
Eating the Enemy: Mycoplasma Strategies to Evade Neutrophil Extracellular Traps (NETs) Promoting Bacterial Nucleotides Uptake and Inflammatory Damage


Abstract
:1. Introduction
2. Role of NETs during Bacterial Infection
3. Mycoplasmas and NETs

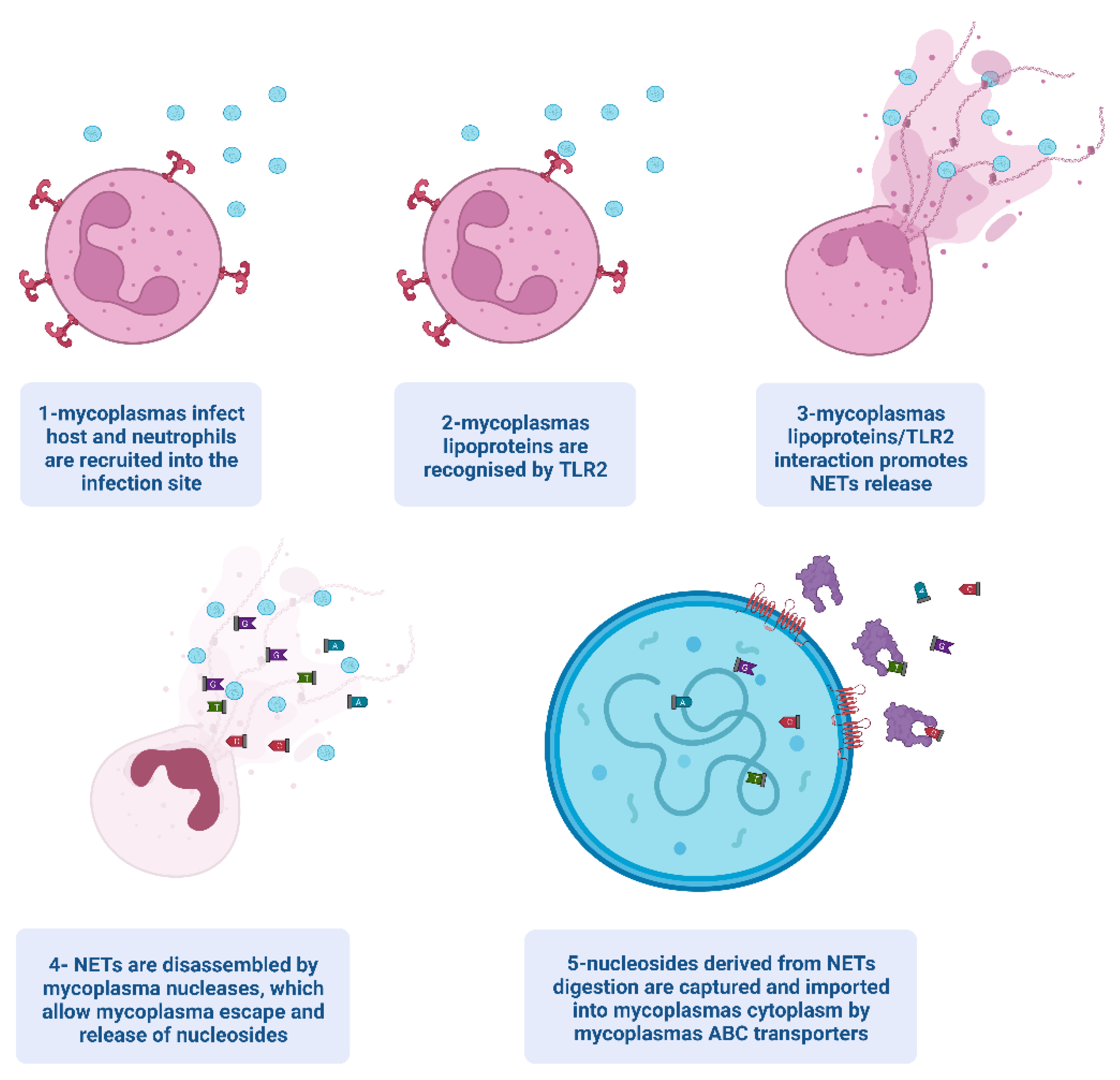{kind=link}
| Species | Nuclease Name | Subcellular Localization | Type of Nuclease | Substrate | Required Divalent Cation | References |
|---|---|---|---|---|---|---|
| M. agalactiae | MAG_5040 | Membrane-exposed | Endo/Exo | ssDNA, dsDNA, RNA, and plasmid | Mg2+ | [38] |
| M. bovis | MnuA | Membrane-exposed | Endo/Exo | dsDNA, plasmid, | Ca2+, Mg2+ | [109] |
| MbovNase | Membrane-bound and secreted | Endo | dsDNA, RNA, and plasmid | Ca2+ | [110] | |
| M. hominis | MHO_0730 | Membrane-exposed | Endo/Exo | ssDNA, dsDNA, RNA, plasmid | Ca2+ | [34] |
| M. pneumoniae | Mpn133 | Membrane-associated | Endo/Exo | ssDNA, dsDNA, RNA, and plasmid | Ca2+ | [111] |
| Mpn491 | Secreted | DNAse | DNA | Mg2 | [37] | |
| M. genitalium | MG_186 | Membrane-associated | Endo/Exo | ssDNA, dsDNA, RNA, and plasmid | Ca2+ | [112] |
| M. hyopneumoniae | Mhp379 | Membrane-exposed | Endo/Exo | ssDNA, dsDNA, RNA, plasmid | Ca2+ | [113] |
| Mhp597 | Secreted | Endo/Exo | ssDNA, dsDNA, RNA, and plasmid | Ca2+, Mg2+ | [40] | |
| M. gallisepticum | MGA_0676 | Membrane-exposed | Endo/Exo | ssDNA, dsDNA, RNA, and plasmid | Ca2+ | [114] |
| M. meleagridis | Mm19 | Membrane-exposed | Endo/Exo | ssDNA, dsDNA, RNA, and plasmid | Mg2 | [115] |
| M. pulmonis | MnuA | Membrane | Undefined | DNA | Undefined | [116] |
| M. penetrans | P40 | Membrane | Endo/Exo | ssDNA, dsDNA, RNA, and plasmid | Ca2+, Mg2+ | [117,118] |
3.1. Mycoplasma agalactiae
3.2. Mycoplasma bovis
3.3. Mycoplasma hominis
3.4. Mycoplasma pneumoniae
3.5. Mycoplasma hyopneumoniae
4. Relevance of NETs to Immunity and Mycoplasma Acute and Chronic Infections
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rosales, C.; Lowell, C.A.; Schnoor, M.; Uribe-Querol, E. Editorial Neutrophils: Their Role in Innate and Adaptive Immunity 2017. J. Immunol. Res. 2017, 2017, 9748345. [Google Scholar] [CrossRef]
- Gierlikowska, B.; Stachura, A.; Gierlikowski, W.; Demkow, U. Phagocytosis, Degranulation and Extracellular Traps Release by Neutrophils—The Current Knowledge, Pharmacological Modulation and Future Prospects. Front. Pharmacol. 2021, 12, 666732. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil Extracellular Traps Kill Bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2017, 18, 134–147. [Google Scholar] [CrossRef]
- Kaplan, M.J.; Radic, M. Neutrophil extracellular traps (NETs): Double-edged swords of innate immunity. J. Immunol. 2012, 189, 2689. [Google Scholar] [CrossRef] [Green Version]
- Monteith, A.J.; Miller, J.M.; Maxwell, C.N.; Chazin, W.J.; Skaar, E.P. Neutrophil extracellular traps enhance macrophage killing of bacterial pathogens. Sci. Adv. 2021, 7, eabj2101. [Google Scholar] [CrossRef] [PubMed]
- Urban, C.F.; Ermert, D.; Schmid, M.; Abu-Abed, U.; Goosmann, C.; Nacken, W.; Brinkmann, V.; Jungblut, P.R.; Zychlinsky, A. Neutrophil Extracellular Traps Contain Calprotectin, a Cytosolic Protein Complex Involved in Host Defense against Candida albicans. PLoS Pathog. 2009, 5, e1000639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juneau, R.A.; Pang, B.; Weimer, K.W.D.; Armbruster, C.E.; Swords, W.E. Nontypeable haemophilus influenzae initiates formation of neutrophil extracellular traps. Infect. Immun. 2011, 79, 431–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schönrich, G.; Raftery, M.J. Neutrophil extracellular traps go viral. Front. Immunol. 2016, 7, 366. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Liu, J.; Chen, Y.; Yan, L.; Wu, J. Neutrophil Extracellular Traps in Candida albicans Infection. Front. Immunol. 2022, 13, 913028. [Google Scholar] [CrossRef]
- Urban, C.F.; Nett, J.E. Neutrophil extracellular traps in fungal infection. Semin. Cell Dev. Biol. 2019, 89, 47–57. [Google Scholar] [CrossRef]
- Abi Abdallah, D.S.; Denkers, E.Y. Neutrophils cast extracellular traps in response to protozoan parasites. Front. Immunol. 2012, 3, 382. [Google Scholar] [CrossRef] [Green Version]
- Guimarães-Costa, A.B.; Nascimento, M.T.C.; Froment, G.S.; Soares, R.P.P.; Morgado, F.N.; Conceição-Silva, F.; Saraiva, E.M. Leishmania amazonensis promastigotes induce and are killed by neutrophil extracellular traps. Proc. Natl. Acad. Sci. USA 2009, 106, 6748–6753. [Google Scholar] [CrossRef] [Green Version]
- Jenne, C.N.; Wong, C.H.Y.; Zemp, F.J.; McDonald, B.; Rahman, M.M.; Forsyth, P.A.; McFadden, G.; Kubes, P. Neutrophils Recruited to Sites of Infection Protect from Virus Challenge by Releasing Neutrophil Extracellular Traps. Cell Host Microbe 2013, 13, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Saitoh, T.; Komano, J.; Saitoh, Y.; Misawa, T.; Takahama, M.; Kozaki, T.; Uehata, T.; Iwasaki, H.; Omori, H.; Yamaoka, S.; et al. Neutrophil Extracellular Traps Mediate a Host Defense Response to Human Immunodeficiency Virus-1. Cell Host Microbe 2012, 12, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Cacciotto, C.; Cubeddu, T.; Addis, M.F.M.F.M.F.; Anfossi, A.G.A.G.; Tedde, V.; Tore, G.; Carta, T.; Rocca, S.; Chessa, B.; Pittau, M.; et al. Mycoplasma lipoproteins are major determinants of neutrophil extracellular trap formation. Cell. Microbiol. 2016, 18, 1751–1762. [Google Scholar] [CrossRef]
- Nakamura, K.; Nakayama, H.; Sasaki, S.; Takahashi, K.; Iwabuchi, K. Mycobacterium avium-intracellulare complex promote release of pro-inflammatory enzymes matrix metalloproteinases by inducing neutrophil extracellular trap formation. Sci. Rep. 2022, 12, 5181. [Google Scholar] [CrossRef] [PubMed]
- Janssen, L.; Muller, H.S.; Martins, V.d.P. Unweaving the NET: Microbial strategies for neutrophil extracellular trap evasion. Microb. Pathog. 2022, 171, 105728. [Google Scholar] [CrossRef] [PubMed]
- Sollberger, G.; Tilley, D.O.; Zychlinsky, A. Neutrophil Extracellular Traps: The Biology of Chromatin Externalization. Dev. Cell 2018, 44, 542–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paessler, M.; Levinson, A.; Patel, J.B.; Schuster, M.; Minda, M.; Nachamkin, I. Disseminated Mycoplasma orale infection in a patient with common variable immunodeficiency syndrome. Diagn. Microbiol. Infect. Dis. 2002, 44, 201–204. [Google Scholar] [CrossRef]
- Razin, S.; Yogev, D.; Naot, Y. Molecular biology and pathogenicity of mycoplasmas. Microbiol. Mol. Biol. Rev. 1998, 62, 1094–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yavlovich, A.; Tarshis, M.; Rottem, S. Internalization and intracellular survival of Mycoplasma pneumoniae by non-phagocytic cells. FEMS Microbiol. Lett. 2004, 233, 241–246. [Google Scholar] [CrossRef]
- Yavlovich, A.; Katzenell, A.; Tarshis, M.; Higazi, A.A.R.; Rottem, S. Mycoplasma fermentans binds to and invades HeLa cells: Involvement of plasminogen and urokinase. Infect. Immun. 2004, 72, 5004–5011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Merwe, J.; Prysliak, T.; Perez-Casal, J. Invasion of bovine peripheral blood mononuclear cells and erythrocytes by Mycoplasma bovis. Infect. Immun. 2010, 78, 4570–4578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raj, J.; Rawre, J.; Dhawan, N.; Khanna, N.; Dhawan, B. Mycoplasma genitalium: A new superbug. Indian J. Sex. Transm. Dis. AIDS 2022, 43, 1. [Google Scholar] [CrossRef]
- Winner, F.; Rosengarten, R.; Citti, C. In vitro cell invasion of Mycoplasma gallisepticum. Infect. Immun. 2000, 68, 4238–4244. [Google Scholar] [CrossRef] [Green Version]
- Rappelli, P.; Addis, M.F.; Carta, F.; Fiori, P.L. Mycoplasma hominis parasitism of Trichomonas vaginalis. Lancet 1998, 352, 1286. [Google Scholar] [CrossRef]
- Chambaud, I.; Heilig, R.; Ferris, S.; Barbe, V.; Samson, D.; Galisson, F.; Moszer, I.; Dybvig, K.; Wróblewski, H.; Viari, A.; et al. The complete genome sequence of the murine respiratory pathogen Mycoplasma pulmonis. Nucleic Acids Res. 2001, 29, 2145–2153. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Westberg, J.; Bölske, G.; Eriksson, S. Novel deoxynucleoside-phosphorylating enzymes in mycoplasmas: Evidence for efficient utilization of deoxynucleosides. Mol. Microbiol. 2001, 42, 1065–1073. [Google Scholar] [CrossRef]
- Benedetti, F.; Curreli, S.; Zella, D. Mycoplasmas–Host Interaction: Mechanisms of Inflammation and Association with Cellular Transformation. Microorganisms 2020, 8, 1351. [Google Scholar] [CrossRef]
- Baseman, J.B.; Tully, J.G. Mycoplasmas: Sophisticated, reemerging, and burdened by their notoriety. Emerg. Infect. Dis. 1997, 3, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Howard, C.J.; Taylor, G. Interaction of mycoplasmas and phagocytes. Yale J. Biol. Med. 1983, 56, 643–648. [Google Scholar]
- Pisanu, S.; Cubeddu, T.; Pagnozzi, D.; Rocca, S.; Cacciotto, C.; Alberti, A.; Marogna, G.; Uzzau, S.; Addis, M.F. Neutrophil extracellular traps in sheep mastitis. Vet. Res. 2015, 46, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cacciotto, C.; Dessì, D.; Cubeddu, T.; Cocco, A.R.; Pisano, A.; Tore, G.; Fiori, P.L.; Rappelli, P.; Pittau, M.; Alberti, A. MHO-0730 as a Surface-Exposed Calcium-Dependent Nuclease of Mycoplasma hominis Promoting Neutrophil Extracellular Trap Formation and Escape. J. Infect. Dis. 2019, 220, 1999–2008. [Google Scholar] [CrossRef] [PubMed]
- Henthorn, C.R.; Minion, F.C.; Sahin, O. Utilization of macrophage extracellular trap nucleotides by Mycoplasma hyopneumoniae. Microbiology 2018, 164, 1394–1404. [Google Scholar] [CrossRef]
- Gondaira, S.; Higuchi, H.; Nishi, K.; Iwano, H.; Nagahata, H. Mycoplasma bovis escapes bovine neutrophil extracellular traps. Vet. Microbiol. 2017, 199, 68–73. [Google Scholar] [CrossRef]
- Yamamoto, T.; Kida, Y.; Sakamoto, Y.; Kuwano, K. Mpn491, a secreted nuclease of Mycoplasma pneumoniae, plays a critical role in evading killing by neutrophil extracellular traps. Cell. Microbiol. 2016, 19, e12666. [Google Scholar] [CrossRef] [Green Version]
- Cacciotto, C.; Addis, M.F.; Coradduzza, E.; Carcangiu, L.; Nuvoli, A.M.; Tore, G.; Dore, G.M.; Pagnozzi, D.; Uzzau, S.; Chessa, B.; et al. Mycoplasma agalactiae MAG_5040 is a Mg2+-Dependent, Sugar-Nonspecific SNase Recognised by the Host Humoral Response during Natural Infection. PLoS ONE 2013, 8, e57775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitiku, F.; Hartley, C.A.; Sansom, F.M.; Coombe, J.E.; Mansell, P.D.; Beggs, D.S.; Browning, G.F. The major membrane nuclease MnuA degrades neutrophil extracellular traps induced by Mycoplasma bovis. Vet. Microbiol. 2018, 218, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhang, Y.; Li, X.; Zhou, W.; Li, X.; Jiang, F.; Wu, W. Mycoplasma hyopneumoniae Mhp597 is a cytotoxicity, inflammation and immunosuppression associated nuclease. Vet. Microbiol. 2019, 235, 53–62. [Google Scholar] [CrossRef]
- Wardini, A.B.; Guimarães-Costa, A.B.; Nascimento, M.T.C.; Nadaes, N.R.; Danelli, M.G.M.; Mazur, C.; Benjamin, C.F.; Saraiva, E.M.; Pinto-da-Silva, L.H. Characterization of neutrophil extracellular traps in cats naturally infected with feline leukemia virus. J. Gen. Virol. 2010, 91, 259–264. [Google Scholar] [CrossRef]
- Palić, D.; Ostojić, J.; Andreasen, C.B.; Roth, J.A. Fish cast NETs: Neutrophil extracellular traps are released from fish neutrophils. Dev. Comp. Immunol. 2007, 31, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Worku, M.; Rehrah, D.; Ismail, H.D.; Asiamah, E.; Adjei-Fremah, S. A Review of the Neutrophil Extracellular Traps (NETs) from Cow, Sheep and Goat Models. Int. J. Mol. Sci 2021, 22, 8046. [Google Scholar] [CrossRef] [PubMed]
- Pilsczek, F.H.; Salina, D.; Poon, K.K.H.; Fahey, C.; Yipp, B.G.; Sibley, C.D.; Robbins, S.M.; Green, F.H.Y.; Surette, M.G.; Sugai, M.; et al. A novel mechanism of rapid nuclear neutrophil extracellular trap formation in response to Staphylococcus aureus. J. Immunol. 2010, 185, 7413–7425. [Google Scholar] [CrossRef] [Green Version]
- Seper, A.; Hosseinzadeh, A.; Gorkiewicz, G.; Lichtenegger, S.; Roier, S.; Leitner, D.R.; Röhm, M.; Grutsch, A.; Reidl, J.; Urban, C.F.; et al. Vibrio cholerae Evades Neutrophil Extracellular Traps by the Activity of Two Extracellular Nucleases. PLOS Pathog. 2013, 9, e1003614. [Google Scholar] [CrossRef] [Green Version]
- Beiter, K.; Wartha, F.; Albiger, B.; Normark, S.; Zychlinsky, A.; Henriques-Normark, B. An Endonuclease Allows Streptococcus pneumoniae to Escape from Neutrophil Extracellular Traps. Curr. Biol. 2006, 16, 401–407. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Kichik, V.; Mondragón-Flores, R.; Mondragón-Castelán, M.; Gonzalez-Pozos, S.; Muñiz-Hernandez, S.; Rojas-Espinosa, O.; Chacón-Salinas, R.; Estrada-Parra, S.; Estrada-García, I. Neutrophil extracellular traps are induced by Mycobacterium tuberculosis. Tuberculosis 2009, 89, 29–37. [Google Scholar] [CrossRef]
- Fuchs, T.A.; Abed, U.; Goosmann, C.; Hurwitz, R.; Schulze, I.; Wahn, V.; Weinrauch, Y.; Brinkmann, V.; Zychlinsky, A. Novel cell death program leads to neutrophil extracellular traps. J. Cell Biol. 2007, 176, 231–241. [Google Scholar] [CrossRef]
- Steinberg, B.E.; Grinstein, S. Unconventional roles of the NADPH oxidase: Signaling, ion homeostasis, and cell death. Sci. STKE 2007, 2007, pe11. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Zychlinsky, A. Beneficial suicide: Why neutrophils die to make NETs. Nat. Rev. Microbiol. 2007, 5, 577–582. [Google Scholar] [CrossRef]
- Yang, H.; Biermann, M.H.; Brauner, J.M.; Liu, Y.; Zhao, Y.; Herrmann, M. New insights into neutrophil extracellular traps: Mechanisms of formation and role in inflammation. Front. Immunol. 2016, 7, 302. [Google Scholar] [CrossRef]
- Halverson, T.W.R.; Wilton, M.; Poon, K.K.H.; Petri, B.; Lewenza, S. DNA is an antimicrobial component of neutrophil extracellular traps. PLoS Pathog. 2015, 11, e1004593. [Google Scholar] [CrossRef] [Green Version]
- Brinkmann, V.; Zychlinsky, A. Neutrophil extracellular traps: Is immunity the second function of chromatin? J. Cell Biol. 2012, 198, 773–783. [Google Scholar] [CrossRef] [Green Version]
- Riyapa, D.; Buddhisa, S.; Korbsrisate, S.; Cuccui, J.; Wren, B.W.; Stevens, M.P.; Ato, M.; Lertmemongkolchai, G. Neutrophil extracellular traps exhibit antibacterial activity against Burkholderia pseudomallei and are influenced by bacterial and host factors. Infect. Immun. 2012, 80, 3921–3929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menegazzi, R.; Decleva, E.; Dri, P. Killing by neutrophil extracellular traps: Fact or folklore? Blood 2012, 119, 1214–1216. [Google Scholar] [CrossRef] [Green Version]
- Rice, W.G.; Weiss, S.J. Regulation of proteolysis at the neutrophil-substrate interface by secretory leukoprotease inhibitor. Science 1990, 249, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Weiss, S.J.; Regiani, S. Neutrophils degrade subendothelial matrices in the presence of alpha-1-proteinase inhibitor. Cooperative use of lysosomal proteinases and oxygen metabolites. J. Clin. Investig. 1984, 73, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Azzouz, L.; Cherry, A.; Riedl, M.; Khan, M.; Pluthero, F.G.; Kahr, W.H.A.; Palaniyar, N.; Licht, C. Relative antibacterial functions of complement and NETs: NETs trap and complement effectively kills bacteria. Mol. Immunol. 2018, 97, 71–81. [Google Scholar] [CrossRef]
- Fadok, V.A.; Bratton, D.L.; Guthrie, L.; Henson, P.M. Differential effects of apoptotic versus lysed cells on macrophage production of cytokines: Role of proteases. J. Immunol. 2001, 166, 6847–6854. [Google Scholar] [CrossRef] [Green Version]
- Nakazawa, D.; Shida, H.; Kusunoki, Y.; Miyoshi, A.; Nishio, S.; Tomaru, U.; Atsumi, T.; Ishizu, A. The responses of macrophages in interaction with neutrophils that undergo NETosis. J. Autoimmun. 2016, 67, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Apel, F.; Andreeva, L.; Knackstedt, L.S.; Streeck, R.; Frese, C.K.; Goosmann, C.; Hopfner, K.P.; Zychlinsky, A. The cytosolic DNA sensor cGAS recognizes neutrophil extracellular traps. Sci. Signal. 2021, 14, eaax7942. [Google Scholar] [CrossRef]
- Warnatsch, A.; Ioannou, M.; Wang, Q.; Papayannopoulos, V. Neutrophil extracellular traps license macrophages for cytokine production in atherosclerosis. Science 2015, 349, 316–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahlenberg, J.M.; Carmona-Rivera, C.; Smith, C.K.; Kaplan, M.J. Neutrophil extracellular trap-associated protein activation of the NLRP3 inflammasome is enhanced in lupus macrophages. J. Immunol. 2013, 190, 1217–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribon, M.; Seninet, S.; Mussard, J.; Sebbag, M.; Clavel, C.; Serre, G.; Boissier, M.C.; Semerano, L.; Decker, P. Neutrophil extracellular traps exert both pro- and anti-inflammatory actions in rheumatoid arthritis that are modulated by C1q and LL-37. J. Autoimmun. 2019, 98, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Linhares-Lacerda, L.; Temerozo, J.R.; Ribeiro-Alves, M.; Azevedo, E.P.; Mojoli, A.; Nascimento, M.T.C.; Silva-Oliveira, G.; Savino, W.; Foguel, D.; Bou-Habib, D.C.; et al. Neutrophil extracellular trap-enriched supernatants carry microRNAs able to modulate TNF-α production by macrophages. Sci. Rep. 2020, 10, 2715. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Romo, G.S.; Caielli, S.; Vega, B.; Connolly, J.; Allantaz, F.; Xu, Z.; Punaro, M.; Baisch, J.; Guiducci, C.; Coffman, R.L.; et al. Netting neutrophils are major inducers of type I IFN production in pediatric systemic lupus erythematosus. Sci. Transl. Med. 2011, 3, 73ra20. [Google Scholar] [CrossRef] [Green Version]
- Barrientos, L.; Bignon, A.; Gueguen, C.; de Chaisemartin, L.; Gorges, R.; Sandré, C.; Mascarell, L.; Balabanian, K.; Kerdine-Römer, S.; Pallardy, M.; et al. Neutrophil extracellular traps downregulate lipopolysaccharide-induced activation of monocyte-derived dendritic cells. J. Immunol. 2014, 193, 5689–5698. [Google Scholar] [CrossRef] [Green Version]
- Dömer, D.; Walther, T.; Möller, S.; Behnen, M.; Laskay, T. Neutrophil Extracellular Traps Activate Proinflammatory Functions of Human Neutrophils. Front. Immunol. 2021, 12, 1190. [Google Scholar] [CrossRef]
- Berends, E.T.M.; Horswill, A.R.; Haste, N.M.; Monestier, M.; Nizet, V.; Von Köckritz-Blickwede, M. Nuclease expression by Staphylococcus aureus facilitates escape from neutrophil extracellular traps. J. Innate Immun. 2010, 2, 576–586. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Sun, L.; Liu, W.; Guo, L.; Liu, Z.; Wei, X.; Ling, J. A nuclease from Streptococcus mutans facilitates biofilm dispersal and escape from killing by neutrophil extracellular traps. Front. Cell. Infect. Microbiol. 2017, 7, 97. [Google Scholar] [CrossRef] [Green Version]
- Juneau, R.A.; Stevens, J.S.; Apicella, M.A.; Criss, A.K. A Thermonuclease of Neisseria gonorrhoeae Enhances Bacterial Escape From Killing by Neutrophil Extracellular Traps. J. Infect. Dis. 2015, 212, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Khandpur, R.; Carmona-Rivera, C.; Vivekanandan-Giri, A.; Gizinski, A.; Yalavarthi, S.; Knight, J.S.; Friday, S.; Li, S.; Patel, R.M.; Subramanian, V.; et al. NETs are a source of citrullinated autoantigens and stimulate inflammatory responses in rheumatoid arthritis. Sci. Transl. Med. 2013, 5, 178ra40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, S.C.S.; Yu, H.S.; Yen, F.L.; Lin, C.L.; Chen, G.S.; Lan, C.C.E. Neutrophil extracellular trap formation is increased in psoriasis and induces human β-defensin-2 production in epidermal keratinocytes. Sci. Rep. 2016, 6, 31119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Zhang, H.; Wang, Y.; Brown, Z.J.; Xia, Y.; Huang, Z.; Shen, C.; Hu, Z.; Beane, J.; Ansa-Addo, E.A.; et al. Regulatory T-cell and neutrophil extracellular trap interaction contributes to carcinogenesis in non-alcoholic steatohepatitis. J. Hepatol. 2021, 75, 1271–1283. [Google Scholar] [CrossRef] [PubMed]
- Arazna, M.; Pruchniak, M.P.; Zycinska, K.; Demkow, U. Neutrophil extracellular trap in human diseases. Adv. Exp. Med. Biol. 2013, 756, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Leffler, J.; Martin, M.; Gullstrand, B.; Tydén, H.; Lood, C.; Truedsson, L.; Bengtsson, A.A.; Blom, A.M. Neutrophil Extracellular Traps That Are Not Degraded in Systemic Lupus Erythematosus Activate Complement Exacerbating the Disease. J. Immunol. 2012, 188, 3522–3531. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, T.A.; Brill, A.; Duerschmied, D.; Schatzberg, D.; Monestier, M.; Myers, D.D.; Wrobleski, S.K.; Wakefield, T.W.; Hartwig, J.H.; Wagner, D.D. Extracellular DNA traps promote thrombosis. Proc. Natl. Acad. Sci. USA 2010, 107, 15880–15885. [Google Scholar] [CrossRef] [Green Version]
- Massberg, S.; Grahl, L.; Von Bruehl, M.L.; Manukyan, D.; Pfeiler, S.; Goosmann, C.; Brinkmann, V.; Lorenz, M.; Bidzhekov, K.; Khandagale, A.B.; et al. Reciprocal coupling of coagulation and innate immunity via neutrophil serine proteases. Nat. Med. 2010, 16, 887–896. [Google Scholar] [CrossRef]
- Swystun, L.L.; Mukherjee, S.; Liaw, P.C. Breast cancer chemotherapy induces the release of cell-free DNA, a novel procoagulant stimulus. J. Thromb. Haemost. 2011, 9, 2313–2321. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, X.; Pelayo, R.; Monestier, M.; Ammollo, C.T.; Semeraro, F.; Taylor, F.B.; Esmon, N.L.; Lupu, F.; Esmon, C.T. Extracellular histones are major mediators of death in sepsis. Nat. Med. 2009, 15, 1318–1321. [Google Scholar] [CrossRef] [Green Version]
- Clancy, D.M.; Sullivan, G.P.; Moran, H.B.T.; Henry, C.M.; Reeves, E.P.; McElvaney, N.G.; Lavelle, E.C.; Martin, S.J. Extracellular Neutrophil Proteases Are Efficient Regulators of IL-1, IL-33, and IL-36 Cytokine Activity but Poor Effectors of Microbial Killing. Cell Rep. 2018, 22, 2937–2950. [Google Scholar] [CrossRef]
- Song, W.; Ye, J.; Pan, N.; Tan, C.; Herrmann, M. Neutrophil Extracellular Traps Tied to Rheumatoid Arthritis: Points to Ponder. Front. Immunol. 2020, 11, 578129. [Google Scholar] [CrossRef]
- Dannenburg, A. Infections as a Cause of Cancer: A History and Outlook. In Proceedings of the AACR Sixth Annual International Conference Frontiers in Cancer Prevention Research, Philadelphia, PA, USA, 5 December 2007. [Google Scholar]
- Masucci, M.T.; Minopoli, M.; Del Vecchio, S.; Carriero, M.V. The Emerging Role of Neutrophil Extracellular Traps (NETs) in Tumor Progression and Metastasis. Front. Immunol. 2020, 11, 1749. [Google Scholar] [CrossRef] [PubMed]
- Monti, M.; De Rosa, V.; Iommelli, F.; Carriero, M.V.; Terlizzi, C.; Camerlingo, R.; Belli, S.; Fonti, R.; Di Minno, G.; Del Vecchio, S. Neutrophil Extracellular Traps as an Adhesion Substrate for Different Tumor Cells Expressing RGD-Binding Integrins. Int. J. Mol. Sci. 2018, 19, 2350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanamaru, R.; Ohzawa, H.; Miyato, H.; Yamaguchi, H.; Hosoya, Y.; Lefor, A.K.; Sata, N.; Kitayama, J. Neutrophil Extracellular Traps Generated by Low Density Neutrophils Obtained from Peritoneal Lavage Fluid Mediate Tumor Cell Growth and Attachment. J. Vis. Exp. 2018, 2018, e58201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cedervall, J.; Zhang, Y.; Huang, H.; Zhang, L.; Femel, J.; Dimberg, A.; Olsson, A.K. Neutrophil Extracellular Traps Accumulate in Peripheral Blood Vessels and Compromise Organ Function in Tumor-Bearing Animals. Cancer Res. 2015, 75, 2653–2662. [Google Scholar] [CrossRef] [Green Version]
- Addis, M.F.; Pisanu, S.; Ghisaura, S.; Pagnozzi, D.; Marogna, G.; Tanca, A.; Biosa, G.; Cacciotto, C.; Alberti, A.; Pittau, M.; et al. Proteomics and pathway analyses of the milk fat globule in sheep naturally infected by Mycoplasma agalactiae provide indications of the in vivo response of the mammary epithelium to bacterial infection. Infect. Immun. 2011, 79, 3833–3845. [Google Scholar] [CrossRef] [Green Version]
- Cubeddu, T.; Cacciotto, C.; Pisanu, S.; Tedde, V.; Alberti, A.; Pittau, M.; Dore, S.; Cannas, A.; Uzzau, S.; Rocca, S.; et al. Cathelicidin production and release by mammary epithelial cells during infectious mastitis. Vet. Immunol. Immunopathol. 2017, 189, 66–70. [Google Scholar] [CrossRef]
- Hoppenbrouwers, T.; Autar, A.S.A.; Sultan, A.R.; Abraham, T.E.; Van Cappellen, W.A.; Houtsmuller, A.B.; Van Wamel, W.J.B.; Van Beusekom, H.M.M.; Van Neck, J.W.; De Maat, M.P.M. In vitro induction of NETosis: Comprehensive live imaging comparison and systematic review. PLoS ONE 2017, 12, e0176472. [Google Scholar] [CrossRef] [Green Version]
- Kenny, E.F.; Herzig, A.; Krüger, R.; Muth, A.; Mondal, S.; Thompson, P.R.; Brinkmann, V.; von Bernuth, H.; Zychlinsky, A. Diverse stimuli engage different neutrophil extracellular trap pathways. Elife 2017, 6, e24437. [Google Scholar] [CrossRef]
- Petretto, A.; Bruschi, M.; Pratesi, F.; Croia, C.; Candiano, G.; Ghiggeri, G.; Migliorini, P. Neutrophil extracellular traps (NET) induced by different stimuli: A comparative proteomic analysis. PLoS ONE 2019, 14, e0218946. [Google Scholar] [CrossRef] [PubMed]
- Rosengarten, R.; Citti, C.; Glew, M.; Lischewski, A.; Droeße, M.; Much, P.; Winner, F.; Brank, M.; Spergser, J. Host-pathogen interactions in mycoplasma pathogenesis: Virulence and survival strategies of minimalist prokaryotes. Int. J. Med. Microbiol. 2000, 290, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Cacciotto, C.; Addis, M.F.; Pagnozzi, D.; Chessa, B.; Coradduzza, E.; Carcangiu, L.; Uzzau, S.; Alberti, A.; Pittau, M. The liposoluble proteome of Mycoplasma agalactiae: An insight into the minimal protein complement of a bacterial membrane. BMC Microbiol. 2010, 10, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browning, G.F.; Marenda, M.S.; Noormohammadi, A.H.; Markham, P.F. The central role of lipoproteins in the pathogenesis of mycoplasmoses. Vet. Microbiol. 2011, 153, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Hegde, S.; Zimmermann, M.; Rosengarten, R.; Chopra-Dewasthaly, R. Novel role of Vpmas as major adhesins of Mycoplasma agalactiae mediating differential cell adhesion and invasion of Vpma expression variants. Int. J. Med. Microbiol. 2018, 308, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Christodoulides, A.; Gupta, N.; Yacoubian, V.; Maithel, N.; Parker, J.; Kelesidis, T. The Role of Lipoproteins in Mycoplasma-Mediated Immunomodulation. Front. Microbiol. 2018, 9, 1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bird, D.W.; Graber, K.; Knutson, A.; Washburn, L.R. Mutation of two Mycoplasma arthritidis surface lipoproteins with divergent functions in cytadherence. Infect. Immun. 2008, 76, 5768–5776. [Google Scholar] [CrossRef] [Green Version]
- May, M.; Dunne, D.W.; Brown, D.R. A Sialoreceptor Binding Motif in the Mycoplasma synoviae Adhesin VlhA. PLoS ONE 2014, 9, e110360. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Q.; Wang, J.; Ji, Y.; Ni, B.; Zhang, B.; Ma, Q.; Wei, Y.; Xiao, S.; Feng, Z.; Liu, M.; et al. The functions of the variable lipoprotein family of Mycoplasma hyorhinis in adherence to host cells. Vet. Microbiol. 2016, 186, 82–89. [Google Scholar] [CrossRef]
- Schenk, M.; Belisle, J.T.; Modlin, R.L. TLR2 Looks at Lipoproteins. Immunity 2009, 31, 847–849. [Google Scholar] [CrossRef] [Green Version]
- Kurokawa, K.; Ryu, K.H.; Ichikawa, R.; Masuda, A.; Kim, M.S.; Lee, H.; Chae, J.H.; Shimizu, T.; Saitoh, T.; Kuwano, K.; et al. Novel Bacterial Lipoprotein Structures Conserved in Low-GC Content Gram-positive Bacteria Are Recognized by Toll-like Receptor 2. J. Biol. Chem. 2012, 287, 13170–13181. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Kida, Y.; Kuwano, K. A triacylated lipoprotein from Mycoplasma genitalium activates NF-kappaB through Toll-like receptor 1 (TLR1) and TLR2. Infect. Immun. 2008, 76, 3672–3678. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, T. Inflammation-inducing Factors of Mycoplasma pneumoniae. Front. Microbiol. 2016, 7, 414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gondaira, S.; Nishi, K.; Fujiki, J.; Iwano, H.; Watanabe, R.; Eguchi, A.; Hirano, Y.; Higuchi, H.; Nagahata, H. Innate immune response in bovine neutrophils stimulated with Mycoplasma bovis. Vet. Res. 2021, 52, 58. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Lopez, N.; Romero, R.; Xu, Y.; Miller, D.; Unkel, R.; Shaman, M.; Jacques, S.M.; Panaitescu, B.; Garcia-Flores, V.; Hassan, S.S. Neutrophil Extracellular Traps in the Amniotic Cavity of Women with Intra-Amniotic Infection: A New Mechanism of Host Defense. Reprod. Sci. 2017, 24, 1139–1153. [Google Scholar] [CrossRef]
- Minion, F.C.; Jarvill-Taylor, K.J.; Billings, D.E.; Tigges, E. Membrane-associated nuclease activities in mycoplasmas. J. Bacteriol. 1993, 175, 7842–7847. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, A.; Finch, L.R. Pathways of nucleotide biosynthesis in Mycoplasma mycoides subsp. mycoides. J. Bacteriol. 1977, 130, 1047–1054. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Tivendale, K.A.; Markham, P.F.; Browning, G.F. Disruption of the membrane nuclease gene (MBOVPG45_0215) of Mycoplasma bovis greatly reduces cellular nuclease activity. J. Bacteriol. 2015, 197, 1549–1558. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhao, G.; Guo, Y.; Menghwar, H.; Chen, Y.; Chen, H.; Guo, A. Mycoplasma bovis MBOV_RS02825 Encodes a Secretory Nuclease Associated with Cytotoxicity. Int. J. Mol. Sci. 2016, 17, 628. [Google Scholar] [CrossRef] [Green Version]
- Somarajan, S.R.; Kannan, T.R.; Baseman, J.B. Mycoplasma pneumoniae Mpn133 is a cytotoxic nuclease with a glutamic acid-, lysine- and serine-rich region essential for binding and internalization but not enzymatic activity. Cell. Microbiol. 2010, 12, 1821–1831. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Krishnan, M.; Baseman, J.B.; Kannan, T.R. Molecular cloning, expression, and characterization of a Ca2+-dependent, membrane-associated nuclease of Mycoplasma genitalium. J. Bacteriol. 2010, 192, 4876–4884. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.A.; Browning, G.F.; Markham, P.F. Mycoplasma hyopneumoniae mhp379 is a Ca2+-dependent, sugar-nonspecific exonuclease exposed on the cell surface. J. Bacteriol. 2007, 189, 3414–3424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Teng, D.; Jiang, F.; Zhang, Y.; El-Ashram, S.A.; Wang, H.; Sun, Z.; He, J.; Shen, J.; Wu, W.; et al. Mycoplasma gallisepticum MGA_0676 is a membrane-associated cytotoxic nuclease with a staphylococcal nuclease region essential for nuclear translocation and apoptosis induction in chicken cells. Appl. Microbiol. Biotechnol. 2014, 99, 1859–1871. [Google Scholar] [CrossRef] [PubMed]
- Yacoub, E.; Mardassi, B.B.A. Mm19, a Mycoplasma meleagridis Major Surface Nuclease that Is Related to the RE_AlwI Superfamily of Endonucleases. PLoS ONE 2016, 11, e0152171. [Google Scholar] [CrossRef]
- Jarvill-Taylor, K.J.; Vandyk, C.; Minion, F.C. Cloning of mnuA, a membrane nuclease gene of Mycoplasma pulmonis, and analysis of its expression in Escherichia coli. J. Bacteriol. 1999, 181, 1853–1860. [Google Scholar] [CrossRef] [Green Version]
- Bendjennat, M.; Blanchard, A.; Loutfi, M.; Montagnier, L.; Bahraoui, E. Purification and characterization of Mycoplasma penetrans Ca2+/Mg2+- dependent endonuclease. J. Bacteriol. 1997, 179, 2210–2220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bendjennat, M.; Blanchard, A.; Loutfi, M.; Montagnier, L.; Bahraoui, E. Role of Mycoplasma penetrans endonuclease P40 as a potential pathogenic determinant. Infect. Immun. 1999, 67, 4456–4462. [Google Scholar] [CrossRef] [Green Version]
- Klose, S.M.; Wawegama, N.; Sansom, F.M.; Marenda, M.S.; Browning, G.F. Efficient disruption of the function of the mnuA nuclease gene using the endogenous CRISPR/Cas system in Mycoplasma gallisepticum. Vet. Microbiol. 2022, 269, 109436. [Google Scholar] [CrossRef]
- Migliore, S.; Puleio, R.; Nicholas, R.A.J.; Loria, G.R. Mycoplasma agalactiae: The Sole Cause of Classical Contagious Agalactia? Animals 2021, 11, 1782. [Google Scholar] [CrossRef]
- DaMassa, A.J.; Brooks, D.L. The external ear canal of goats and other animals as a mycoplasma habitat. Small Rumin. Res. 1991, 4, 85–93. [Google Scholar] [CrossRef]
- Kumar, A.; Rahal, A.; Chakraborty, S.; Verma, A.K.; Dhama, K. Mycoplasma agalactiae, an Etiological Agent of Contagious Agalactia in Small Ruminants: A Review. Vet. Med. Int. 2014, 2014, 286752. [Google Scholar] [CrossRef]
- Sirand-Pugnet, P.; Lartigue, C.; Marenda, M.; Jacob, D.; Barré, A.; Barbe, V.; Schenowitz, C.; Mangenot, S.; Couloux, A.; Segurens, B.; et al. Being pathogenic, plastic, and sexual while living with a nearly minimal bacterial genome. PLoS Genet. 2007, 3, e75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nouvel, L.X.; Sirand-Pugnet, P.; Marenda, M.S.; Sagné, E.; Barbe, V.; Mangenot, S.; Schenowitz, C.; Jacob, D.; Barré, A.; Claverol, S.; et al. Comparative genomic and proteomic analyses of two Mycoplasma agalactiae strains: Clues to the macro- and micro-events that are shaping mycoplasma diversity. BMC Genom. 2010, 11, 86. [Google Scholar] [CrossRef]
- Fleury, B.; Bergonier, D.; Berthelot, X.; Peterhans, E.; Frey, J.; Vilei, E.M. Characterization of P40, a cytadhesin of Mycoplasma agalactiae. Infect. Immun. 2002, 70, 5612–5621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glew, M.D.; Papazisi, L.; Poumarat, F.; Bergonier, D.; Rosengarten, R.; Citti, C. Characterization of a multigene family undergoing high-frequency DNA rearrangements and coding for abundant variable surface proteins in Mycoplasma agalactiae. Infect. Immun. 2000, 68, 4539–4548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glew, M.D.; Marenda, M.; Rosengarten, R.; Citti, C. Surface diversity in Mycoplasma agalactiae is driven by site-specific DNA inversions within the vpma multigene locus. J. Bacteriol. 2002, 184, 5987–5998. [Google Scholar] [CrossRef] [Green Version]
- Cacciotto, C.; Addis, M.F.; Pagnozzi, D.; Coradduzza, E.; Pittau, M.; Alberti, A. Identification of conserved Mycoplasma agalactiae surface antigens by immunoproteomics. Vet. Immunol. Immunopathol. 2021, 236, 110239. [Google Scholar] [CrossRef]
- Maunsell, F.P.; Woolums, A.R.; Francoz, D.; Rosenbusch, R.F.; Step, D.L.; Wilson, D.J.; Janzen, E.D. Mycoplasma bovis Infections in Cattle. J. Vet. Intern. Med. 2011, 25, 772–783. [Google Scholar] [CrossRef]
- Gelgie, A.E.; Korsa, M.G.; Kerro Dego, O. Mycoplasma bovis Mastitis. Curr. Res. Microb. Sci. 2022, 3, 100123. [Google Scholar] [CrossRef]
- Wise, K.S.; Calcutt, M.J.; Foecking, M.F.; Röske, K.; Madupu, R.; Methé, B.A. Missouri Complete genome sequence of Mycoplasma bovis type strain PG45 (ATCC 25523). Infect. Immun. 2011, 79, 982–983. [Google Scholar] [CrossRef] [Green Version]
- Muramatsu, E.; Sakurai, A.; Kawabe, Y.; Ritsuno, Y.; Sasaki, T.; Tsutsumi, Y.; Isobe, R.; Jinno, R.; Ide, T.; Doi, Y. Periprosthetic joint infection due to Mycoplasma hominis in a multiple sclerosis patient treated with fingolimod. J. Infect. Chemother. 2022, 28, 1672–1676. [Google Scholar] [CrossRef] [PubMed]
- Sadhar, B.; Ko-Keeney, E.; Fornelli, R.; Lipman, S. Mycoplasma Hominis: A Rare Case of Acute Otitis Media and Facial Nerve Paralysis. Ear. Nose. Throat J. 2022. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.L.; Cai, X.; Que, Q.; Zhao, H.; Zhang, K.L.; Lv, S. Mycoplasma hominis meningitis after operative neurosurgery: A case report and review of literature. World J. Clin. cases 2022, 10, 1131–1139. [Google Scholar] [CrossRef] [PubMed]
- Mitsani, D.; Nguyen, M.H.; Silveira, F.P.; Bermudez, C.; Toyoda, Y.; Pasculle, A.W.; Clancy, C.J. Mycoplasma hominis pericarditis in a lung transplant recipient: Review of the literature about an uncommon but important cardiothoracic pathogen. Transpl. Infect. Dis. 2010, 12, 146–150. [Google Scholar] [CrossRef]
- Barykova, Y.A.; Logunov, D.Y.; Shmarov, M.M.; Vinarov, A.Z.; Fiev, D.N.; Vinarova, N.A.; Rakovskaya, I.V.; Baker, P.S.; Shyshynova, I.; Stephenson, A.J.; et al. Association of Mycoplasma hominis infection with prostate cancer. Oncotarget 2011, 2, 289–297. [Google Scholar] [CrossRef] [Green Version]
- Astrauskiene, D.; Griskevicius, A.; Luksiene, R.; Panaviene, V.; Venaliene, J. Chlamydia trachomatis, Ureaplasma urealyticum, and Mycoplasma hominis in sexually intact girls with arthritides. Scand. J. Rheumatol. 2012, 41, 275–279. [Google Scholar] [CrossRef]
- Capoccia, R.; Greub, G.; Baud, D. Ureaplasma urealyticum, Mycoplasma hominis and adverse pregnancy outcomes. Curr. Opin. Infect. Dis. 2013, 26, 231–240. [Google Scholar] [CrossRef]
- Bergin, S.M.; Mendis, S.M.; Young, B.; Izharuddin, E.B. Postoperative Mycoplasma hominis brain abscess: Keep it in mind! BMJ Case Rep. 2017, 2017, bcr2016218022. [Google Scholar] [CrossRef]
- Fiori, P.L.; Diaz, N.; Cocco, A.R.; Rappelli, P.; Dessì, D. Association of Trichomonas vaginalis with its symbiont Mycoplasma hominis synergistically upregulates the in vitro proinflammatory response of human monocytes. Sex. Transm. Infect. 2013, 89, 449–454. [Google Scholar] [CrossRef]
- Goret, J.; Le Roy, C.; Touati, A.; Mesureur, J.; Renaudin, H.; Claverol, S.; Bébéar, C.; Béven, L.; Pereyre, S. Surface lipoproteome of Mycoplasma hominis PG21 and differential expression after contact with human dendritic cells. Future Microbiol. 2016, 11, 179–194. [Google Scholar] [CrossRef]
- Pereyre, S.; Sirand-Pugnet, P.; Beven, L.; Charron, A.; Renaudin, H.; Barré, A.; Avenaud, P.; Jacob, D.; Couloux, A.; Barbe, V.; et al. Life on arginine for Mycoplasma hominis: Clues from its minimal genome and comparison with other human urogenital mycoplasmas. PLoS Genet. 2009, 50, e1000677. [Google Scholar] [CrossRef] [PubMed]
- Waites, K.B.; Talkington, D.F. Mycoplasma pneumoniae and Its Role as a Human Pathogen. Clin. Microbiol. Rev. 2004, 17, 697–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Shao, X.; Dou, X.; Zhang, X.; Wang, Y.; Zhu, C.; Hao, C.; Fan, M.; Ji, W.; Yan, Y. Role of the Mycoplasma pneumoniae/interleukin-8/neutrophil axis in the pathogenesis of pneumonia. PLoS ONE 2016, 11, e0146377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yiwen, C.; Yueyue, W.; Lianmei, Q.; Cuiming, Z.; Xiaoxing, Y. Infection strategies of mycoplasmas: Unraveling the panoply of virulence factors. Virulence 2021, 12, 788–817. [Google Scholar] [CrossRef] [PubMed]
- Marshall, A.J.; Miles, R.J.; Richards, L. The phagocytosis of mycoplasmas. J. Med. Microbiol. 1995, 43, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Himmelreich, R.; Hubert, H.; Plagens, H.; Pirkl, E.; Li, B.C.; Herrmann, R. Complete sequence analysis of the genome of the bacterium Mycoplasma pneumoniae. Nucleic Acids Res. 1996, 24, 4420–4449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Morante, B.; Maes, D.; Sibila, M.; Betlach, A.M.; Sponheim, A.; Canturri, A.; Pieters, M. Improving Mycoplasma hyopneumoniae diagnostic capabilities by harnessing the infection dynamics. Vet. J. 2022, 288, 105877. [Google Scholar] [CrossRef]
- Minion, F.C.; Lefkowitz, E.J.; Madsen, M.L.; Cleary, B.J.; Swartzell, S.M.; Mahairas, G.G. The genome sequence of Mycoplasma hyopneumoniae strain 232, the agent of swine mycoplasmosis. J. Bacteriol. 2004, 186, 7123–7133. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, F.; Castro, P. Differential Cytokine Expression in Natural and Experimental Mastitis Induced byMycoplasma agalactiaein Dairy Goats. Reprod. Domest. Anim. 2014, 50, 159–163. [Google Scholar] [CrossRef]
- Hardy, R.D.; Jafri, H.S.; Olsen, K.; Hatfield, J.; Iglehart, J.; Rogers, B.B.; Patel, P.; Cassell, G.; McCracken, G.H.; Ramilo, O. Mycoplasma pneumoniae Induces Chronic Respiratory Infection, Airway Hyperreactivity, and Pulmonary Inflammation: A Murine Model of Infection-Associated Chronic Reactive Airway Disease. Infect. Immun. 2002, 70, 649–654. [Google Scholar] [CrossRef] [Green Version]
- Caswell, J.L.; Archambault, M. Mycoplasma bovis pneumonia in cattle. Anim. Health Res. Rev. 2007, 8, 161–186. [Google Scholar] [CrossRef] [PubMed]
- Da Rocha Sobrinho, H.M.; Jarach, R.; Da Silva, N.A.; Shio, M.T.; Jancar, S.; Timenetsky, J.; Oliveira, M.A.P.; Dorta, M.L.; Ribeiro-Dias, F. Mycoplasmal lipid-associated membrane proteins and Mycoplasma arthritidis mitogen recognition by serum antibodies from patients with rheumatoid arthritis. Rheumatol. Int. 2011, 31, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Russo, E.; Giudici, F.; Ricci, F.; Scaringi, S.; Nannini, G.; Ficari, F.; Luceri, C.; Niccolai, E.; Baldi, S.; D’ambrosio, M.; et al. Diving into Inflammation: A Pilot Study Exploring the Dynamics of the Immune–Microbiota Axis in Ileal Tissue Layers of Patients with Crohn’s Disease. J. Crohn’s Colitis 2021, 15, 1500–1516. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Li, D.; Paulus, B.; Wilson, I.; Chadwick, V.S. High Prevalence of Mycoplasma pneumoniae in Intestinal Mucosal Biopsies from Patients with Inflammatory Bowel Disease and Controls. Dig. Dis. Sci. 2001, 46, 2529–2535. [Google Scholar] [CrossRef] [PubMed]
- Chu, K.A.; Ou, T.Y.; Hung, W.H.; Sung, J.; Chen, W.; Lin, C.L.; Hung, Y.M.; Wei, J.C.C. Mycoplasma pneumonia Infection Is Associated With an Increased Risk of Systemic Lupus Erythematosus: A Nationwide, Retrospective Cohort Study. Front. Microbiol. 2022, 13, 884. [Google Scholar] [CrossRef] [PubMed]
- Poddighe, D. Extra-pulmonary diseases related to Mycoplasma pneumoniae in children: Recent insights into the pathogenesis. Curr. Opin. Rheumatol. 2018, 30, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Chu, K.A.; Chen, W.; Hsu, C.Y.; Hung, Y.M.; Wei, J.C.C. Increased risk of rheumatoid arthritis among patients with Mycoplasma pneumonia: A nationwide population-based cohort study in Taiwan. PLoS ONE 2019, 14, e0210750. [Google Scholar] [CrossRef]
- Hum, R.M.; Iftikhar, H.; Tang, E.; Ho, P. Septic arthritis caused by Mycoplasma hominis in a patient with systemic lupus erythematosus. BMJ Case Rep. 2022, 15, e247675. [Google Scholar] [CrossRef]
- Bozo, N.; Ravn, C.; Stenz Justesen, U.; Dahlerup Rasmussen, L. Mycoplasma hominis septic arthritis in a patient with hypogammaglobinaemia and rheumatoid arthritis. BMJ Case Rep. 2021, 14, e237798. [Google Scholar] [CrossRef]
- Kawahito, Y.; Ichinose, S.; Sano, H.; Tsubouchi, Y.; Kohno, M.; Yoshikawa, T.; Tokunaga, D.; Hojo, T.; Harasawa, R.; Nakano, T.; et al. Mycoplasma fermentans glycolipid-antigen as a pathogen of rheumatoid arthritis. Biochem. Biophys. Res. Commun. 2008, 369, 561–566. [Google Scholar] [CrossRef]
- Namiki, K.; Goodison, S.; Porvasnik, S.; Allan, R.W.; Iczkowski, K.A.; Urbanek, C.; Reyes, L.; Sakamoto, N.; Rosser, C.J. Persistent Exposure to Mycoplasma Induces Malignant Transformation of Human Prostate Cells. PLoS ONE 2009, 4, e6872. [Google Scholar] [CrossRef]
- Yacoub, E.; Saed Abdul-Wahab, O.M.; Al-Shyarba, M.H.; Ben Abdelmoumen Mardassi, B. The Relationship between Mycoplasmas and Cancer: Is It Fact or Fiction ? Narrative Review and Update on the Situation. J. Oncol. 2021, 2021, 9986550. [Google Scholar] [CrossRef] [PubMed]
- Saadat, S.; Karami, P.; Jafari, M.; Kholoujini, M.; Tehrani, Z.R.; Mohammadi, Y.; Alikhani, M.Y. The silent presence of Mycoplasma hominis in patients with prostate cancer. Pathog. Dis. 2020, 78, ftaa037. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Wahab, O.M.S.; Al-Shyarba, M.H.; Mardassi, B.B.A.; Sassi, N.; Al Fayi, M.S.S.; Otifi, H.; Al Murea, A.H.; Mlik, B.; Yacoub, E. Molecular detection of urogenital mollicutes in patients with invasive malignant prostate tumor. Infect. Agent. Cancer 2021, 16, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wear, D.J.; Lo, S.-C. Mycoplasmal infections alter gene expression in cultured human prostatic and cervical epithelial cells. FEMS Immunol. Med. Microbiol. 2000, 27, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.; Samwel, K.; Kahesa, C.; Mwaiselage, J.; West, J.T.; Wood, C.; Angeletti, P.C. Mycoplasma Co-Infection Is Associated with Cervical Cancer Risk. Cancers 2020, 12, 1093. [Google Scholar] [CrossRef]
- Tsai, S.; Wear, D.J.; Shih, J.W.K.; Lo, S.C. Mycoplasmas and oncogenesis: Persistent infection and multistage malignant transformation. Proc. Natl. Acad. Sci. USA 1995, 92, 10197–10201. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Li, H.; Chen, W.; Yao, X.; Xing, Y.; Wang, X.; Zhong, J.; Meng, G. Mycoplasma hyorhinis activates the NLRP3 inflammasome and promotes migration and invasion of gastric chancer cells. PLoS ONE 2013, 8, e77955. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cacciotto, C.; Alberti, A. Eating the Enemy: Mycoplasma Strategies to Evade Neutrophil Extracellular Traps (NETs) Promoting Bacterial Nucleotides Uptake and Inflammatory Damage. Int. J. Mol. Sci. 2022, 23, 15030. https://doi.org/10.3390/ijms232315030
Cacciotto C, Alberti A. Eating the Enemy: Mycoplasma Strategies to Evade Neutrophil Extracellular Traps (NETs) Promoting Bacterial Nucleotides Uptake and Inflammatory Damage. International Journal of Molecular Sciences. 2022; 23(23):15030. https://doi.org/10.3390/ijms232315030
Chicago/Turabian StyleCacciotto, Carla, and Alberto Alberti. 2022. "Eating the Enemy: Mycoplasma Strategies to Evade Neutrophil Extracellular Traps (NETs) Promoting Bacterial Nucleotides Uptake and Inflammatory Damage" International Journal of Molecular Sciences 23, no. 23: 15030. https://doi.org/10.3390/ijms232315030