
Immune Responses to COVID-19 Vaccines in Patients with Chronic Kidney Disease and Lead Exposure
 , and
, and
Abstract
:
1. Introduction
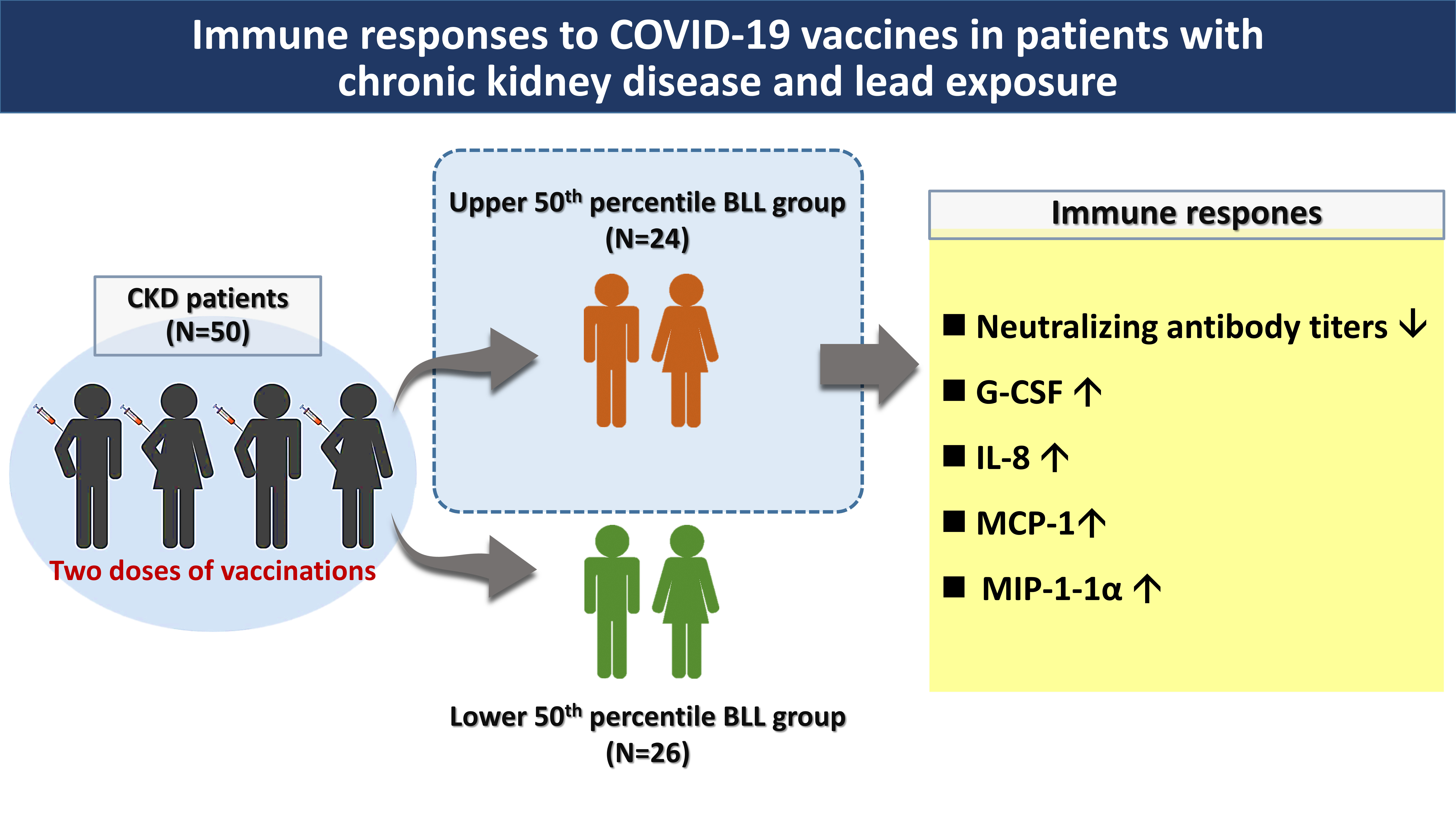
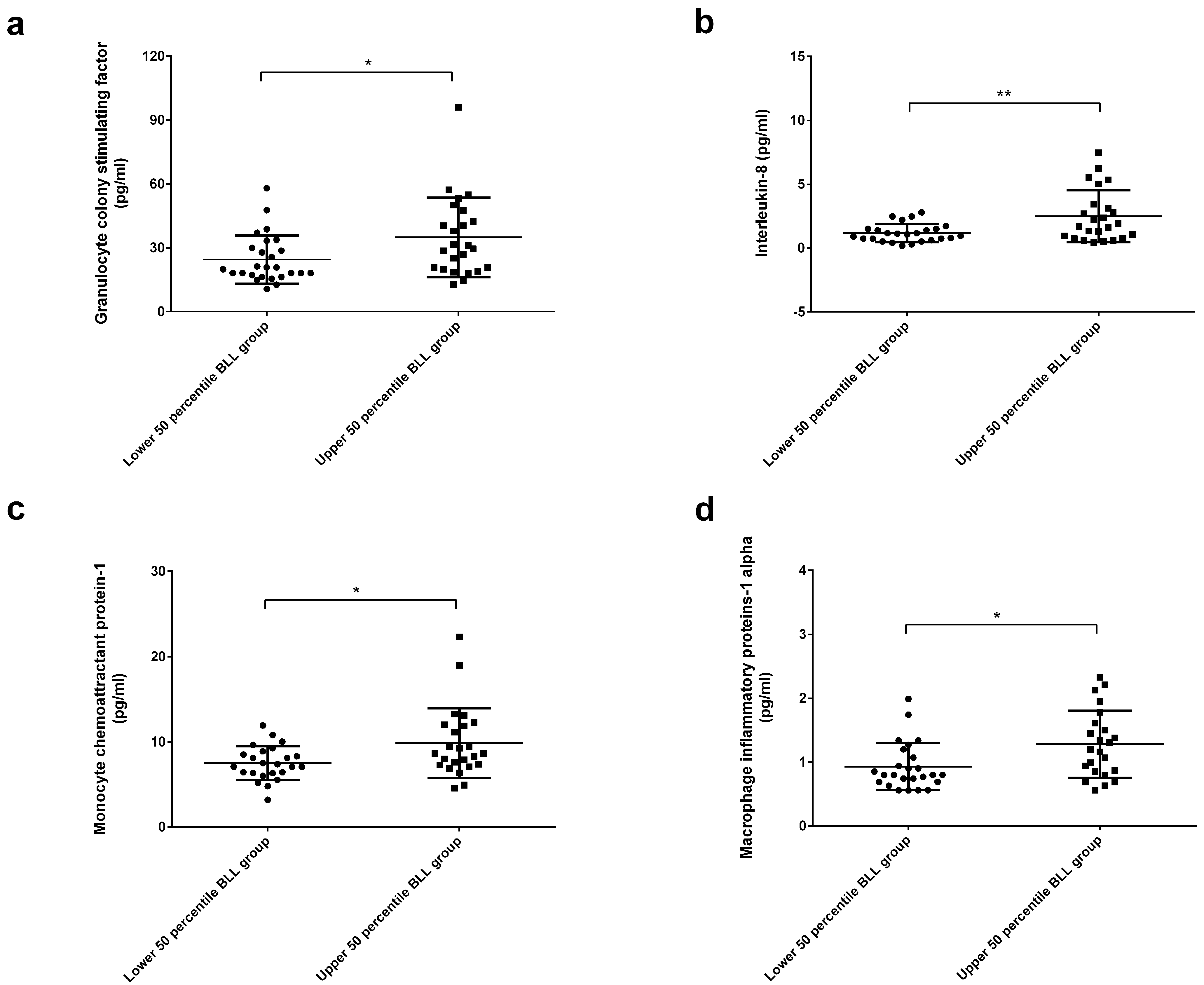
2. Results
3. Discussion
4. Materials and Methods
4.1. Inclusion and Exclusion Criteria
4.2. Study Design
4.3. Measurement of Neutralizing Antibodies against COVID-19
4.4. Blood Lead Determination Using Graphite Furnace Atomic Absorption Spectrometry
4.5. Cytokine Measurements Using Multiplex Immunoassay
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and efficacy of the chadox1 ncov-19 vaccine (azd1222) against SARS-CoV-2: An interim analysis of four randomised controlled trials in brazil, south africa, and the uk. Lancet 2021, 397, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Marc, G.P.; Moreira, E.D.; Zerbini, C.; et al. Safety and efficacy of the bnt162b2 mrna COVID-19 vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and safety of the mrna-1273 SARS-CoV-2 vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Lee, A.; Wong, S.Y.; Chai, L.Y.A.; Lee, S.C.; Lee, M.X.; Muthiah, M.D.; Tay, S.H.; Teo, C.B.; Tan, B.K.J.; Chan, Y.H.; et al. Efficacy of COVID-19 vaccines in immunocompromised patients: Systematic review and meta-analysis. BMJ 2022, 376, e068632. [Google Scholar] [CrossRef] [PubMed]
- Yen, J.S.; Wang, I.K.; Yen, T.H. COVID-19 vaccination and dialysis patients: Why the variable response. QJM 2021, 114, 440–444. [Google Scholar] [CrossRef]
- Anand, S.; Montez-Rath, M.E.; Han, J.; Garcia, P.; Cadden, L.; Hunsader, P.; Kerschmann, R.; Beyer, P.; Dittrich, M.; Block, G.A.; et al. Antibody response to COVID-19 vaccination in patients receiving dialysis. J. Am. Soc. Nephrol. 2021, 32, 2435–2438. [Google Scholar] [CrossRef]
- Metryka, E.; Chibowska, K.; Gutowska, I.; Falkowska, A.; Kupnicka, P.; Barczak, K.; Chlubek, D.; Baranowska-Bosiacka, I. Lead (pb) exposure enhances expression of factors associated with inflammation. Int. J. Mol. Sci. 2018, 19, 1813. [Google Scholar] [CrossRef] [Green Version]
- Skalny, A.V.; Lima, T.R.R.; Ke, T.; Zhou, J.C.; Bornhorst, J.; Alekseenko, S.I.; Aaseth, J.; Anesti, O.; Sarigiannis, D.A.; Tsatsakis, A.; et al. Toxic metal exposure as a possible risk factor for COVID-19 and other respiratory infectious diseases. Food Chem. Toxicol. 2020, 146, 111809. [Google Scholar] [CrossRef]
- Sahin, M.; Karayakar, F.; Koksal, A.R.; Yetim, A.; İyisoy, M.S.; Şen, İ.; Alkım, H.; Alkım, C.; Colak, T. Changes in liver tissue trace element concentrations during hepatitis b viral infection treatment. Biol. Trace Elem. Res. 2019, 188, 245–250. [Google Scholar] [CrossRef]
- Rashed, M.N. The role of trace elements on hepatitis virus infections: A review. J. Trace Elem. Med. Biol. 2011, 25, 181–187. [Google Scholar] [CrossRef]
- Syed-Ahmed, M.; Narayanan, M. Immune dysfunction and risk of infection in chronic kidney disease. Adv. Chronic. Kidney Dis. 2019, 26, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, L.; Silvestroni, A.; Martino, M.G.; Gagliardi, T.; Corfiati, M.; Soleo, L. Evaluation of peripheral blood neutrophil leucocytes in lead-exposed workers. Int. Arch. Occup. Environ. Health 2006, 79, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Flohé, S.B.; Brüggemann, J.; Herder, C.; Goebel, C.; Kolb, H. Enhanced proinflammatory response to endotoxin after priming of macrophages with lead ions. J. Leukoc. Biol. 2002, 71, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Kasten-Jolly, J.; Lawrence, D.A. Lead modulation of macrophages causes multiorgan detrimental health effects. J. Biochem. Mol. Toxicol. 2014, 28, 355–372. [Google Scholar] [CrossRef] [PubMed]
- Heo, Y.; Lee, B.K.; Ahn, K.D.; Lawrence, D.A. Serum ige elevation correlates with blood lead levels in battery manufacturing workers. Hum. Exp. Toxicol. 2004, 23, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Karmaus, W.; Brooks, K.R.; Nebe, T.; Witten, J.; Obi-Osius, N.; Kruse, H. Immune function biomarkers in children exposed to lead and organochlorine compounds: A cross-sectional study. Environ. Health 2005, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Mishra, K.P.; Chauhan, U.K.; Naik, S. Effect of lead exposure on serum immunoglobulins and reactive nitrogen and oxygen intermediate. Hum. Exp. Toxicol. 2006, 25, 661–665. [Google Scholar] [CrossRef]
- Mishra, K.P.; Rani, R.; Yadav, V.S.; Naik, S. Effect of lead exposure on lymphocyte subsets and activation markers. Immunopharmacol. Immunotoxicol. 2010, 32, 446–449. [Google Scholar] [CrossRef]
- Gao, D.; Mondal, T.K.; Lawrence, D.A. Lead effects on development and function of bone marrow-derived dendritic cells promote th2 immune responses. Toxicol. Appl. Pharmacol. 2007, 222, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Dobrakowski, M.; Boroń, M.; Czuba, Z.P.; Kasperczyk, A.; Machoń-Grecka, A.; Kasperczyk, S. Cytokines related to three major types of cell-mediated immunity in short- and long-term exposures to lead compounds. J. Immunotoxicol. 2016, 13, 770–774. [Google Scholar] [CrossRef]
- Sadarangani, M.; Marchant, A.; Kollmann, T.R. Immunological mechanisms of vaccine-induced protection against COVID-19 in humans. Nat. Rev. Immunol. 2021, 21, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Lutz, P.M.; Bauer, S.; Gale, N.L.; Hewett, J.; Phillips, P.E.; Looney, F.M.; Bengsch, H. Immunity in children with exposure to environmental lead: Ii. Effects on humoral immunity. Environ. Geochem. Health 1994, 16, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Xu, X.; Zeng, X.; Xu, L.; Zeng, Z.; Huo, X. Decreased vaccine antibody titers following exposure to multiple metals and metalloids in e-waste-exposed preschool children. Environ. Pollut. 2017, 220, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Chen, X.; Zhang, J.; Guo, P.; Fu, T.; Dai, Y.; Lin, S.L.; Huo, X. Decreased blood hepatitis b surface antibody levels linked to e-waste lead exposure in preschool children. J. Hazard. Mater. 2015, 298, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Di Lenardo, T.Z.; Ward, B.J.; Pillet, S.; Mann, K.; Bornman, R.; Obida, M.; Chevrier, J. Exposure to lead and vaccine-specific igg titers in south african children participating in the venda health examination of mothers, babies and their environment (vhembe): A longitudinal study. Environ. Res. 2020, 180, 108794. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.M.; Lee, N.Y.; Lin, C.H.; Hsu, Y.S.; Chang, Y.C.; Chung, M.Y.; Lee, Y.F.; Tseng, W.P.; Wu, J.L.; Chen, S.Y.; et al. Immunogenicity and safety of homologous and heterologous chadox1-s and mrna-1273 vaccinations in healthy adults in taiwan. J. Clin. Virol. 2022, 150, 105156. [Google Scholar] [CrossRef]
- Bendall, L.J.; Bradstock, K.F. G-csf: From granulopoietic stimulant to bone marrow stem cell mobilizing agent. Cytokine Growth Factor Rev. 2014, 25, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Di Lorenzo, L.; Vacca, A.; Corfiati, M.; Lovreglio, P.; Soleo, L. Evaluation of tumor necrosis factor-alpha and granulocyte colony-stimulating factor serum levels in lead-exposed smoker workers. Int. J. Immunopathol. Pharmacol. 2007, 20, 239–247. [Google Scholar] [CrossRef]
- Waugh, D.J.; Wilson, C. The interleukin-8 pathway in cancer. Clin. Cancer Res. 2008, 14, 6735–6741. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.C.; Wei, P.L.; Tsai, Y.T.; Wong, J.H.; Chang, C.M.; Wang, J.Y.; Hou, M.F.; Lee, Y.C.; Chuang, H.Y.; Chang, W.C. Pb2+ induced il-8 gene expression by extracellular signal-regulated kinases and the transcription factor, activator protein 1, in human gastric carcinoma cells. Environ. Toxicol. 2015, 30, 315–322. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, X.; Fu, Y.; Yang, H. Leptin and il-8: Two novel cytokines screened out in childhood lead exposure. Toxicol. Lett. 2014, 227, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Dobrakowski, M.; Kasperczyk, A.; Pawlas, N.; Birkner, E.; Hudziec, E.; Chwalińska, E.; Kasperczyk, S. Association between subchronic and chronic lead exposure and levels of antioxidants and chemokines. Int. Arch. Occup. Environ. Health 2016, 89, 1077–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (mcp-1): An overview. J. Interferon. Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Kumawat, K.L.; Kaushik, D.K.; Goswami, P.; Basu, A. Acute exposure to lead acetate activates microglia and induces subsequent bystander neuronal death via caspase-3 activation. Neurotoxicology 2014, 41, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Soliman, M.M.; Baiomy, A.A.; Yassin, M.H. Molecular and histopathological study on the ameliorative effects of curcumin against lead acetate-induced hepatotoxicity and nephrototoxicity in wistar rats. Biol. Trace Elem. Res. 2015, 167, 91–102. [Google Scholar] [CrossRef]
- Maurer, M.; von Stebut, E. Macrophage inflammatory protein-1. Int. J. Biochem. Cell Biol. 2004, 36, 1882–1886. [Google Scholar] [CrossRef]
- Gillis, B.S.; Arbieva, Z.; Gavin, I.M. Analysis of lead toxicity in human cells. BMC Genomics 2012, 13, 344. [Google Scholar] [CrossRef] [Green Version]
- Machoń-Grecka, A.; Dobrakowski, M.; Kasperczyk, A.; Birkner, E.; Pryzwan, T.; Kasperczyk, S. The effect of subacute lead exposure on selected blood inflammatory biomarkers and angiogenetic factors. J. Occup. Health 2018, 60, 369–375. [Google Scholar] [CrossRef] [Green Version]
- de Alwis, R.; Chen, S.; Gan, E.S.; Ooi, E.E. Impact of immune enhancement on COVID-19 polyclonal hyperimmune globulin therapy and vaccine development. EBioMedicine 2020, 55, 102768. [Google Scholar] [CrossRef]
- Ewer, K.J.; Barrett, J.R.; Belij-Rammerstorfer, S.; Sharpe, H.; Makinson, R.; Morter, R.; Flaxman, A.; Wright, D.; Bellamy, D.; Bittaye, M.; et al. T cell and antibody responses induced by a single dose of chadox1 ncov-19 (azd1222) vaccine in a phase 1/2 clinical trial. Nat. Med. 2021, 27, 270–278. [Google Scholar] [CrossRef]
- Anderson, E.J.; Rouphael, N.G.; Widge, A.T.; Jackson, L.A.; Roberts, P.C.; Makhene, M.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; Pruijssers, A.J.; et al. Safety and immunogenicity of SARS-CoV-2 mrna-1273 vaccine in older adults. N. Engl. J. Med. 2020, 383, 2427–2438. [Google Scholar] [CrossRef] [PubMed]
- Bergamaschi, C.; Terpos, E.; Rosati, M.; Angel, M.; Bear, J.; Stellas, D.; Karaliota, S.; Apostolakou, F.; Bagratuni, T.; Patseas, D.; et al. Systemic il-15, ifn-γ, and ip-10/cxcl10 signature associated with effective immune response to SARS-CoV-2 in bnt162b2 mrna vaccine recipients. Cell Rep. 2021, 36, 109504. [Google Scholar] [CrossRef] [PubMed]
- Tahtinen, S.; Tong, A.J.; Himmels, P.; Oh, J.; Paler-Martinez, A.; Kim, L.; Wichner, S.; Oei, Y.; McCarron, M.J.; Freund, E.C.; et al. Il-1 and il-1ra are key regulators of the inflammatory response to rna vaccines. Nat. Immunol. 2022, 23, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.W.; Ning, H.C.; Huang, Y.C.; Chiang, Y.S.; Chuang, C.W.; Wang, I.K.; Fan, N.C.; Weng, C.H.; Huang, W.H.; Hsu, C.W.; et al. Trend in blood lead levels in taiwanese adults 2005–2017. PLoS ONE 2021, 16, e0260744. [Google Scholar] [CrossRef] [PubMed]
- Shefa, S.T.; Heroux, P. Both physiology and epidemiology support zero tolerable blood lead levels. Toxicol. Lett. 2017, 280, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Mohsin, M.; Mahmud, S. Omicron SARS-CoV-2 variant of concern: A review on its transmissibility, immune evasion, reinfection, and severity. Medicine 2022, 101, e29165. [Google Scholar] [CrossRef] [PubMed]
- Yen, J.S.; Wang, I.K.; Liang, C.C.; Fu, J.F.; Hou, Y.C.; Chang, C.C.; Gu, P.W.; Tsai, K.F.; Weng, C.H.; Huang, W.H.; et al. Cytokine changes in fatal cases of paraquat poisoning. Am. J. Transl. Res. 2021, 13, 11571–11584. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Total (n = 50) | Patients with Upper 50th Percentile BLL (n = 24) | Patients with Lower 50th Percentile BLL (n = 26) | p Value |
|---|---|---|---|---|
| Demographics | ||||
| Male, n (%) | 32 (64.0) | 18 (75.0) | 14 (53.8) | 0.119 |
| Age, year | 65.9 ± 11.8 | 68.1 ± 9.1 | 63.8 ± 12.8 | 0.180 |
| Body mass index, kg/m2 | 26.1 ± 4.4 | 26.2 ± 3.3 | 26.0 ± 5.3 | 0.199 |
| Smoking habit, n (%) | 10 (20.0) | 7 (29.2) | 3 (11.5) | 0.119 |
| Alcohol consumption, n (%) | 8 (16.0) | 5 (20.8) | 3 (11.5) | 0.370 |
| Betel nut usage, n (%) | 1 (2.0) | 1 (4.2) | 0 (0) | 0.293 |
| Chronic kidney disease staging | ||||
| Stage 1, n (%) | 13 (26.0) | 3 (12.5) | 10 (38.5) | 0.054 |
| Stage 2, n (%) | 10 (20.0) | 5 (20.8) | 5 (19.2) | 1.000 |
| Stage 3, n (%) | 11 (22.0) | 5 (20.8) | 6 (23.1) | 1.000 |
| Stage 4, n (%) | 4 (8.0) | 2 (8.3) | 2 (7.7) | 1.000 |
| Stage 5, n (%) | 12 (24.0) | 9 (37.5) | 3 (11.5) | 0.047 * |
| Diabetic mellitus, n (%) | 20 (40.0) | 12 (50.0) | 8 (30.8) | 0.166 |
| Hypertension, n (%) | 40 (80.0) | 21 (87.5) | 19 (73.1) | 0.203 |
| Biopsy-proved chronic glomerulonephritis, n (%) | 4 (8.0) | 1 (4.2) | 3 (11.5) | 0.337 |
| Biopsy-proved tubulointerstitial nephritis | 0 (0) | 0 (0) | 0 (0) | 1.000 |
| Polycystic kidney disease, n (%) | 0 (0) | 0 (0) | 0 (0) | 1.000 |
| Gouty arthritis, n (%) | 11 (22.0) | 6 (25.0) | 5 (19.2) | 0.623 |
| Dyslipidemia, n (%) | 20 (40.0) | 7 (29.2) | 13 (50.0) | 0.133 |
| Malignancy, n (%) | 3 (6.0) | 1 (4.2) | 2 (7.7) | 0.600 |
| Solid organ transplant, n (%) | 0 (0) | 0 (0) | 0 (0) | 1.000 |
| Immunosuppressive therapy, n (%) | 0 (0) | 0 (0) | 0 (0) | 1.000 |
| Variable | Total (n = 50) | Patients with Upper 50th Percentile BLL (n = 24) | Patients with Lower 50th Percentile BLL (n = 26) | p Value |
|---|---|---|---|---|
| Blood lead level, μg/dL | 1.9 ± 1.9 | 3.1 ± 2.3 | 0.9 ± 0.2 | <0.001 *** |
| Hemogram | ||||
| Hemoglobin, g/dL | 11.8 ± 2.7 | 11.5 ± 2.9 | 12.0 ± 2.6 | 0.582 |
| Hematocrit, % | 36.0 ± 8.2 | 35.6 ± 9.2 | 36.4 ± 7.3 | 0.776 |
| Red blood cell count, 106/μL | 4.2 ± 0.8 | 4.1 ± 1.1 | 4.2 ± 0.7 | 0.822 |
| Mean corpuscular volume, fL | 90.0 ± 7.2 | 91.6 ± 5.2 | 89.4 ± 8.0 | 0.577 |
| Platelet count, 103/μL | 216.7 ± 68.5 | 197.6 ± 38.2 | 224.0 ± 77.2 | 0.481 |
| White blood cell count, 103/μL | 8.6 ± 8.4 | 6.8 ± 1.6 | 9.2 ± 9.9 | 0.602 |
| Biochemistry | ||||
| Blood urea nitrogen, mg/dL | 32.6 ± 23.3 | 39.2 ± 23.3 | 27.8 ± 22.6 | 0.115 |
| Creatinine, mg/dL | 3.2 ± 3.8 | 4.7 ± 4.3 | 1.9 ± 2.8 | 0.011 * |
| Albumin, g/dL | 3.9 ± 0.3 | 3.9 ± 0.3 | 3.9 ± 0.3 | 0.826 |
| Uric acid, mg/dL | 6.6 ± 2.0 | 6.6 ± 1.9 | 6.5 ± 2.1 | 0.838 |
| Alanine aminotransferase, U/L | 22.3 ± 11.1 | 19.8 ± 7.8 | 24.2 ± 12.9 | 0.252 |
| Total cholesterol, mg/dL | 200.3 ± 38.7 | 183.5 ± 31.2 | 210.0 ± 40.0 | 0.069 |
| High-density lipoprotein cholesterol, mg/dL | 53.7 ± 15.3 | 50.7 ± 12.7 | 55.4 ± 16.7 | 0.432 |
| Low-density lipoprotein cholesterol, mg/dL | 115.8 ± 34.3 | 111.0 ± 27.2 | 119.3 ± 39.0 | 0.502 |
| Triglyceride, mg/dL | 142.7 ± 91.8 | 119.8 ± 44.2 | 158.3 ± 112.2 | 0.252 |
| Fasting blood sugar, mg/dL | 115.6 ± 33.8 | 119.5 ± 30.4 | 112.9 ± 36.6 | 0.567 |
| Glycated hemoglobin, % | 6.3 ± 0.9 | 6.1 ± 0.9 | 6.7 ± 0.8 | 0.259 |
| Variable | Total (n = 50) | Patients with Upper 50th Percentile BLL (n = 24) | Patients with Lower 50th Percentile BLL (n = 26) | p Value |
|---|---|---|---|---|
| Time elapsed between second vaccination and blood test (day) | 26.1 (16.4) | 30.8 ± 12.6 | 21.8 ± 18.5 | 0.051 |
| Type of vaccine | 0.031 * | |||
| ChAdOx1-S, n (%) | 17 (34.0) | 12 (50.0) | 5 (19.2) | |
| mRNA-1273, n (%) | 32 (64.0) | 11 (45.8) | 21 (80.8) | |
| BNT162b2, n (%) | 0 (0) | 0 (0) | 0 (0) | |
| MVC-COV1901, n (%) | 1 (2.0) | 1 (4.2) | 0 (0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yen, J.-S.; Wu, Y.-C.; Yen, J.-C.; Wang, I.-K.; Fu, J.-F.; Cheng, C.-M.; Yen, T.-H. Immune Responses to COVID-19 Vaccines in Patients with Chronic Kidney Disease and Lead Exposure. Int. J. Mol. Sci. 2022, 23, 15003. https://doi.org/10.3390/ijms232315003
Yen J-S, Wu Y-C, Yen J-C, Wang I-K, Fu J-F, Cheng C-M, Yen T-H. Immune Responses to COVID-19 Vaccines in Patients with Chronic Kidney Disease and Lead Exposure. International Journal of Molecular Sciences. 2022; 23(23):15003. https://doi.org/10.3390/ijms232315003
Chicago/Turabian StyleYen, Ju-Shao, Yao-Cheng Wu, Ju-Ching Yen, I-Kuan Wang, Jen-Fen Fu, Chao-Min Cheng, and Tzung-Hai Yen. 2022. "Immune Responses to COVID-19 Vaccines in Patients with Chronic Kidney Disease and Lead Exposure" International Journal of Molecular Sciences 23, no. 23: 15003. https://doi.org/10.3390/ijms232315003