

The Role of Adipokines in Inflammatory Mechanisms of Obesity
 , , ,
, , ,  and
and
Abstract
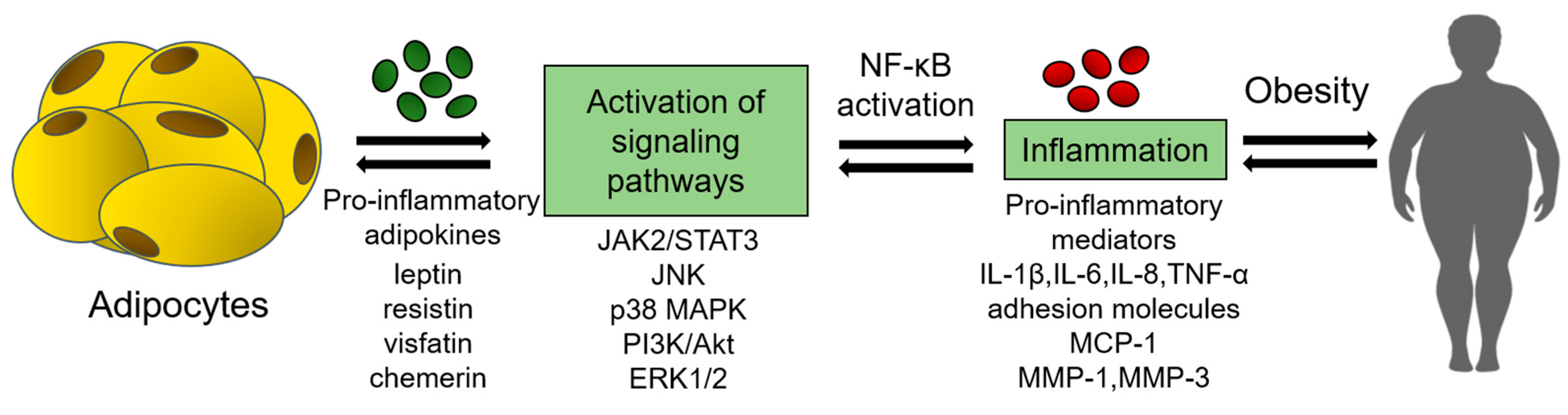
:1. Introduction
2. Pro-Inflammatory Adipokines in Obesity and Its Complications
2.1. Leptin
2.2. Resistin
2.3. Visfatin
2.4. Chemerin
2.5. DPP4
3. Anti-Inflammatory Adipokines in Obesity and Its Complications
3.1. Adiponectin
3.2. Omentin
3.3. Isthmin 1
3.4. Nesfatin 1
4. Adipokines in Clinical Trials
4.1. Adipokines as Prognostic Markers
4.2. Adipokines as Therapeutic Targets

{kind=link}
| Intervention | Study Design | Effects |
|---|---|---|
| Metformin [143,144] | Individuals with metabolic syndrome without restriction in age treated with a dose of 500–3000 mg/day for 3–12 months (several studies included in systematic review) | ↑ adiponectin ↓ leptin ↓ resistin ↓ visfatin |
| Dapagliflozin [146] | Patients with type 2 diabetes and class 3 obesity, 10 mg/day over a 12-month treatment period | ↓ leptin n/a adiponectin |
| Vildagliptin [147] | Women with obesity and type 2 diabetes, 50 mg twice/day over a 30-day treatment period | ↓ DPP4 ↑ adiponectin |
| Pioglitazone [148] | Patients with nonalcoholic steatohepatitis, 45 mg/day with 6 months of administration | ↑ adiponectin n/a leptin |
| PUFAs [150] | Patients with type 2 diabetes, >8 weeks of treatment | ↑ adiponectin ↓ leptin |
| Lactobacillus probiotic [151] | Subjects with BMI of 25–30 kg/m2 on a diet including 4 × 109 colony-forming units of L. plantarum for 12 weeks | ↓ leptin |
| Statins [153] | Studies included in systematic review, statin administration for >2 weeks at the following doses: atorvastatin, 10–80 mg/day; simvastatin, 10–40 mg/day; rosuvastatin, 2.5–10 mg/day; pravastatin, 10–40 mg/day; pitavastatin, 2 mg/day | ↑ adiponectin |
| Telmisartan [156] | Patients with arterial hypertension combined with obesity, 80 mg/day, 12-week treatment period | ↑ adiponectin |
| Perindopril [157] | Overweight or obese patients with hypertension, 10 mg/day, 24-week treatment period | ↓ leptin |
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- One Billion People Globally Estimated to Be Living with Obesity by 2030|World Obesity Federation. Available online: https://www.worldobesity.org/news/one-billion-people-globally-estimated-to-be-living-with-obesity-by-2030 (accessed on 17 November 2022).
- Wei, W.; Zhang, X.; Zhou, B.; Ge, B.; Tian, J.; Chen, J. Effects of Female Obesity on Conception, Pregnancy and the Health of Offspring. Front. Endocrinol. 2022, 13, 949228. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Shi, Z.; Ji, X.; Zhang, W.; Luan, J.; Zahr, T.; Qiang, L. Adipokines, Adiposity, and Atherosclerosis. Cell. Mol. Life Sci. 2022, 79, 272. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.; Pessin, J.E. Adipokines Mediate Inflammation and Insulin Resistance. Front. Endocrinol. 2013, 4, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fasshauer, M.; Blüher, M. Adipokines in Health and Disease. Trends Pharmacol. Sci. 2015, 36, 461–470. [Google Scholar] [CrossRef]
- Metz, S.; Huang, L.O.; Kilpeläinen, T.O. Genetic Variation, Adipokines, and Cardiometabolic Disease. Curr. Opin. Pharmacol. 2020, 52, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Farkhondeh, T.; Llorens, S.; Pourbagher-Shahri, A.M.; Ashrafizadeh, M.; Talebi, M.; Shakibaei, M.; Samarghandian, S. An Overview of the Role of Adipokines in Cardiometabolic Diseases. Molecules 2020, 25, 5218. [Google Scholar] [CrossRef]
- Schleinitz, D.; Böttcher, Y.; Blüher, M.; Kovacs, P. The Genetics of Fat Distribution. Diabetologia 2014, 57, 1276–1286. [Google Scholar] [CrossRef] [PubMed]
- Gastaldelli, A.; Miyazaki, Y.; Pettiti, M.; Matsuda, M.; Mahankali, S.; Santini, E.; DeFronzo, R.A.; Ferrannini, E. Metabolic Effects of Visceral Fat Accumulation in Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2002, 87, 5098–5103. [Google Scholar] [CrossRef] [Green Version]
- Lehr, S.; Hartwig, S.; Sell, H. Adipokines: A Treasure Trove for the Discovery of Biomarkers for Metabolic Disorders. Proteom. Clin. Appl. 2012, 6, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Guilherme, A.; Virbasius, J.V.; Puri, V.; Czech, M.P. Adipocyte Dysfunctions Linking Obesity to Insulin Resistance and Type 2 Diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 367. [Google Scholar] [CrossRef] [PubMed]
- Zorena, K.; Jachimowicz-Duda, O.; Ślęzak, D.; Robakowska, M.; Mrugacz, M. Adipokines and Obesity. Potential Link to Metabolic Disorders and Chronic Complications. Int. J. Mol. Sci. 2020, 21, 3570. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in Inflammation and Metabolic Disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.B. The Complex Role of Adipokines in Obesity, Inflammation, and Autoimmunity. Clin. Sci. 2021, 135, 731. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W. Obesity Is Associated with Macrophage Accumulation in Adipose Tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Leal, V.D.O.; Mafra, D. Adipokines in Obesity. Clin. Chim. Acta 2013, 419, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.L.; Yang, Z.; Yang, S.S. Roles of Adipokines in Digestive Diseases: Markers of Inflammation, Metabolic Alteration and Disease Progression. Int. J. Mol. Sci. 2020, 21, 8308. [Google Scholar] [CrossRef]
- Hsu, P.S.; Wu, C.S.; Chang, J.F.; Lin, W.N. Leptin Promotes CPLA2 Gene Expression through Activation of the MAPK/NF-ΚB/P300 Cascade. Int. J. Mol. Sci. 2015, 16, 27640. [Google Scholar] [CrossRef] [Green Version]
- Yao, J.; Wu, D.; Qiu, Y. Adipose Tissue Macrophage in Obesity-Associated Metabolic Diseases. Front. Immunol. 2022, 13, 977485. [Google Scholar] [CrossRef]
- Ohashi, K.; Parker, J.L.; Ouchi, N.; Higuchi, A.; Vita, J.A.; Gokce, N.; Pedersen, A.A.; Kalthoff, C.; Tullin, S.; Sams, A.; et al. Adiponectin Promotes Macrophage Polarization toward an Anti-Inflammatory Phenotype. J. Biol. Chem. 2010, 285, 6153. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Pérez, A.; Vilariño-García, T.; Fernández-Riejos, P.; Martín-González, J.; Segura-Egea, J.J.; Sánchez-Margalet, V. Role of Leptin as a Link between Metabolism and the Immune System. Cytokine Growth Factor Rev. 2017, 35, 71–84. [Google Scholar] [CrossRef]
- Unamuno, X.; Gómez-Ambrosi, J.; Rodríguez, A.; Becerril, S.; Frühbeck, G.; Catalán, V. Adipokine Dysregulation and Adipose Tissue Inflammation in Human Obesity. Eur. J. Clin. Investig. 2018, 48, e12997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Z.; Chen, Y.; Upender, R.P. Sleep Disturbance and Metabolic Dysfunction: The Roles of Adipokines. Int. J. Mol. Sci. 2022, 23, 1706. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Margalet, V.; Martín-Romero, C.; Santos-Alvarez, J.; Goberna, R.; Najib, S.; Gonzalez-Yanes, C. Role of Leptin as an Immunomodulator of Blood Mononuclear Cells: Mechanisms of Action. Clin. Exp. Immunol. 2003, 133, 11. [Google Scholar] [CrossRef]
- Roca-Rodríguez, M.D.M.; Ramos-García, P.; López-Tinoco, C.; Aguilar-Diosdado, M. Significance of Serum-Plasma Leptin Profile during Pregnancy in Gestational Diabetes Mellitus: A Systematic Review and Meta-Analysis. J. Clin. Med. 2022, 11, 2433. [Google Scholar] [CrossRef]
- Denver, R.J.; Bonett, R.M.; Boorse, G.C. Evolution of Leptin Structure and Function. Neuroendocrinology 2011, 94, 21–38. [Google Scholar] [CrossRef] [PubMed]
- Morris, D.L.; Rui, L. Recent Advances in Understanding Leptin Signaling and Leptin Resistance. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E1247–E1259. [Google Scholar] [CrossRef] [Green Version]
- Sinha, M.K.; Ohannesian, J.P.; Heiman, M.L.; Kriauciunas, A.; Stephens, T.W.; Magosin, S.; Marco, C.; Caro, J.F. Nocturnal Rise of Leptin in Lean, Obese, and Non-Insulin-Dependent Diabetes Mellitus Subjects. J. Clin. Investig. 1996, 97, 1344. [Google Scholar] [CrossRef]
- Wada, N.; Hirako, S.; Takenoya, F.; Kageyama, H.; Okabe, M.; Shioda, S. Leptin and Its Receptors. J. Chem. Neuroanat. 2014, 61–62, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Münzberg, H.; Morrison, C.D. Structure, Production and Signaling of Leptin. Metabolism 2015, 64, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Kargasheh, F.B.; Ansaripour, S.; Borumandnia, N.; Moradi, N.; Zandieh, Z.; Maleki, M.; Mokhtar, S.; Karimi, A.; Fatemi, F.; Kheirollahi, A.; et al. Association of Leptin G2548A and Leptin Receptor Q223R Polymorphisms and Their Serum Levels with Infertility and Recurrent Pregnancy Loss in Iranian Women with Polycystic Ovary Syndrome. PLoS ONE 2021, 16, e0255920. [Google Scholar] [CrossRef]
- Li, S.; Li, X. Leptin in Normal Physiology and Leptin Resistance. Sci. Bull. 2016, 61, 1480–1488. [Google Scholar] [CrossRef] [Green Version]
- Socol, C.T.; Chira, A.; Martinez-Sanchez, M.A.; Nuñez-Sanchez, M.A.; Maerescu, C.M.; Mierlita, D.; Rusu, A.V.; Ruiz-Alcaraz, A.J.; Trif, M.; Ramos-Molina, B. Leptin Signaling in Obesity and Colorectal Cancer. Int. J. Mol. Sci. 2022, 23, 4713. [Google Scholar] [CrossRef] [PubMed]
- Aleffi, S.; Petrai, I.; Bertolani, C.; Parola, M.; Colombatto, S.; Novo, E.; Vizzutti, F.; Anania, F.A.; Milani, S.; Rombouts, K.; et al. Upregulation of Proinflammatory and Proangiogenic Cytokines by Leptin in Human Hepatic Stellate Cells. Hepatology 2005, 42, 1339–1348. [Google Scholar] [CrossRef] [PubMed]
- Rajesh, Y.; Sarkar, D. Association of Adipose Tissue and Adipokines with Development of Obesity-Induced Liver Cancer. Int. J. Mol. Sci. 2021, 22, 2163. [Google Scholar] [CrossRef]
- Para, I.; Albu, A.; Porojan, M.D. Adipokines and Arterial Stiffness in Obesity. Medicina 2021, 57, 653. [Google Scholar] [CrossRef]
- Flores-Cordero, J.A.; Pérez-Pérez, A.; Jiménez-Cortegana, C.; Alba, G.; Flores-Barragán, A.; Sánchez-Margalet, V. Obesity as a Risk Factor for Dementia and Alzheimer’s Disease: The Role of Leptin. Int. J. Mol. Sci. 2022, 23, 5202. [Google Scholar] [CrossRef]
- Jung, C.H.; Kim, B.Y.; Mok, J.O.; Kang, S.K.; Kim, C.H. Association between Serum Adipocytokine Levels and Microangiopathies in Patients with Type 2 Diabetes Mellitus. J. Diabetes Investig. 2014, 5, 333. [Google Scholar] [CrossRef] [Green Version]
- Yadav, A.; Jyoti, P.; Jain, S.K.; Bhattacharjee, J. Correlation of Adiponectin and Leptin with Insulin Resistance: A Pilot Study in Healthy North Indian Population. Indian J. Clin. Biochem. 2011, 26, 193. [Google Scholar] [CrossRef] [Green Version]
- Muskiet, F.A.J.; Carrera-Bastos, P.; Pruimboom, L.; Lucia, A.; Furman, D. Obesity and Leptin Resistance in the Regulation of the Type I Interferon Early Response and the Increased Risk for Severe COVID-19. Nutrients 2022, 14, 1388. [Google Scholar] [CrossRef]
- Mir, M.M.; Mir, R.; Alghamdi, M.A.A.; Wani, J.I.; Sabah, Z.U.; Jeelani, M.; Marakala, V.; Sohail, S.K.; O’haj, M.; Alharthi, M.H.; et al. Differential Association of Selected Adipocytokines, Adiponectin, Leptin, Resistin, Visfatin and Chemerin, with the Pathogenesis and Progression of Type 2 Diabetes Mellitus (T2DM) in the Asir Region of Saudi Arabia: A Case Control Study. J. Pers. Med. 2022, 12, 735. [Google Scholar] [CrossRef] [PubMed]
- Adiga, U.; Banawalikar, N.; Rai, T. Association of Leptin and Leptin Receptor Gene Polymorphisms with Insulin Resistance in Pregnant Women: A Cross-Sectional Study. F1000Research 2022, 11, 692. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Peng, D. Emerging Functions of Adipokines in Linking the Development of Obesity and Cardiovascular Diseases. Mol. Biol. Rep. 2020, 47, 7991–8006. [Google Scholar] [CrossRef] [PubMed]
- Markin, A.M.; Sobenin, I.A.; Grechko, A.V.; Zhang, D.; Orekhov, A.N. Cellular Mechanisms of Human Atherogenesis: Focus on Chronification of Inflammation and Mitochondrial Mutations. Front. Pharmacol. 2020, 11, 642. [Google Scholar] [CrossRef] [PubMed]
- Kirichenko, T.V.; Markina, Y.V.; Sukhorukov, V.N.; Khotina, V.A.; Wu, W.K.; Orekhov, A.N. A Novel Insight at Atherogenesis: The Role of Microbiome. Front. Cell Dev. Biol. 2020, 8, 586189. [Google Scholar] [CrossRef] [PubMed]
- Gerasimova, E.V.; Popkova, T.V.; Gerasimova, D.A.; Kirichenko, T.V. Macrophage Dysfunction in Autoimmune Rheumatic Diseases and Atherosclerosis. Int. J. Mol. Sci. 2022, 23, 4513. [Google Scholar] [CrossRef] [PubMed]
- Gruen, M.L.; Hao, M.; Piston, D.W.; Hasty, A.H. Leptin Requires Canonical Migratory Signaling Pathways for Induction of Monocyte and Macrophage Chemotaxis. Am. J. Physiol. Cell Physiol. 2007, 293, C1481–C1488. [Google Scholar] [CrossRef] [PubMed]
- Raman, P.; Khanal, S. Leptin in Atherosclerosis: Focus on Macrophages, Endothelial and Smooth Muscle Cells. Int. J. Mol. Sci. 2021, 22, 5446. [Google Scholar] [CrossRef]
- Cabrero, À.; Cubero, M.; Llaverías, G.; Alegret, M.; Sánchez, R.; Laguna, J.C.; Vázquez-Carrera, M. Leptin Down-Regulates Peroxisome Proliferator-Activated Receptor Gamma (PPAR-Gamma) MRNA Levels in Primary Human Monocyte-Derived Macrophages. Mol. Cell. Biochem. 2005, 275, 173–179. [Google Scholar] [CrossRef]
- Nedosugova, L.V.; Markina, Y.V.; Bochkareva, L.A.; Kuzina, I.A.; Petunina, N.A.; Yudina, I.Y.; Kirichenko, T.V. Inflammatory Mechanisms of Diabetes and Its Vascular Complications. Biomedicines 2022, 10, 1168. [Google Scholar] [CrossRef]
- Oda, A.; Taniguchi, T.; Yokoyama, M. Leptin Stimulates Rat Aortic Smooth Muscle Cell Proliferation and Migration. Kobe J. Med. Sci. 2001, 47, 141–150. [Google Scholar] [CrossRef]
- Parhami, F.; Tintut, Y.; Ballard, A.; Fogelman, A.M.; Demer, L.L. Leptin Enhances the Calcification of Vascular Cells. Circ. Res. 2001, 88, 954–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, S.; Le, Y. Role of Resistin in Inflammation and Inflammation-Related Diseases. Cell. Mol. Immunol. 2006, 3, 29–34. [Google Scholar] [PubMed]
- Steppan, C.M.; Lazar, M.A. The Current Biology of Resistin. J. Intern. Med. 2004, 255, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Yannakoulia, M.; Yiannakouris, N.; Blüher, S.; Matalas, A.L.; Klimis-Zacas, D.; Mantzoros, C.S. Body Fat Mass and Macronutrient Intake in Relation to Circulating Soluble Leptin Receptor, Free Leptin Index, Adiponectin, and Resistin Concentrations in Healthy Humans. J. Clin. Endocrinol. Metab. 2003, 88, 1730–1736. [Google Scholar] [CrossRef]
- Cebeci, E.; Cakan, C.; Gursu, M.; Uzun, S.; Karadag, S.; Koldas, M.; Calhan, T.; Helvaci, S.A.; Ozturk, S. The Main Determinants of Serum Resistin Level in Type 2 Diabetic Patients Are Renal Function and Inflammation Not Presence of Microvascular Complication, Obesity and Insulin Resistance. Exp. Clin. Endocrinol. Diabetes 2019, 127, 189–194. [Google Scholar] [CrossRef]
- Patel, L.; Buckels, A.C.; Kinghorn, I.J.; Murdock, P.R.; Holbrook, J.D.; Plumpton, C.; Macphee, C.H.; Smith, S.A. Resistin Is Expressed in Human Macrophages and Directly Regulated by PPARγ Activators. Biochem. Biophys. Res. Commun. 2003, 300, 472–476. [Google Scholar] [CrossRef]
- Jamaluddin, M.S.; Weakley, S.M.; Yao, Q.; Chen, C. Resistin: Functional Roles and Therapeutic Considerations for Cardiovascular Disease. Br. J. Pharmacol. 2012, 165, 622–632. [Google Scholar] [CrossRef] [Green Version]
- Nava-Salazar, S.; Flores-Pliego, A.; Pérez-Martínez, G.; Parra-Hernández, S.; Vanoye-Carlo, A.; Ibarguengoitia-Ochoa, F.; Perichart-Perera, O.; Reyes-Muñoz, E.; Solis-Paredes, J.M.; Espino y Sosa, S.; et al. Resistin Modulates Low-Density Lipoprotein Cholesterol Uptake in Human Placental Explants via PCSK9. Reprod. Sci. 2022, 29, 3242–3253. [Google Scholar] [CrossRef]
- Adeghate, E. An Update on the Biology and Physiology of Resistin. Cell. Mol. Life Sci. 2004, 61, 2485–2496. [Google Scholar] [CrossRef]
- Ren, Y.; Zhao, H.; Yin, C.; Lan, X.; Wu, L.; Du, X.; Griffiths, H.R.; Gao, D. Adipokines, Hepatokines and Myokines: Focus on Their Role and Molecular Mechanisms in Adipose Tissue Inflammation. Front. Endocrinol. 2022, 13, 873699. [Google Scholar] [CrossRef]
- Shin, J.H.; Park, S.H.; Cho, H.; Kim, J.H.; Choi, H. Adipokine Human Resistin Promotes Obesity-Associated Inflammatory Intervertebral Disc Degeneration via pro-Inflammatory Cytokine Cascade Activation. Sci. Rep. 2022, 12, 8936. [Google Scholar] [CrossRef] [PubMed]
- Kollari, E.; Zografou, I.; Sampanis, C.; Athyros, V.G.; Didangelos, T.; Mantzoros, C.S.; Karagiannis, A. Serum Adipokine Levels in Patients with Type 1 Diabetes Are Associated with Degree of Obesity but Only Resistin Is Independently Associated with Atherosclerosis Markers. Hormones 2022, 21, 91–101. [Google Scholar] [CrossRef]
- Su, K.Z.; Li, Y.R.; Zhang, D.; Yuan, J.H.; Zhang, C.S.; Liu, Y.; Song, L.M.; Lin, Q.; Li, M.W.; Dong, J. Relation of Circulating Resistin to Insulin Resistance in Type 2 Diabetes and Obesity: A Systematic Review and Meta-Analysis. Front. Physiol. 2019, 10, 1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saddi-Rosa, P.; Oliveira, C.S.; Giuffrida, F.M.; Reis, A.F. Visfatin, Glucose Metabolism and Vascular Disease: A Review of Evidence. Diabetol. Metab. Syndr. 2010, 2, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curat, C.A.; Wegner, V.; Sengenès, C.; Miranville, A.; Tonus, C.; Busse, R.; Bouloumié, A. Macrophages in Human Visceral Adipose Tissue: Increased Accumulation in Obesity and a Source of Resistin and Visfatin. Diabetologia 2006, 49, 744–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dakroub, A.; Nasser, S.A.; Kobeissy, F.; Yassine, H.M.; Orekhov, A.; Sharifi-Rad, J.; Iratni, R.; El-Yazbi, A.F.; Eid, A.H. Visfatin: An Emerging Adipocytokine Bridging the Gap in the Evolution of Cardiovascular Diseases. J. Cell. Physiol. 2021, 236, 6282–6296. [Google Scholar] [CrossRef] [PubMed]
- Brentano, F.; Schorr, O.; Ospelt, C.; Stanczyk, J.; Gay, R.E.; Gay, S.; Kyburz, D. Pre-B Cell Colony-Enhancing Factor/Visfatin, a New Marker of Inflammation in Rheumatoid Arthritis with Proinflammatory and Matrix-Degrading Activities. Arthritis Rheum. 2007, 56, 2829–2839. [Google Scholar] [CrossRef] [PubMed]
- Dahl, T.B.; Yndestad, A.; Skjelland, M.; Øie, E.; Dahl, A.; Michelsen, A.; Damås, J.K.; Tunheim, S.H.; Ueland, T.; Smith, C.; et al. Increased Expression of Visfatin in Macrophages of Human Unstable Carotid and Coronary Atherosclerosis: Possible Role in Inflammation and Plaque Destabilization. Circulation 2007, 115, 972–980. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.-J.; Wu, C.-S.; Lin, H.; Lee, I.-T.; Wu, C.-M.; Tseng, J.-J.; Chou, M.-M.; Sheu, W.H.-H. Visfatin-Induced Expression of Inflammatory Mediators in Human Endothelial Cells through the NF-ΚB Pathway. Int. J. Obes. 2009, 33, 465–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.C.; Song, J.; Lee, A.; Cho, D.; Kim, T.S. Visfatin Promotes Wound Healing through the Activation of ERK1/2 and JNK1/2 Pathway. Int. J. Mol. Sci. 2018, 19, 3642. [Google Scholar] [CrossRef]
- Aydoğan Baykara, R.; Küçük, A.; Tuzcu, A.; Tuzcu, G.; Cüre, E.; Uslu, A.U.; Omma, A. The Relationship of Serum Visfatin Levels with Clinical Parameters, Flow-Mediated Dilation, and Carotid Intima-Media Thickness in Patients with Ankylosing Spondylitis. Turk. J. Med. Sci. 2021, 51, 1865–1874. [Google Scholar] [CrossRef]
- Buechler, C.; Feder, S.; Haberl, E.M.; Aslanidis, C. Chemerin Isoforms and Activity in Obesity. Int. J. Mol. Sci. 2019, 20, 1128. [Google Scholar] [CrossRef] [Green Version]
- De Henau, O.; Degroot, G.N.; Imbault, V.; Robert, V.; de Poorter, C.; McHeik, S.; Galés, C.; Parmentier, M.; Springael, J.Y. Signaling Properties of Chemerin Receptors CMKLR1, GPR1 and CCRL2. PLoS ONE 2016, 11, e0164179. [Google Scholar] [CrossRef] [PubMed]
- Bozaoglu, K.; Bolton, K.; McMillan, J.; Zimmet, P.; Jowett, J.; Collier, G.; Walder, K.; Segal, D. Chemerin Is a Novel Adipokine Associated with Obesity and Metabolic Syndrome. Endocrinology 2007, 148, 4687–4694. [Google Scholar] [CrossRef] [PubMed]
- Kaur, J.; Adya, R.; Tan, B.K.; Chen, J.; Randeva, H.S. Identification of Chemerin Receptor (ChemR23) in Human Endothelial Cells: Chemerin-Induced Endothelial Angiogenesis. Biochem. Biophys. Res. Commun. 2010, 391, 1762–1768. [Google Scholar] [CrossRef] [Green Version]
- Dimitriadis, G.K.; Kaur, J.; Adya, R.; Miras, A.D.; Mattu, H.S.; Hattersley, J.G.; Kaltsas, G.; Tan, B.K.; Randeva, H.S. Chemerin Induces Endothelial Cell Inflammation: Activation of Nuclear Factor-Kappa Beta and Monocyte-Endothelial Adhesion. Oncotarget 2018, 9, 16678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sell, H.; Divoux, A.; Poitou, C.; Basdevant, A.; Bouillot, J.L.; Bedossa, P.; Tordjman, J.; Eckel, J.; Clément, K. Chemerin Correlates with Markers for Fatty Liver in Morbidly Obese Patients and Strongly Decreases after Weight Loss Induced by Bariatric Surgery. J. Clin. Endocrinol. Metab. 2010, 95, 2892–2896. [Google Scholar] [CrossRef] [Green Version]
- Mulvihill, E.E. Dipeptidyl Peptidase Inhibitor Therapy in Type 2 Diabetes: Control of the Incretin Axis and Regulation of Postprandial Glucose and Lipid Metabolism. Peptides 2018, 100, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Barchetta, I.; Cimini, F.A.; Dule, S.; Cavallo, M.G. Dipeptidyl Peptidase 4 (DPP4) as A Novel Adipokine: Role in Metabolism and Fat Homeostasis. Biomedicines 2022, 10, 2306. [Google Scholar] [CrossRef] [PubMed]
- Sell, H.; Blüher, M.; Klöting, N.; Schlich, R.; Willems, M.; Ruppe, F.; Knoefel, W.T.; Dietrich, A.; Fielding, B.A.; Arner, P.; et al. Adipose Dipeptidyl Peptidase-4 and Obesity: Correlation with Insulin Resistance and Depot-Specific Release from Adipose Tissue in Vivo and in Vitro. Diabetes Care 2013, 36, 4083–4090. [Google Scholar] [CrossRef] [PubMed]
- Sedighi, M.; Baluchnejadmojarad, T.; Fallah, S.; Moradi, N.; Afshin-Majd, S.; Roghani, M. The Association Between Circulating Klotho and Dipeptidyl Peptidase-4 Activity and Inflammatory Cytokines in Elderly Patients With Alzheimer Disease. Basic Clin. Neurosci. 2020, 11, 349. [Google Scholar] [CrossRef] [PubMed]
- Stengel, A.; Goebel-Stengel, M.; Teuffel, P.; Hofmann, T.; Buße, P.; Kobelt, P.; Rose, M.; Klapp, B.F. Obese Patients Have Higher Circulating Protein Levels of Dipeptidyl Peptidase IV. Peptides 2014, 61, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Röhrborn, D.; Brückner, J.; Sell, H.; Eckel, J. Reduced DPP4 Activity Improves Insulin Signaling in Primary Human Adipocytes. Biochem. Biophys. Res. Commun. 2016, 471, 348–354. [Google Scholar] [CrossRef]
- Brochu-Gaudreau, K.; Rehfeldt, C.; Blouin, R.; Bordignon, V.; Murphy, B.D.; Palin, M.F. Adiponectin Action from Head to Toe. Endocrine 2010, 37, 11–32. [Google Scholar] [CrossRef]
- Fruebis, J.; Tsao, T.-S.; Javorschi, S.; Ebbets-Reed, D.; Erickson, M.R.S.; Yen, F.T.; Bihain, B.E.; Lodish, H.F. Proteolytic Cleavage Product of 30-KDa Adipocyte Complement-Related Protein Increases Fatty Acid Oxidation in Muscle and Causes Weight Loss in Mice. Proc. Natl. Acad. Sci. USA 2001, 98, 2005. [Google Scholar] [CrossRef]
- Iwabu, M.; Okada-Iwabu, M.; Yamauchi, T.; Kadowaki, T. Adiponectin/Adiponectin Receptor in Disease and Aging. NPJ Aging Mech. Dis. 2015, 1, 15013. [Google Scholar] [CrossRef]
- Galic, S.; Oakhill, J.S.; Steinberg, G.R. Adipose Tissue as an Endocrine Organ. Mol. Cell. Endocrinol. 2010, 316, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Judd, R.L. Adiponectin Regulation and Function. Compr. Physiol. 2018, 8, 1031–1063. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, T.A.; Ouchi, N.; Shibata, R.; Walsh, K. Adiponectin Actions in the Cardiovascular System. Cardiovasc. Res. 2007, 74, 11. [Google Scholar] [CrossRef]
- Ouchi, N.; Kihara, S.; Arita, Y.; Okamoto, Y.; Maeda, K.; Kuriyama, H.; Hotta, K.; Nishida, M.; Takahashi, M.; Muraguchi, M.; et al. Adiponectin, an Adipocyte-Derived Plasma Protein, Inhibits Endothelial NF-KappaB Signaling through a CAMP-Dependent Pathway. Circulation 2000, 102, 1296–1301. [Google Scholar] [CrossRef]
- Ouchi, N.; Kihara, S.; Arita, Y.; Maeda, K.; Kuriyama, H.; Okamoto, Y.; Hotta, K.; Nishida, M.; Takahashi, M.; Nakamura, T.; et al. Novel Modulator for Endothelial Adhesion Molecules: Adipocyte-Derived Plasma Protein Adiponectin. Circulation 1999, 100, 2473–2476. [Google Scholar] [CrossRef] [Green Version]
- Lau, W.B.; Ohashi, K.; Wang, Y.; Ogawa, H.; Murohara, T.; Ma, X.L.; Ouchi, N. Role of Adipokines in Cardiovascular Disease. Circ. J. 2017, 81, 920–928. [Google Scholar] [CrossRef] [Green Version]
- Gradinaru, D.; Margina, D.; Borsa, C.; Ionescu, C.; Ilie, M.; Costache, M.; Dinischiotu, A.; Prada, G.I. Adiponectin: Possible Link between Metabolic Stress and Oxidative Stress in the Elderly. Aging Clin. Exp. Res. 2017, 29, 621–629. [Google Scholar] [CrossRef]
- Cnop, M.; Havel, P.J.; Utzschneider, K.M.; Carr, D.B.; Sinha, M.K.; Boyko, E.J.; Retzlaff, B.M.; Knopp, R.H.; Brunzell, J.D.; Kahn, S.E. Relationship of Adiponectin to Body Fat Distribution, Insulin Sensitivity and Plasma Lipoproteins: Evidence for Independent Roles of Age and Sex. Diabetologia 2003, 46, 459–469. [Google Scholar] [CrossRef] [Green Version]
- Turer, A.T.; Khera, A.; Ayers, C.R.; Turer, C.B.; Grundy, S.M.; Vega, G.L.; Scherer, P.E. Adipose Tissue Mass and Location Affect Circulating Adiponectin Levels. Diabetologia 2011, 54, 2515. [Google Scholar] [CrossRef] [Green Version]
- Gradidge, P.J.-L.; Jaff, N.G.; Norris, S.A.; Toman, M.; Crowther, N.J. The Negative Association of Lower Body Fat Mass with Cardiometabolic Disease Risk Factors Is Partially Mediated by Adiponectin. Endocr. Connect. 2022, 11, e220156. [Google Scholar] [CrossRef]
- Yan, H.; Yu, B.; Tian, J.; Xia, D.; Xu, Y.; Li, C. Serum Leptin and Adiponectin: Indicators of Cardiovascular Disease Secondary to Psoriasis. Indian J. Dermatol. 2022, 67, 109–114. [Google Scholar] [CrossRef]
- De Souza Batista, C.M.; Yang, R.Z.; Lee, M.J.; Glynn, N.M.; Yu, D.Z.; Pray, J.; Ndubuizu, K.; Patil, S.; Schwartz, A.; Kligman, M.; et al. Omentin Plasma Levels and Gene Expression Are Decreased in Obesity. Diabetes 2007, 56, 1655–1661. [Google Scholar] [CrossRef] [Green Version]
- Katsi, V.; Vamvakou, G.; Lekakis, J.; Tousoulis, D.; Stefanadis, C.; Makris, T.; Kallikazaros, I. Omentin, Fat and Heart: Classical Music with New Instruments. Heart Lung Circ. 2014, 23, 802–806. [Google Scholar] [CrossRef]
- Zhou, J.-Y.; Chan, L.; Zhou, S.-W. Omentin: Linking Metabolic Syndrome and Cardiovascular Disease. Curr. Vasc. Pharmacol. 2014, 12, 136–143. [Google Scholar] [CrossRef]
- Yin, J.; Hou, P.; Wu, Z.; Nie, Y. Decreased Levels of Serum Omentin-1 in Patients with Inflammatory Bowel Disease. Med. Sci. Monit. 2015, 21, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, Z.Y.; Liu, P.Y. Omentin-1: One Novel Biomarker for Calcified Aortic Valve Stenosis. Acta Cardiol. Sin. 2022, 38, 591. [Google Scholar] [CrossRef]
- Yan, X.; Wu, L.; Gao, M.; Yang, P.; Yang, J.; Deng, Y. Omentin Inhibits the Resistin-Induced Hypertrophy of H9c2 Cardiomyoblasts by Inhibiting the TLR4/MyD88/NF-ΚB Signaling Pathway. Exp. Ther. Med. 2022, 23, 292. [Google Scholar] [CrossRef]
- Lin, X.; Sun, Y.; Yang, S.; Yu, M.; Pan, L.; Yang, J.; Yang, J.; Shao, Q.; Liu, J.; Liu, Y.; et al. Omentin-1 Modulates Macrophage Function via Integrin Receptors Avβ3 and Avβ5 and Reverses Plaque Vulnerability in Animal Models of Atherosclerosis. Front. Cardiovasc. Med. 2021, 8, 757926. [Google Scholar] [CrossRef]
- Wang, J.; Gao, Y.; Lin, F.; Han, K.; Wang, X. Omentin-1 Attenuates Lipopolysaccharide (LPS)-Induced U937 Macrophages Activation by Inhibiting the TLR4/MyD88/NF-ΚB Signaling. Arch. Biochem. Biophys. 2020, 679, 108187. [Google Scholar] [CrossRef] [PubMed]
- Leandro, A.; Queiroz, M.; Azul, L.; Seiça, R.; Sena, C.M. Omentin: A Novel Therapeutic Approach for the Treatment of Endothelial Dysfunction in Type 2 Diabetes. Free Radic. Biol. Med. 2021, 162, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Cheng, Y.; Zhang, M.; Du, L.; Wu, X.; Lu, C.; Li, X.; Liu, S.; Zhao, A.; Zhang, S.; et al. Neuroprotective Effects of Omentin-1 Against Cerebral Hypoxia/Reoxygenation Injury via Activating GAS6/Axl Signaling Pathway in Neuroblastoma Cells. Front. Cell Dev. Biol. 2022, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Shibata, R.; Takahashi, R.; Kataoka, Y.; Ohashi, K.; Ikeda, N.; Kihara, S.; Murohara, T.; Ouchi, N. Association of a Fat-Derived Plasma Protein Omentin with Carotid Artery Intima-Media Thickness in Apparently Healthy Men. Hypertens. Res. 2011, 34, 1309–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, M.; Morioka, T.; Hayashi, M.; Kakutani, Y.; Yamazaki, Y.; Kurajoh, M.; Mori, K.; Fukumoto, S.; Shioi, A.; Shoji, T.; et al. Plasma Omentin Levels Are Inversely Associated with Atherosclerosis in Type 2 Diabetes Patients with Increased Plasma Adiponectin Levels: A Cross-Sectional Study. Cardiovasc. Diabetol. 2019, 18, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Gao, Y. Clinical Relevance of Serum Omentin-1 Levels as a Biomarker of Prognosis in Patients with Acute Cerebral Infarction. Brain Behav. 2020, 10, e01678. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Zhao, M.; Voilquin, L.; Jung, Y.; Aikio, M.A.; Sahai, T.; Dou, F.Y.; Roche, A.M.; Carcamo-Orive, I.; Knowles, J.W.; et al. Isthmin-1 Is an Adipokine That Promotes Glucose Uptake and Improves Tolerance and Hepatic Steatosis. Cell Metab. 2021, 33, 1836. [Google Scholar] [CrossRef]
- Hu, M.; Zhang, X.; Hu, C.; Teng, T.; Tang, Q.Z. A Brief Overview about the Adipokine: Isthmin-1. Front. Cardiovasc. Med. 2022, 9, 939757. [Google Scholar] [CrossRef]
- Heeren, J.; Scheja, L. Isthmin 1—A Novel Insulin-like Adipokine. Nat. Rev. Endocrinol. 2021, 17, 709–710. [Google Scholar] [CrossRef]
- Nguyen, N.; Xu, S.; Lam, T.Y.W.; Liao, W.; Wong, W.S.F.; Ge, R. ISM1 Suppresses LPS-Induced Acute Lung Injury and Post-Injury Lung Fibrosis in Mice. Mol. Med. 2022, 28, 72. [Google Scholar] [CrossRef]
- Xiang, W.; Ke, Z.; Zhang, Y.; Ho-Yuet Cheng, G.; Irwan, I.D.; Sulochana, K.N.; Potturi, P.; Wang, Z.; Yang, H.; Wang, J.; et al. Isthmin Is a Novel Secreted Angiogenesis Inhibitor That Inhibits Tumour Growth in Mice. J. Cell. Mol. Med. 2011, 15, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayada, C.; Toru, Ü; Korkut, Y. Nesfatin-1 and Its Effects on Different Systems. Hippokratia 2015, 19, 4. [Google Scholar] [PubMed]
- Stengel, A.; Mori, M.; Taché, Y. The Role of Nesfatin-1 in the Regulation of Food Intake and Body Weight–Recent Developments and Future Endeavors. Obes. Rev. 2013, 14, 859–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruszała, M.; Pilszyk, A.; Niebrzydowska, M.; Kimber-trojnar, Ż.; Trojnar, M.; Leszczyńska-gorzelak, B. Novel Biomolecules in the Pathogenesis of Gestational Diabetes Mellitus 2.0. Int. J. Mol. Sci. 2022, 23, 4364. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, K.; Ren, Y. Nesfatin-1 Protects H9c2 Cardiomyocytes against Cobalt Chloride-Induced Hypoxic Injury by Modulating the MAPK and Notch1 Signaling Pathways. J. Biol. Res. 2021, 28, 21. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Dong, J.; Mu, Y.; Liu, X. Nesfatin-1 Protects against Diabetic Cardiomyopathy in the Streptozotocin-Induced Diabetic Mouse Model via the P38-MAPK Pathway. Bioengineered 2022, 13, 14670. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Zhu, Y.; Chen, L.; Wang, Y. Nesfatin-1 Alleviated Lipopolysaccharide-Induced Acute Lung Injury through Regulating Inflammatory Response Associated with Macrophages Modulation. J. Cardiothorac. Surg. 2022, 17, 206. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.J.; Wen, F.J.; Qiu, D.; Wang, S.Z. Nesfatin-1 in Lipid Metabolism and Lipid-Related Diseases. Clin. Chim. Acta 2021, 522, 23–30. [Google Scholar] [CrossRef]
- Sjögren, P.; Sierra-Johnson, J.; Kallings, L.V.; Cederholm, T.; Kolak, M.; Halldin, M.; Brismar, K.; de Faire, U.; Hellénius, M.L.; Fisher, R.M. Functional Changes in Adipose Tissue in a Randomised Controlled Trial of Physical Activity. Lipids Health Dis. 2012, 11, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashad, N.M.; Sayed, S.E.; Sherif, M.H.; Sitohy, M.Z. Effect of a 24-Week Weight Management Program on Serum Leptin Level in Correlation to Anthropometric Measures in Obese Female: A Randomized Controlled Clinical Trial. Diabetes Metab. Syndr. 2019, 13, 2230–2235. [Google Scholar] [CrossRef] [PubMed]
- Siegrist, M.; Heitkamp, M.; Braun, I.; Vogg, N.; Haller, B.; Langhof, H.; Koenig, W.; Halle, M. Changes of Omentin-1 and Chemerin during 4 Weeks of Lifestyle Intervention and 1 Year Follow-up in Children with Obesity. Clin. Nutr. 2021, 40, 5648–5654. [Google Scholar] [CrossRef]
- Seven, E.; Husemoen, L.L.N.; Sehested, T.S.G.; Ibsen, H.; Wachtell, K.; Linneberg, A.; Jeppesen, J.L. Adipocytokines, C-Reactive Protein, and Cardiovascular Disease: A Population-Based Prospective Study. PLoS ONE 2015, 10, e0128987. [Google Scholar] [CrossRef] [Green Version]
- Castela, I.; Morais, J.; Barreiros-Mota, I.; Silvestre, M.P.; Marques, C.; Rodrigues, C.; Ismael, S.; Araújo, J.R.; Ângelo-Dias, M.; Martins, C.; et al. Decreased Adiponectin/Leptin Ratio Relates to Insulin Resistance in Adults with Obesity. Am. J. Physiol. Endocrinol. Metab. 2022. [Google Scholar] [CrossRef] [PubMed]
- Effect of Obesity on Resistin Concentrations in Normal, Pre-Obese and Obese Apparently Healthy Nigerian-Africans. Available online: https://pubmed.ncbi.nlm.nih.gov/35924881/ (accessed on 24 November 2022).
- Gruzdeva, O.V.; Dyleva, Y.A.; Belik, E.V.; Sinitsky, M.Y.; Stasev, A.N.; Kokov, A.N.; Brel, N.K.; Krivkina, E.O.; Bychkova, E.E.; Tarasov, R.S.; et al. Relationship between Epicardial and Coronary Adipose Tissue and the Expression of Adiponectin, Leptin, and Interleukin 6 in Patients with Coronary Artery Disease. J. Pers. Med. 2022, 12, 129. [Google Scholar] [CrossRef] [PubMed]
- Varma, B.; Ogunmoroti, O.; Ndumele, C.E.; Zhao, D.; Szklo, M.; Sweeney, T.; Allison, M.A.; Budoff, M.J.; Subramanya, V.; Bertoni, A.G.; et al. Higher Leptin Levels Are Associated with Coronary Artery Calcium Progression: The Multi-Ethnic Study of Atherosclerosis (MESA). Diabetes Epidemiol. Manag. 2022, 6, 100047. [Google Scholar] [CrossRef] [PubMed]
- Puchałowicz, K.; Kłoda, K.; Dziedziejko, V.; Rać, M.; Wojtarowicz, A.; Chlubek, D.; Safranow, K. Association of Adiponectin, Leptin and Resistin Plasma Concentrations with Echocardiographic Parameters in Patients with Coronary Artery Disease. Diagnostics 2021, 11, 1774. [Google Scholar] [CrossRef] [PubMed]
- Özkan, E.A.; Sadigov, A.; Öztürk, O. Evaluation of Serum Omentin-1, Vaspin, Leptin, Adiponectin Levels in Obese/Overweight Children and Their Relationship with Non-Alcoholic Fatty Liver Disease. Clin. Nutr. Res. 2022, 11, 194. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Zusi, C.; Csermely, A.; Salvagno, G.L.; Colecchia, A.; Lippi, G.; Maffeis, C.; Targher, G. Association between Lower Plasma Adiponectin Levels and Higher Liver Stiffness in Type 2 Diabetic Individuals with Nonalcoholic Fatty Liver Disease: An Observational Cross-Sectional Study. Hormones 2022, 21, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, M.; Sakuma, Y.; Ogino, J.; Iwai, R.; Watanabe, S.; Inoue, T.; Takahashi, H.; Suzuki, Y.; Kinoshita, D.; Takemura, K.; et al. Sex Differences in Predictive Factors for Onset of Type 2 Diabetes in Japanese Individuals: A 15-Year Follow-up Study. J. Diabetes Investig. 2022. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, H. Association of Serum Leptin and Insulin Levels among Type 2 Diabetes Mellitus Patients: A Case-Control Study. Medicine 2022, 101, e31006. [Google Scholar] [CrossRef] [PubMed]
- Musialik, K.; Miller-Kasprzak, E.; Walczak, M.; Markuszewski, L.; Bogdański, P. The Association between Serum Resistin Level, Resistin (-420C/G) Single Nucleotide Variant, and Markers of Endothelial Dysfunction, Including Salt Taste Preference in Hypertensive Patients. Nutrients 2022, 14, 1789. [Google Scholar] [CrossRef]
- Wu, O.; Hang Leng, J.; Zhang, X.Y.; Liu, W.; Zhang, H.; Yang, F.F.; Li, J.J.; Zhang, G.Z.; Lu, X. The Value of Adiponectin-Resistin (AR) Index in Newly Diagnosed Obesity Hypertension: A Case Control Study among Chinese Adult. Clin. Exp. Hypertens. 2022, 44, 40–45. [Google Scholar] [CrossRef]
- Nan, J.; Zhang, X.; Li, D.; Jin, M.; Guo, G.; Dong, J.; Li, X. Effect of Leptin Levels of Type 2 Diabetes Mellitus and Diabetic Complications. Clin. Investig. Med. 2022, 45, 32–46. [Google Scholar] [CrossRef]
- Venojärvi, M.; Lindström, J.; Aunola, S.; Nuutila, P.; Atalay, M. Improved Aerobic Capacity and Adipokine Profile Together with Weight Loss Improve Glycemic Control without Changes in Skeletal Muscle GLUT-4 Gene Expression in Middle-Aged Subjects with Impaired Glucose Toerance. Int. J. Environ. Res. Public Health 2022, 19, 8327. [Google Scholar] [CrossRef]
- Golubović, M.V.; Dimić, D.; Antić, S.; Radenković, S.; Djindjić, B.; Jovanović, M. Relationship of Adipokine to Insulin Sensitivity and Glycemic Regulation in Obese Women--the Effect of Body Weight Reduction by Caloric Restriction. Vojnosanit. Pregl. 2013, 70, 284–291. [Google Scholar] [CrossRef]
- Askarpour, M.; Alizadeh, S.; Hadi, A.; Symonds, M.E.; Miraghajani, M.; Sheikhi, A.; Ghaedi, E. Effect of Bariatric Surgery on the Circulating Level of Adiponectin, Chemerin, Plasminogen Activator Inhibitor-1, Leptin, Resistin, and Visfatin: A Systematic Review and Meta-Analysis. Horm. Metab. Res. 2020, 52, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Dludla, P.V.; Nkambule, B.B.; Mazibuko-Mbeje, S.E.; Nyambuya, T.M.; Mxinwa, V.; Mokgalaboni, K.; Ziqubu, K.; Cirilli, I.; Marcheggiani, F.; Louw, J.; et al. Adipokines as a Therapeutic Target by Metformin to Improve Metabolic Function: A Systematic Review of Randomized Controlled Trials. Pharmacol. Res. 2021, 163, 105219. [Google Scholar] [CrossRef] [PubMed]
- Firouzjaei, A.; Li, G.-C.; Wang, N.; Liu, W.-X.; Zhu, B.-M. Comparative Evaluation of the Therapeutic Effect of Metformin Monotherapy with Metformin and Acupuncture Combined Therapy on Weight Loss and Insulin Sensitivity in Diabetic Patients. Nutr. Diabetes 2016, 6, e209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Xia, N. Influence of Sodium-Glucose Cotransporter-2 Inhibitors on Plasma Adiponectin in Patients with Type 2 Diabetes: A Meta-Analysis of Randomized Controlled Trials. Horm. Metab. Res. 2022. [Google Scholar] [CrossRef]
- Hanson, P.; Randeva, H.; Cuthbertson, D.J.; O’Hare, P.J.; Parsons, N.; Chatha, K.; Reidy, G.; Weickert, M.O.; Barber, T.M. The DAPA-DIET Study: Metabolic Response to Dapagliflozin Combined with Dietary Carbohydrate Restriction in Patients with Type 2 Diabetes Mellitus and Obesity—A Longitudinal Cohort Study. Endocrinol. Diabetes Metab. 2022, 5, e381. [Google Scholar] [CrossRef] [PubMed]
- Schiapaccassa, A.; Maranhão, P.A.; de Souza, M.D.G.C.; Panazzolo, D.G.; Nogueira Neto, J.F.; Bouskela, E.; Kraemer-Aguiar, L.G. 30-Days Effects of Vildagliptin on Vascular Function, Plasma Viscosity, Inflammation, Oxidative Stress, and Intestinal Peptides on Drug-Naïve Women with Diabetes and Obesity: A Randomized Head-to-Head Metformin-Controlled Study. Diabetol. Metab. Syndr. 2019, 11, 70. [Google Scholar] [CrossRef] [Green Version]
- Francisco, V.; Sanz, M.J.; Real, J.T.; Marques, P.; Capuozzo, M.; Ait Eldjoudi, D.; Gualillo, O. Adipokines in Non-Alcoholic Fatty Liver Disease: Are We on the Road toward New Biomarkers and Therapeutic Targets? Biology 2022, 11, 1237. [Google Scholar] [CrossRef]
- Watanabe, T.; Watanabe-Kominato, K.; Takahashi, Y.; Kojima, M.; Watanabe, R. Adipose Tissue-Derived Omentin-1 Function and Regulation. In Comprehensive Physiology; Wiley: Hoboken, NJ, USA, 2017; pp. 765–781. [Google Scholar]
- Bahreini, M.; Ramezani, A.-H.; Shishehbor, F.; Mansoori, A. The Effect of Omega-3 on Circulating Adiponectin in Adults with Type 2 Diabetes Mellitus: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Can. J. Diabetes 2018, 42, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Sohn, M.; Na, G.Y.; Chu, J.; Joung, H.; Kim, B.-K.; Lim, S. Efficacy and Safety of Lactobacillus Plantarum K50 on Lipids in Koreans With Obesity: A Randomized, Double-Blind Controlled Clinical Trial. Front. Endocrinol. 2022, 12, 790046. [Google Scholar] [CrossRef]
- Naseri, K.; Saadati, S.; Ghaemi, F.; Ashtary-Larky, D.; Asbaghi, O.; Sadeghi, A.; Afrisham, R.; de Courten, B. The Effects of Probiotic and Synbiotic Supplementation on Inflammation, Oxidative Stress, and Circulating Adiponectin and Leptin Concentration in Subjects with Prediabetes and Type 2 Diabetes Mellitus: A GRADE-Assessed Systematic Review, Meta-Analysis, and Meta-Regression of Randomized Clinical Trials. Eur. J. Nutr. 2022. [Google Scholar] [CrossRef]
- Chruściel, P.; Sahebkar, A.; Rembek-Wieliczko, M.; Serban, M.-C.; Ursoniu, S.; Mikhailidis, D.P.; Jones, S.R.; Mosteoru, S.; Blaha, M.J.; Martin, S.S.; et al. Impact of Statin Therapy on Plasma Adiponectin Concentrations: A Systematic Review and Meta-Analysis of 43 Randomized Controlled Trial Arms. Atherosclerosis 2016, 253, 194–208. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, W.; Zhao, M.; Jia, G.; Sun, R. Effect of Atorvastatin Treatment on Circulating Adiponectin: A Meta-Analysis of Randomized Controlled Trials. Lipids Health Dis. 2019, 18, 228. [Google Scholar] [CrossRef] [Green Version]
- Kadoglou, N.P.E.; Velidakis, N.; Khattab, E.; Kassimis, G.; Patsourakos, N. The Interplay between Statins and Adipokines. Is This Another Explanation of Statins’ ‘Pleiotropic’ Effects? Cytokine 2021, 148, 155698. [Google Scholar] [CrossRef] [PubMed]
- Bochar, O.M.; Sklyarova, H.Y.; Faynyk, A.F.; Bochar, V.T.; Kuzminov, Y.B.; Bazylevych, A.Y. The Effect of Therapy with Olmesartan or Telmisartan in Patients with Arterial Hypertension Combined with Obesity. Wiad Lek. 2020, 73, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Nedogoda, S.V.; Ledyaeva, A.A.; Chumachok, E.V.; Tsoma, V.V.; Mazina, G.; Salasyuk, A.S.; Barykina, I.N. Randomized Trial of Perindopril, Enalapril, Losartan and Telmisartan in Overweight or Obese Patients with Hypertension. Clin. Drug Investig. 2013, 33, 553–561. [Google Scholar] [CrossRef] [PubMed]

| Obesity-Associated Diseases | Study Design | Results |
|---|---|---|
| Obesity | Study of the adiponectin/leptin ratio (AdipoQ/Lep) as a biomarker of adipose tissue and metabolic function in obese individuals without diabetes. | Lower AdipoQ/Lep correlated with higher BMI, body fat mass, waist-to-height ratio, and plasma resistin and may, therefore, be an early marker for the development of insulin resistance in obese adults [128]. |
| Study of resistin levels in obese Nigerian Africans. | The resistin concentration was higher in obese individuals [129]. | |
| Study in control vs. obese women at baseline and after 12 weeks of following the Mediterranean diet. | Serum leptin levels were higher in obese women and weight loss was associated with a decrease in serum leptin [125]. | |
| Coronary artery disease | Study of patients with coronary artery disease with isolation of adipocytes from subcutaneous, perivascular, and epicardial adipose tissue. | Decreased expression and secretion of the adiponectin gene in epicardial adipose tissue, and high level of leptin gene expression [130]. |
| Study of adipokine levels in individuals with coronary artery calcium progression. | Higher leptin levels were associated with coronary artery calcium progression; no association was observed for resistin and adiponectin [131]. | |
| Study of anthropometric and echocardiographic parameters and adipokine levels in Caucasian patients. | Adiponectin and resistin, but not leptin, were associated with echocardiographic parameters of cardiac remodeling and dysfunction [132]. | |
| NAFLD | Study of adipokine levels in obese/overweight children and their association with the degree of hepatosteatosis. | Levels of leptin, omentin-1, and adiponectin were higher in the obese group and increased with a higher degree of hepatosteatosis [133]. |
| Study of adipokines in patients without NAFLD, with steatosis and with NAFLD. | Lower plasma levels of adiponectin were associated with the presence and severity of NAFLD [134]. | |
| Diabetes | Study of the factors associated with the onset of type 2 diabetes, including serum adiponectin. | In women, unlike men, a decrease in the level of adiponectin was the only significant risk factor for the development of type 2 diabetes [135]. |
| Study of the level of leptin as a biomarker for the development of insulin resistance in patients with type 2 diabetes. | Higher serum leptin levels were found in patients with type 2 diabetes and metabolic syndrome [136]. | |
| Study of the relationship of adipokines in patients with type 2 diabetes in Saudi Arabia. | The decrease in adiponectin levels and the increase in leptin, visfatin, and chemerin were more significant as BMI increased in patients with type 2 diabetes; no association of resistin with type 2 diabetes was found [41]. | |
| Arterial hypertension | Study of resistin levels in patients with and without arterial hypertension. | The level of resistin did not differ significantly between patients with and without arterial hypertension [137]. |
| Study of the adiponectin–resistin index as a marker of hypertension in obese patients. | The adiponectin–resistin index was strongly associated with an increased risk of obesity-related hypertension [138]. | |
| Study of the relationship between leptin levels and arterial hypertension as a complication of type 2 diabetes mellitus. | Leptin levels in patients with arterial hypertension and in patients with type 2 diabetes were higher than in the group without hypertension [139]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirichenko, T.V.; Markina, Y.V.; Bogatyreva, A.I.; Tolstik, T.V.; Varaeva, Y.R.; Starodubova, A.V. The Role of Adipokines in Inflammatory Mechanisms of Obesity. Int. J. Mol. Sci. 2022, 23, 14982. https://doi.org/10.3390/ijms232314982
Kirichenko TV, Markina YV, Bogatyreva AI, Tolstik TV, Varaeva YR, Starodubova AV. The Role of Adipokines in Inflammatory Mechanisms of Obesity. International Journal of Molecular Sciences. 2022; 23(23):14982. https://doi.org/10.3390/ijms232314982
Chicago/Turabian StyleKirichenko, Tatiana V., Yuliya V. Markina, Anastasia I. Bogatyreva, Taisiya V. Tolstik, Yurgita R. Varaeva, and Antonina V. Starodubova. 2022. "The Role of Adipokines in Inflammatory Mechanisms of Obesity" International Journal of Molecular Sciences 23, no. 23: 14982. https://doi.org/10.3390/ijms232314982