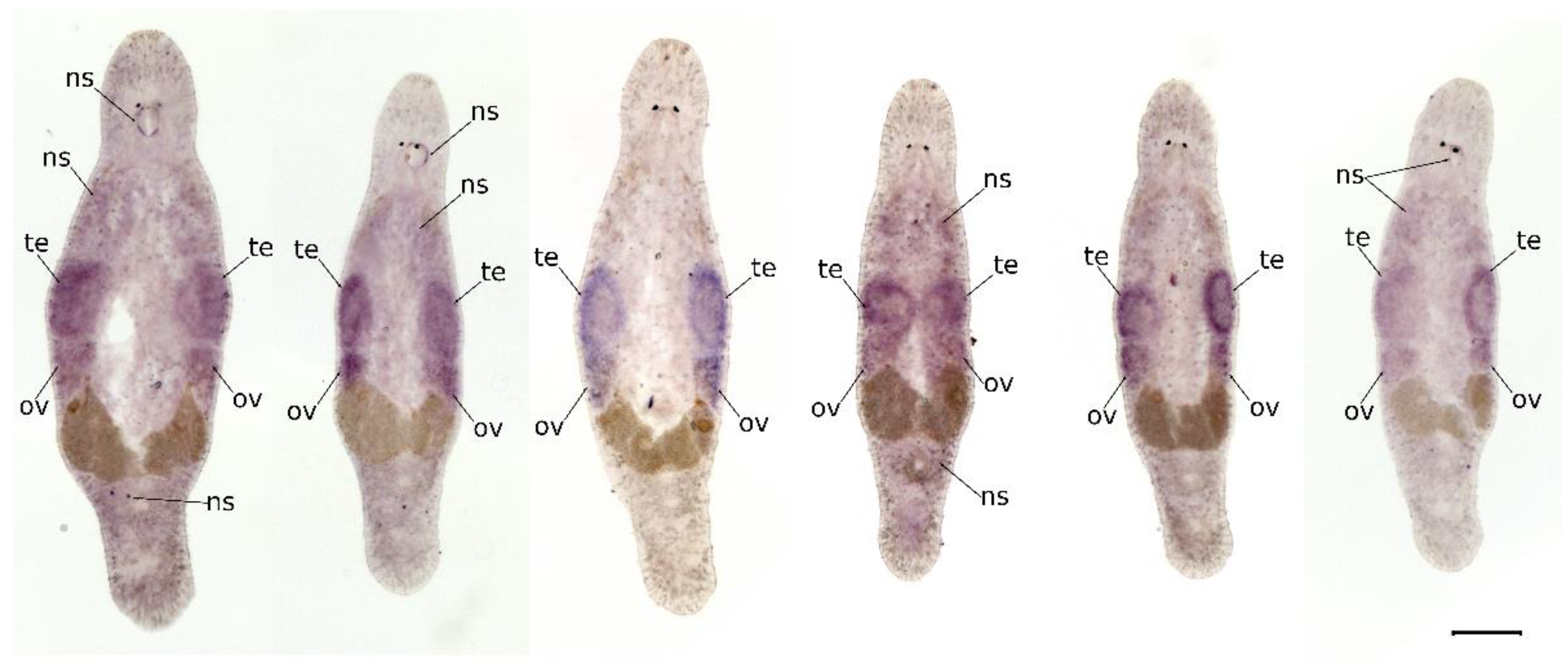
2.2. Mlig-SKP1 Is Expressed in Gonads
2.3. Mlig-SKP1 Knockdown Leads to Decreased Fertility
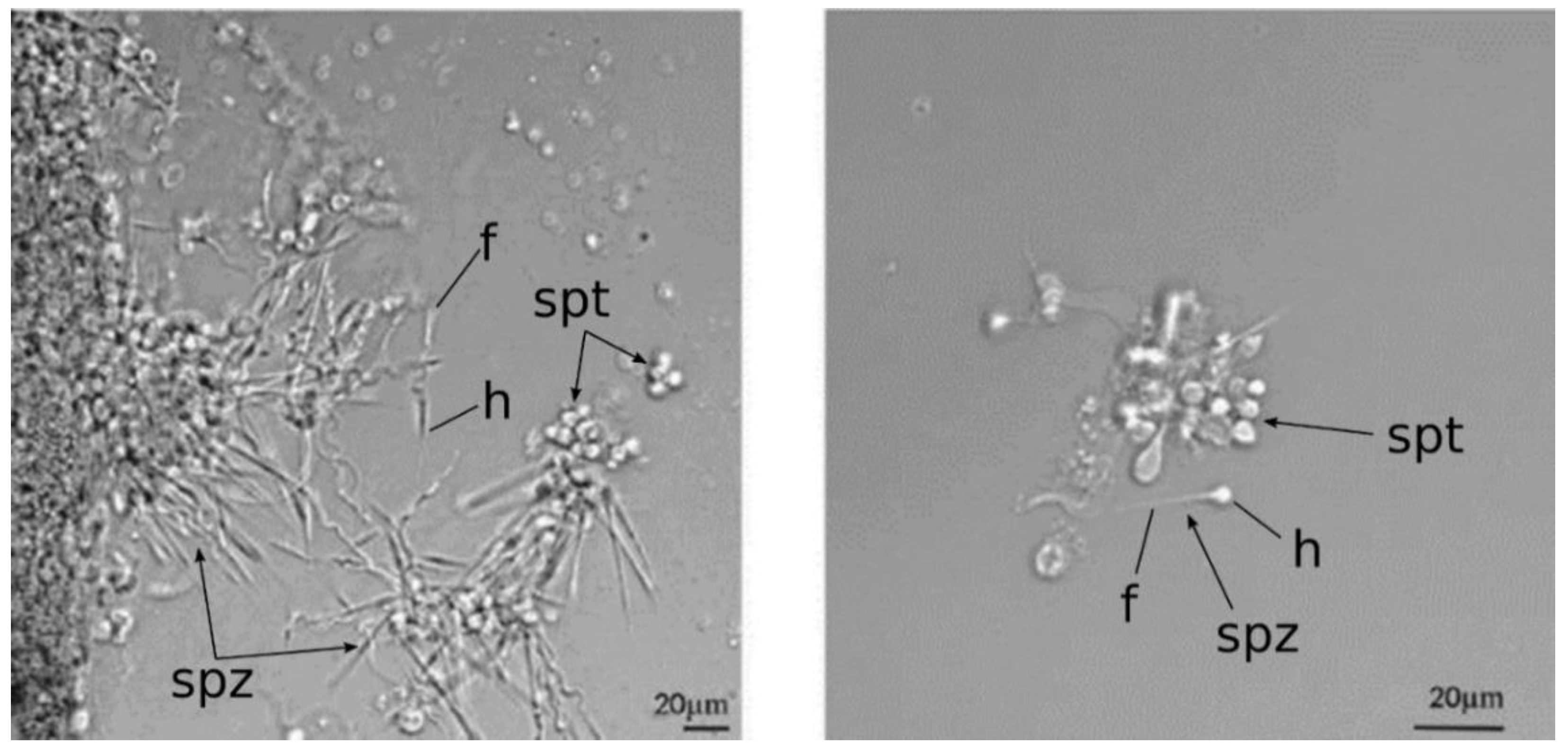
2.4. Mlig-SKP1 Knockdown Results in Aberrant Morphology of Sperm Cells
2.5. Conserved Role of SKP1 in Regulation of Spermatogenesis
3. Materials and Methods
3.1. Organism and Its Culture Conditions
3.2. RNA Interference
3.3. Whole-Mount In Situ Hybridization
3.4. RNAi Fertility Experiments
3.5. qRT-PCR for Knock Down Verification
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wudarski, J.; Egger, B.; Ramm, S.A.; Schärer, L.; Ladurner, P.; Zadesenets, K.S.; Rubtsov, N.B.; Mouton, S.; Berezikov, E. The free-living flatworm Macrostomum lignano. EvoDevo 2020, 11, 5. [Google Scholar] [CrossRef] [PubMed]
- Schärer, L.; Ladurner, P. Phenotypically plastic adjustment of sex allocation in a simultaneous hermaphrodite. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270, 935–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Mulder, K.; Pfister, D.; Kuales, G.; Egger, B.; Salvenmoser, W.; Willems, M.; Steger, J.; Fauster, K.; Micura, R.; Borgonie, G.; et al. Stem cells are differentially regulated during development, regeneration and homeostasis in flatworms. Dev. Biol. 2009, 334, 198–212. [Google Scholar] [CrossRef] [PubMed]
- Kuales, G.; De Mulder, K.; Glashauser, J.; Salvenmoser, W.; Takashima, S.; Hartenstein, V.; Berezikov, E.; Salzburger, W.; Ladurner, P. Boule-like genes regulate male and female gametogenesis in the flatworm Macrostomum lignano. Dev. Biol. 2011, 357, 117–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladurnera, P.; Riegera, R.; Baguñàb, J. Spatial Distribution and Differentiation Potential of Stem Cells in Hatchlings and Adults in the Marine Platyhelminth Macrostomum sp.: A Bromodeoxyuridine Analysis. Dev. Biol. 2000, 226, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Ladurner, P.; Pfister, D.; Seifarth, C.; Mahlknecht, M.; Salvenmoser, W.; Gerth, R.; Marx, F.; Rieger, R. Production and characterisation of cell- and tissue-specific monoclonal antibodies for the flatworm Macrostomum sp. Histochem. Cell Biol. 2005, 123, 89–104. [Google Scholar] [CrossRef]
- Pfister, D.; De Mulder, K.; Philipp, I.; Kuales, G.; Hrouda, M.; Eichberger, P.; Borgonie, G.; Hartenstein, V.; Ladurner, P. The exceptional stem cell system of Macrostomum lignano: Screening for gene expression and studying cell proliferation by hydroxyurea treatment and irradiation. Front. Zool. 2007, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Pfister, D.; De Mulder, K.; Hartenstein, V.; Kuales, G.; Borgonie, G.; Marx, F.; Morris, J.; Ladurner, P. Flatworm stem cells and the germ line: Developmental and evolutionary implications of macvasa expression in Macrostomum lignano. Dev. Biol. 2008, 319, 146–159. [Google Scholar] [CrossRef]
- Wudarski, J.; Simanov, D.; Ustyantsev, K.; de Mulder, K.; Grelling, M.; Grudniewska, M.; Beltman, F.; Glazenburg, L.; Demircan, T.; Wunderer, J.; et al. Efficient transgenesis and annotated genome sequence of the regenerative flatworm model Macrostomum lignano. Nat. Commun. 2017, 8, 2120. [Google Scholar] [CrossRef] [Green Version]
- Zadesenets, K.S.; Schärer, L.; Rubtsov, N.B. New insights into the karyotype evolution of the free-living flatworm Macrostomum lignano (Platyhelminthes, Turbellaria). Sci. Rep. 2017, 7, 6066. [Google Scholar] [CrossRef]
- Lengerer, B.; Pjeta, R.; Wunderer, J.; Rodrigues, M.; Arbore, R.; Schärer, L.; Berezikov, E.; Hess, M.W.; Pfaller, K.; Egger, B.; et al. Biological adhesion of the flatworm Macrostomum lignano relies on a duo-gland system and is mediated by a cell type-specific intermediate filament protein. Front. Zool. 2014, 11, 12. [Google Scholar] [CrossRef] [Green Version]
- Grudniewska, M.; Mouton, S.; Simanov, D.; Beltman, F.; Grelling, M.; de Mulder, K.; Arindrarto, W.; Weissert, P.M.; van der Elst, S.; Berezikov, E. Transcriptional signatures of somatic neoblasts and germline cells in Macrostomum lignano. eLife 2016, 5, e20607. [Google Scholar] [CrossRef]
- Mouton, S.; Wudarski, J.; Grudniewska, M.; Berezikov, E. The regenerative flatworm Macrostomum lignano, a model organism with high experimental potential. Int. J. Dev. Biol. 2018, 62, 551–558. [Google Scholar] [CrossRef] [Green Version]
- De Miguel-Bonet, M.D.M.; Ahad, S.; Hartenstein, V. Role of neoblasts in the patterned postembryonic growth of the platyhelminth Macrostomum lignano. Neurogenesis 2018, 5, e1469944. [Google Scholar] [CrossRef] [Green Version]
- Grudniewska, M.; Mouton, S.; Grelling, M.; Wolters, A.H.G.; Kuipers, J.; Giepmans, B.N.G.; Berezikov, E. A novel flatworm-specific gene implicated in reproduction in Macrostomum lignano. Sci. Rep. 2018, 8, 3192. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Battistoni, G.; El Demerdash, O.; Gurtowski, J.; Wunderer, J.; Falciatori, I.; Ladurner, P.; Schatz, M.C.; Hannon, G.J.; Wasik, K.A. Dual functions of Macpiwi1 in transposon silencing and stem cell maintenance in the flatworm Macrostomum lignano. RNA 2015, 21, 1885–1897. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-O.; Kim, H.-N.; Kim, K.-H.; Baek, E.J.; Park, J.-Y.; Ha, K.; Heo, D.R.; Seo, M.-D.; Park, S.G. Development and characterization of a fully human antibody targeting SCF/c-kit signaling. Int. J. Biol. Macromol. 2020, 159, 66–78. [Google Scholar] [CrossRef]
- Deng, L.; Meng, T.; Chen, L.; Wei, W.; Wang, P. The role of ubiquitination in tumorigenesis and targeted drug discovery. Signal Transduct. Target. Ther. 2020, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Nayak, S.; Santiago, F.; Jin, H.; Lin, D.; Schedl, T.; Kipreos, E.T. The Caenorhabditis elegans Skp1-Related Gene Family: Diverse Functions in Cell Proliferation, Morphogenesis, and Meiosis. Curr. Biol. 2002, 12, 277–287. [Google Scholar] [CrossRef] [Green Version]
- Guan, Y.; Leu, N.A.; Ma, J.; Chmátal, L.; Ruthel, G.; Bloom, J.C.; Lampson, M.A.; Schimenti, J.C.; Luo, M.; Wang, P.J. SKP1 drives the prophase I to metaphase I transition during male meiosis. Sci. Adv. 2020, 6, eaaz2129. [Google Scholar] [CrossRef]
- Guan, Y.; Lin, H.; Leu, N.A.; Ruthel, G.; Fuchs, S.Y.; Busino, L.; Luo, M.; Wang, P.J. SCF ubiquitin E3 ligase regulates DNA double-strand breaks in early meiotic recombination. Nucleic Acids Res. 2022, 50, 5129–5144. [Google Scholar] [CrossRef] [PubMed]
- Strand, N.S.; Allen, J.M.; Ghulam, M.; Taylor, M.R.; Munday, R.K.; Carrillo, M.; Movsesyan, A.; Zayas, R.M. Dissecting the function of Cullin-RING ubiquitin ligase complex genes in planarian regeneration. Dev. Biol. 2018, 433, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Ladurner, P.; Scharer, L.; Salvenmoser, W.; Rieger, R.M. A new model organism among the lower Bilateria and the use of digital microscopy in taxonomy of meiobenthic Platyhelminthes: Macrostomum lignano, n. sp. (Rhabditophora, Macrostomorpha). J. Zool. Syst. Evol. Res. 2005, 43, 114–126. [Google Scholar] [CrossRef]
- Sekii, K.; Salvenmoser, W.; De Mulder, K.; Scharer, L.; Ladurner, P. Melav2, an elav-like gene, is essential for spermatid differentiation in the flatworm Macrostomum lignano. BMC Dev. Biol. 2009, 9, 62. [Google Scholar] [CrossRef] [Green Version]
- Andersen, R.A. Algal Culturing Techniques; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Orii, H.; Mochii, M.; Watanabe, K. A simple “soaking method” for RNA interference in the planarian Dugesia japonica. Dev. Genes Evol. 2003, 213, 138–141. [Google Scholar] [CrossRef]
- Wudarski, J.; Ustyantsev, K.; Glazenburg, L.; Berezikov, E. Influence of temperature on development, reproduction and regeneration in the flatworm model organism, Macrostomum lignano. Zool. Lett. 2019, 5, 7. [Google Scholar] [CrossRef] [Green Version]
- Egger, B.; Ladurner, P.; Nimeth, K.; Gschwentner, R.; Rieger, R. The regeneration capacity of the flatworm Macrostomum lignano—On repeated regeneration, rejuvenation, and the minimal size needed for regeneration. Dev. Genes Evol. 2006, 216, 565–577. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biryukov, M.; Dmitrieva, A.; Vavilova, V.; Ustyantsev, K.; Bazarova, E.; Sukhikh, I.; Berezikov, E.; Blinov, A. Mlig-SKP1 Gene Is Required for Spermatogenesis in the Flatworm Macrostomum lignano. Int. J. Mol. Sci. 2022, 23, 15110. https://doi.org/10.3390/ijms232315110
Biryukov M, Dmitrieva A, Vavilova V, Ustyantsev K, Bazarova E, Sukhikh I, Berezikov E, Blinov A. Mlig-SKP1 Gene Is Required for Spermatogenesis in the Flatworm Macrostomum lignano. International Journal of Molecular Sciences. 2022; 23(23):15110. https://doi.org/10.3390/ijms232315110
Chicago/Turabian StyleBiryukov, Mikhail, Anastasia Dmitrieva, Valeriya Vavilova, Kirill Ustyantsev, Erzhena Bazarova, Igor Sukhikh, Eugene Berezikov, and Alexandr Blinov. 2022. "Mlig-SKP1 Gene Is Required for Spermatogenesis in the Flatworm Macrostomum lignano" International Journal of Molecular Sciences 23, no. 23: 15110. https://doi.org/10.3390/ijms232315110