
Integrative Proteomics and Transcriptomics Profiles of the Oviduct Reveal the Prolificacy-Related Candidate Biomarkers of Goats (Capra hircus) in Estrous Periods

 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Overview of Quantitative Proteomics Analysis of Goat Oviducts
2.2. Oviduct Protein Co-Expression Network Analysis Identifies Module–Trait Relationships and Points to Core Mediators of Fecundity
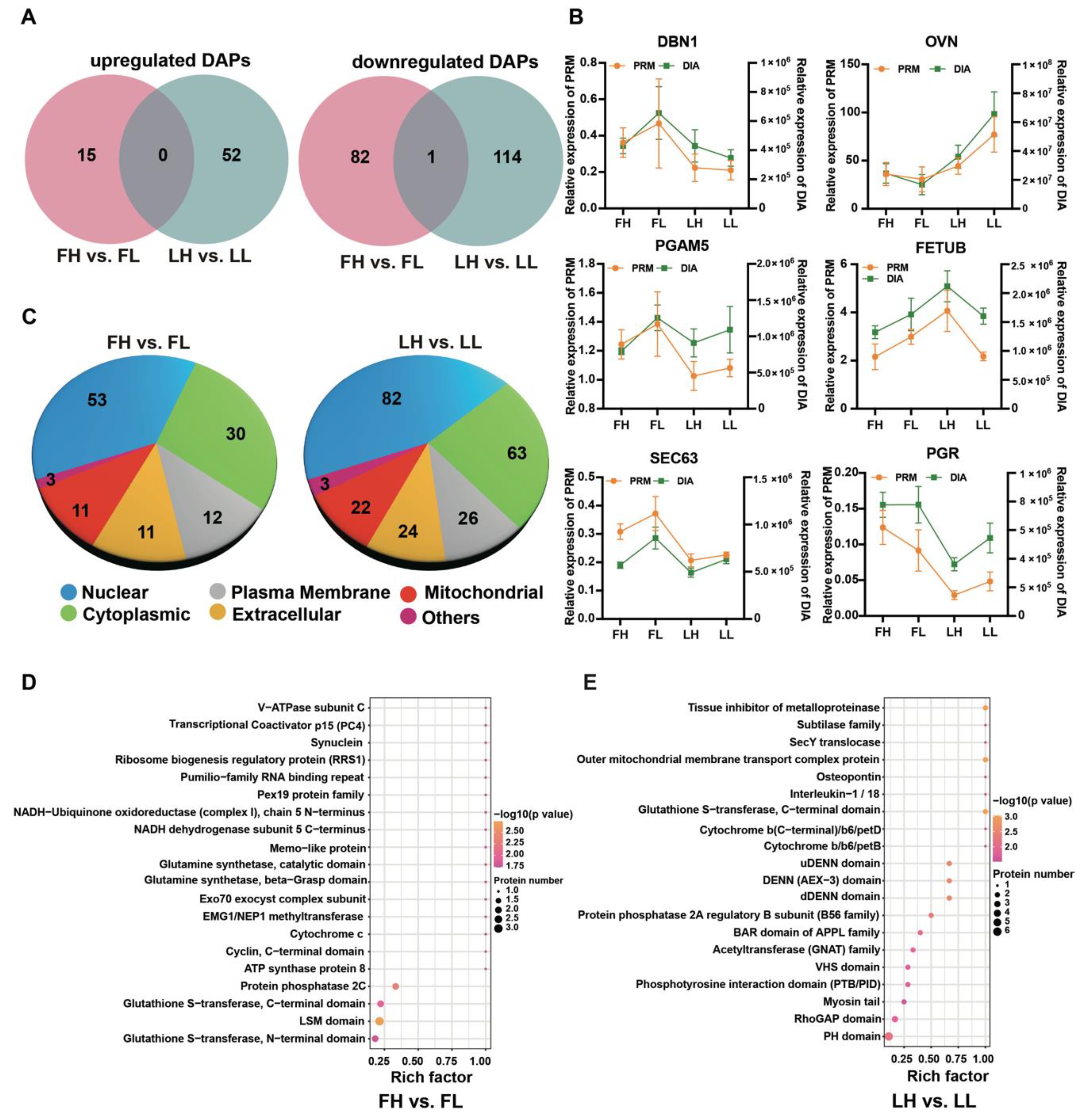
2.3. Identification of Differentially Abundant Proteins
2.4. Parallel Reaction Monitoring (PRM) Analysis
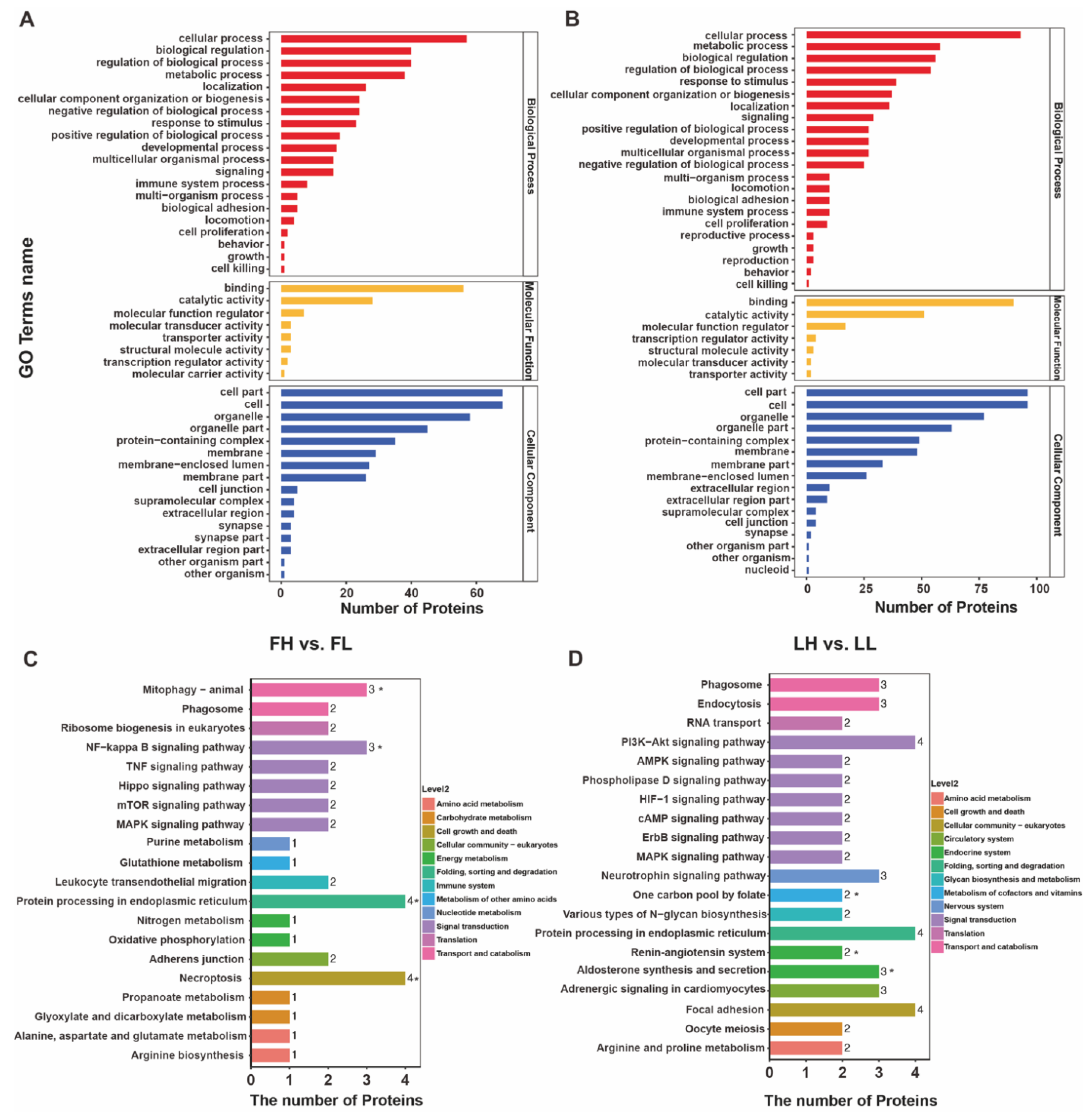
2.5. Bioinformatic Analysis of DAPs
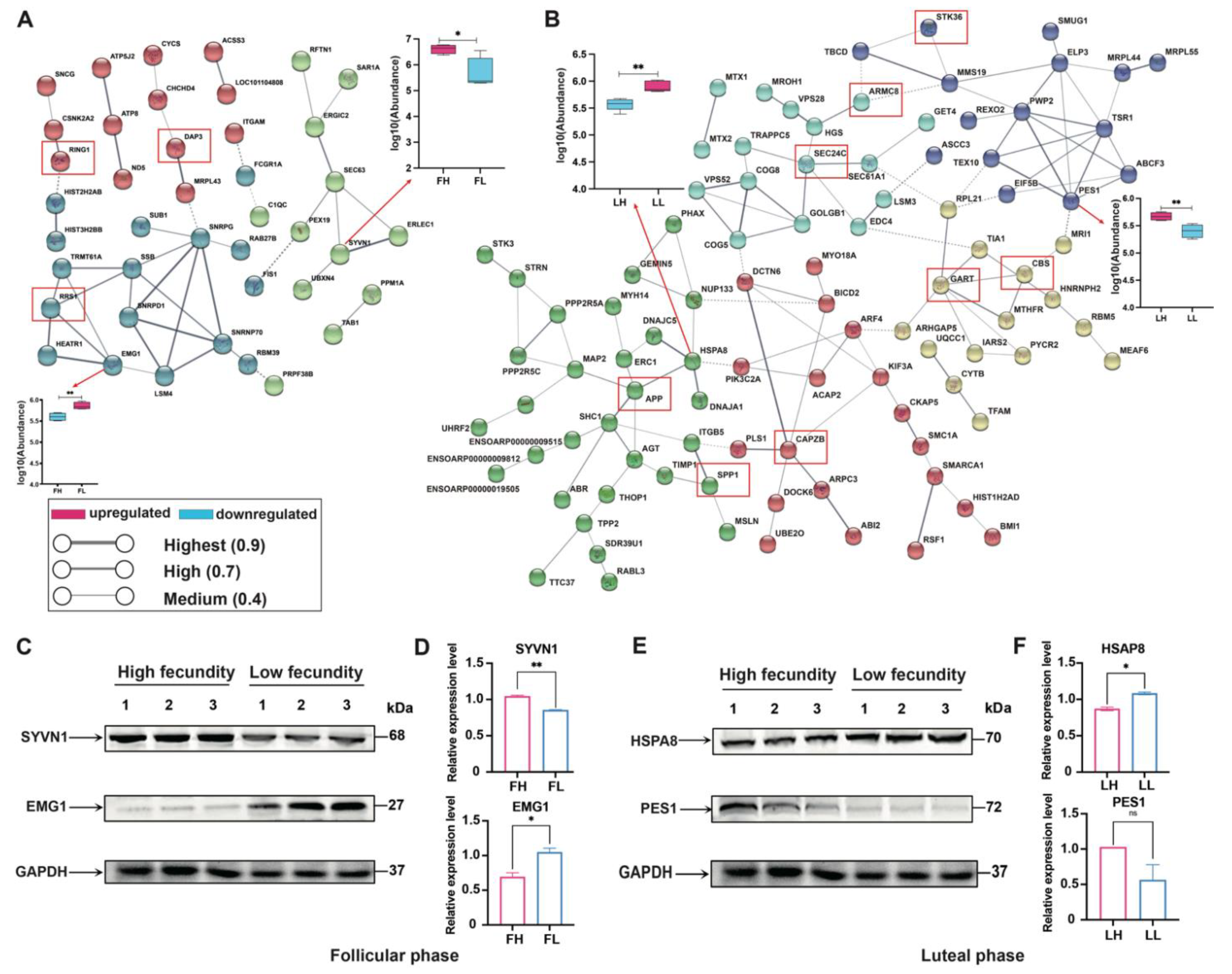
2.6. Protein–Protein Interaction (PPI) Network Analysis for Goat Prolificacy Trait
2.7. Western Blot Analysis of Hub Proteins
2.8. Combined Analysis of Proteome and Transcriptome (DAPs/DEGs) Responding to Prolificacy Trait of Yunshang Black Goat in Oviducts
2.9. RT-qPCR Validation of Candidate DEGs
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Animals and Sample Acquisition
4.3. Protein Preparation and Fractionation for Data-Dependent Acquisition (DDA) Library Generation
4.4. Data-Dependent Acquisition (DDA) Mass Spectrometry Assay
4.5. Data-Independent Acquisition (DIA) Mass Spectrometry Assay
4.6. Raw Mass Spectrometry Data Analysis
4.7. Weighted Gene Co-expression Network Analysis
4.8. Data Analysis and Identification of Differentially Abundant Proteins
4.9. Functional Annotation of Differentially Abundant Proteins
4.10. Transcriptome Sequencing
4.11. Correlation Analyses of Transcriptome and Proteome Profiles
4.12. Validation of Proteomic Data with Parallel Reaction Monitoring (PRM)
4.13. Western Blot Analysis
4.14. Measuring mRNA Levels Using Real-Time Quantitative PCR
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Shah, A.M.; Cai, Y.M.; Zou, H.W.; Zhang, X.F.; Wang, L.Z.; Xue, B.; Yu, P.Q.; Wang, Z.S.; Peng, Q.H. Effects of supplementation of branches and leaves trimmed from tea plant on growth performance, rumen fermentation and meat composition of Nanjiang yellow goats. Animals 2019, 9, 590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridi, A.; Perecin, F.; da Silveira, J.C. Extracellular vesicles mediated early embryo–maternal interactions. Int. J. Mol. Sci. 2022, 21, 1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fazeli, A. Maternal communication with gametes and embryos. Theriogenology 2008, 70, 1182–1187. [Google Scholar] [CrossRef]
- Li, S.; Winuthayanon, W. Oviduct: Roles in fertilization and early embryo development. J. Endocrinol. 2017, 232, R1–R26. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.W.; Liu, X.Y.; Tang, B.; Li, C.; Kou, Z.H.; Li, L.; Liu, W.Q.; Wu, Y.; Kou, X.C.; Li, J.Y.; et al. Protein expression landscape of mouse embryos during pre-implantation development. Cell. Rep. 2017, 21, 3957–3969. [Google Scholar] [CrossRef] [Green Version]
- Stitzel, M.L.; Seydoux, G. Regulation of the oocyte-to-zygote transition. Science. 2007, 316, 407–408. [Google Scholar] [CrossRef]
- Besenfelder, U.; Havlicek, V.; Brem, G. Role of the oviduct in early embryo development. Reprod. Domest. Anim. 2012, 47, 156–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenngott, R.A.; Sinowatz, F. Prenatal development of the bovine oviduct. Anat. Histol. Embryol. 2007, 36, 272–283. [Google Scholar] [CrossRef]
- Goudet, G. Fertilisation in the horse and paracrine signalling in the oviduct. Reprod. Fertil. Dev. 2011, 23, 941–951. [Google Scholar] [CrossRef]
- Lee, S.H.; Song, E.J.; Hwangbo, Y.; Lee, S.; Park, C.K. Change of uterine histroph proteins during follicular and luteal phase in pigs. Anim. Reprod. Sci. 2016, 168, 26–33. [Google Scholar] [CrossRef]
- Coy, P.; García-Vázquez, F.A.; Visconti, P.E.; Avilés, M. Roles of the oviduct in mammalian fertilization. Reproduction 2012, 144, 649–660. [Google Scholar] [CrossRef] [Green Version]
- Mondejar, I.; Acuna, O.S.; Izquierdo-Rico, M.J.; Coy, P.; Aviles, M. The oviduct: Functional genomic and proteomic approach. Reprod. Domest. Anim. 2012, 47, 22–29. [Google Scholar] [CrossRef]
- Seytanoglu, A.; Georgiou, A.S.; Sostaric, E.; Watson, P.F.; Holt, W.V.; Fazeli, A. Oviductal cell proteome alterations during the reproductive cycle in pigs. J. Proteome Res. 2008, 7, 2825–2833. [Google Scholar] [CrossRef]
- Hu, J.J.; Xiao, L.F.; Song, L.L.; Ge, W.B.; Duan, H.W.; Jiang, Y. The expression of melatonin receptors MT1 and MT2 is regulated by E2 in sheep oviduct. Gen. Comp. Endocrinol. 2020, 286, 113135. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.S.; Ge, W.B.; Ding, Z.Q.; Zeng, J.L.; Wang, W.J.; Duan, H.W.; Hu, J.J.; Zhang, Y.; Zhao, X.X. Expression of dihydrotestosterone synthases and androgen receptor in sheep oviduct ampulla and its regulation by estradiol and progesterone. Reprod. Biol. 2022, 22, 100573. [Google Scholar] [CrossRef] [PubMed]
- Lapointe, J.; Kimmins, S.; Maclaren, L.A.; Bilodeau, J.F. Estrogen selectively up-regulates the phospholipid hydroperoxide glutathione peroxidase in the oviducts. Endocrinology 2005, 146, 2583–2592. [Google Scholar] [CrossRef] [Green Version]
- Binelli, M.; Gonella-Diaza, A.M.; Mesquita, F.S.; Membrive, C.M.B. Sex steroid-mediated control of oviductal function in Cattle. Biology 2018, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- McNutt, T.L.; Olds-Clarke, P.; Way, A.L.; Suarez, S.S.; Killian, G.J. Effect of follicular or oviductal fluids on movement characteristics of bovine sperm during capacitation in vitro. J. Androl. 1994, 15, 328–336. [Google Scholar] [PubMed]
- Satake, N.; Elliott, R.M.; Watson, P.F.; Holt, W.V. Sperm selection and competition in pigs may be mediated by the differential motility activation and suppression of sperm subpopulations within the oviduct. J. Exp. Biol. 2006, 209, 1560–1572. [Google Scholar] [CrossRef] [Green Version]
- Pradeep, M.A.; Jagadeesh, J.; De, A.K.; Kaushik, J.K.; Malakar, D.; Kumar, S.; Dang, A.K.; Das, S.K.; Mohanty, A.K. Purification, sequence characterization and effect of goat oviduct-specific glycoprotein on in vitro embryo development. Theriogenology 2011, 75, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Abdulghani, M.; Song, G.Y.; Kaur, H.; Walley, J.W.; Tuteja, G. Comparative analysis of the transcriptome and proteome during mouse placental development. J. Proteome Res. 2019, 18, 2088–2099. [Google Scholar] [CrossRef]
- Schwanhausser, B.; Busse, D.; Li, N.; Dittmar, G.; Schuchhardt, J.; Wolf, J.; Chen, W.; Selbach, M. Global quantification of mammalian gene expression control. Nature 2011, 473, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Walley, J.W.; Sartor, R.C.; Shen, Z.X.; Schmitz, R.J.; Wu, K.J.; Urich, M.A.; Nery, J.R.; Smith, L.G.; Schnable, J.C.; Ecker, J.R.; et al. Integration of Omic Networks in a Developmental Atlas of Maize. Science 2016, 353, 814–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La, Y.F.; Tang, J.S.; Guo, X.F.; Zhang, L.P.; Gan, S.Q.; Zhang, X.S.; Zhang, J.L.; Hu, W.P.; Chu, M.X. Proteomic analysis of sheep uterus reveals its role in prolificacy. J. Proteom. 2020, 210, 103526. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Zhou, M.; Dorsey, T.H.; Prieto, D.A.; Wang, X.W.; Ruppin, E.; Veenstra, T.D.; Ambs, S. Integrated proteotranscriptomics of breast cancer reveals globally increased protein-mRNA concordance associated with subtypes and survival. Genome. Med. 2018, 10, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.L.; Xia, M.L.; Su, X.J.; Yuan, P.; Li, X.K.; Zhou, C.Y.; Wan, Z.P.; Zou, W. Photolytic degradation elevated the toxicity of polylactic acid microplastics to developing zebrafish by triggering mitochondrial dysfunction and apoptosis. J. Hazard. Mater. 2021, 413, 125321. [Google Scholar] [CrossRef]
- Liao, Q.; Tang, J.Q.; Wang, H.Y.; Yang, W.C.; He, L.X.; Wang, Y.Y.; Yang, Z.H. Dynamic proteome responses to sequential reduction of Cr(VI) and adsorption of Pb(II) by Pannonibacter phragmitetus BB. J. Hazard. Mater. 2020, 386, 121988. [Google Scholar] [CrossRef]
- Sun, Z.P.; Hong, Q.H.; Liu, Y.F.; He, X.Y.; Di, R.; Wang, X.Y.; Ren, C.H.; Zhang, Z.J.; Chu, M.X. Characterization of circular RNA profiles of oviduct reveal the potential mechanism in prolificacy trait of goat in the estrus cycle. Front. Physiol. 2022, 13, 990691. [Google Scholar] [CrossRef]
- Fang, L.; Hu, X.R.; Cui, L.; Lv, P.P.; Ma, X.Q.; Ye, Y.H. Serum and follicular fluid fetuin-B levels are correlated with fertilization rates in conventional IVF cycles. J. Assist. Reprod. Genet. 2019, 36, 1101–1107. [Google Scholar] [CrossRef] [PubMed]
- Bylander, A.; Lind, K.; Goksor, M.; Billig, H.; Larsson, D. The classical progesterone receptor mediates the rapid reduction of fallopian tube ciliary beat frequency by progesterone. Reprod. Biol. Endocrinol. 2013, 11, 33. [Google Scholar] [CrossRef] [PubMed]
- Hadek, R. Alteration of pH in the sheep’s oviduct. Nature 1953, 171, 976. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.D.; Qi, X.Y.; Chen, S.S.; Feng, J.; Chen, H.J.; Qin, Z.Y.; Deng, Y.M. An integrated transcriptomic and proteomic approach to dynamically study the mechanism of pollen-pistil interactions during jasmine crossing. J. Proteom. 2021, 249, 104380. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.P.; Liu, N.X.; Geng, Z.; Ji, M.J.; Wang, S.M.; Zhuang, Y.M.; Wang, D.; He, G.; Zhao, S.T.; Zhou, G.K.; et al. Integrated transcriptome and proteome analysis reveals brassinosteroid-mediated regulation of cambium initiation and patterning in woody stem. Hortic. Res. 2022, 9, uhab048. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.Y.; Li, G.C.; Wang, Z.Q.; Guo, Y.R.; Liu, N.Y. Combined transcriptomic, proteomic and genomic analysis identifies reproductive-related proteins and potential modulators of female behaviors in Spodoptera litura. Genomics 2021, 113, 1876–1894. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.V.; Frey, B.L.; Scalf, M.; Cesnik, A.J.; Rolfs, Z.; Pike, K.A.; Yang, B.; Keller, M.P.; Jarrard, D.F.; Shortreed, M.R.; et al. MetaNetwork enhances biological insights from quantitative proteomics differences by combining clustering and enrichment analyses. J. Proteome Res. 2022, 21, 410–419. [Google Scholar] [CrossRef]
- Liu, X.C.; Han, M.X.; Xu, Y.; Wang, H.Y.; Li, B. Knockdown of the premature ovarian insufficiency candidate gene NUP107 in ovarian granulosa cells affects cell functions, including receptor expression and estrogen synthesis. Reprod. Dev. Med. 2019, 3, 133–140. [Google Scholar] [CrossRef]
- Smith, B.N.; Topp, S.D.; Fallini, C.; Shibata, H.; Chen, H.J.; Troakes, C.; King, A.; Ticozzi, N.; Kenna, K.P.; Soragia-Gkazi, A.; et al. Mutations in the vesicular trafficking protein Annexin A11 are associated with amyotrophic lateral sclerosis. Sci. Transl. Med. 2017, 9, eaad9157. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.C.; Fernandopulle, M.S.; Wang, G.Z.; Choi, H.; Hao, L.; Drerup, C.M.; Patel, R.; Qamar, S.; Nixon-Abell, J.; Shen, Y.; et al. RNA granules hitchhike on lysosomes for long-distance transport, using Annexin A11 as a molecular tether. Cell 2019, 179, 147–164.e20. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.S.; Guo, C.M.; Liu, S.Q.; Qi, H.B.; Yin, Y.; Liang, R.; Sun, M.Z.; Greenaway, F.T. Annexin A11 in disease. Clin. Chim. Acta 2014, 431, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Bridi, A.; Perecin, F.; da Silveira, J.C. The role of the oviduct and extracellular vesicles during early embryo development in bovine. Anim. Reprod. 2022, 19, e20220015. [Google Scholar] [CrossRef]
- Lyndin, M.; Kravtsova, O.; Sikora, K.; Lyndina, Y.; Kuzenko, Y.; Awuah, W.A.; Abdul-Rahman, T.; Hyriavenko, N.; Sikora, V.; Romaniuk, A. COX2 Effects on endometrial carcinomas progression. Pathol. Res. Pract. 2022, 238, 154082. [Google Scholar] [CrossRef] [PubMed]
- Jarzabek, K.; Koda, M.; Walentowicz-Sadlecka, M.; Grabiec, M.; Laudanski, P.; Wolczynski, S. Altered expression of ERs, aromatase, and COX2 connected to estrogen action in type 1 endometrial cancer biology. Tumour. Biol. 2013, 34, 4007–4016. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.G.; Stewart, C.L. Loss of cyclooxygenase-2 retards decidual growth but does not inhibit embryo implantation or development to term. Biol. Reprod. 2003, 68, 401–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Yang, X.Y.; Wang, T.; Yang, L.; He, Y.Y.; Miskimins, K.; Qian, S.Y. Knockdown delta-5-desaturase in breast cancer cells that overexpress COX-2 results in inhibition of growth, migration and invasion via a dihomo-γ-linolenic acid peroxidation dependent mechanism. BMC Cancer 2018, 18, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosseini, F.; Mahdian-Shakib, A.; Jadidi-Niaragh, F.; Enderami, S.E.; Mohammadi, H.; Hemmatzadeh, M.; Mohammed, H.A.; Anissian, A.; Kokhaei, P.; Mirshafiey, A.; et al. Anti-inflammatory and anti-tumor effects of α-l-guluronic acid (G2013) on cancer-related inflammation in a murine breast cancer model. Biomed. Pharm. 2018, 98, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.W.; Yuan, R.R.; Bai, Y.Y.; Yang, Y.T.; Song, X.Y.; Lan, X.Y.; Pan, C.Y. A deletion mutation within the goat AKAP13 gene is significantly associated with litter size. Anim. Biotechnol. 2021, 1–7. [Google Scholar] [CrossRef]
- Jiang, R.W.; Tang, X.F.; Pan, J.L.; Li, G.Z.; Yang, N.J.; Tang, Y.D.; Bi, S.L.; Cai, H.; Chen, Q.H.; Chen, D.J.; et al. CDC42 governs normal oviduct multiciliogenesis through activating AKT to ensure timely embryo transport. Cell. Death Dis. 2022, 13, 757. [Google Scholar] [CrossRef]
- Ward, S.M.; Hwang, S.J.; Yan, W.; Offermanns, S.; Sanders, K.M. Intrinsic pacemaker activity and propulsive forces provided by the myosalpinx are necessary for egg and embryo transport in the oviduct. Biol. Reprod. 2022, 106, 4–5. [Google Scholar] [CrossRef] [PubMed]
- Nita-Lazar, M.; Noonan, V.; Rebustini, I.; Walker, J.; Menko, A.S.; Kukuruzinska, M.A. Overexpression of DPAGT1 leads to aberrant N-glycosylation of E-cadherin and cellular discohesion in oral cancer. Cancer. Res. 2009, 69, 5673–5680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budna-Tukan, J.; Swiatly-Blaszkiewicz, A.; Celichowski, P.; Kaluzna, S.; Konwerska, A.; Sujka-Kordowska, P.; Jankowski, M.; Kulus, M.; Jeseta, M.; Piotrowska-Kempisty, H.; et al. “Biological Adhesion” is a significantly regulated molecular process during long-term primary in vitro culture of oviductal epithelial cells (Oecs): A transcriptomic and proteomic study. Int. J. Mol. Sci. 2019, 20, 3387. [Google Scholar] [CrossRef]
- Cal, S.; Freije, J.M.; Lopez, J.M.; Takada, Y.; Lopez-Otín, C. ADAM 23/MDC3, a human disintegrin that promotes cell adhesion via interaction with the alphavbeta3 integrin through an RGD-independent mechanism. Mol. Biol. Cell. 2000, 11, 1457–1469. [Google Scholar] [CrossRef]
- Kempisty, B.; Ziolkowska, A.; Ciesiolka, S.; Piotrowska, H.; Antosik, P.; Bukowska, D.; Nowicki, M.; Brussow, K.P.; Zabel, M. Study on connexin gene and protein expression and cellular distribution in relation to real-time proliferation of porcine granulosa cells. J. Biol. Regul. Homeost. Agents 2014, 28, 625–635. [Google Scholar] [PubMed]
- Mahe, C.; Zlotkowska, A.M.; Reynaud, K.; Tsikis, G.; Mermillod, P.; Druart, X.; Schoen, J.; Saint-Dizier, M. Sperm migration, selection, survival, and fertilizing ability in the mammalian oviduct. Biol. Reprod. 2021, 105, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Tokuhiro, K.; Ikawa, M.; Benham, A.M.; Okabe, M. Protein disulfide isomerase homolog PDILT is required for quality control of sperm membrane protein ADAM3 and male fertility [corrected]. Proc. Natl. Acad. Sci. USA 2012, 109, 3850–3855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Ubeda, R.; Garcia-Vazquez, F.A.; Romar, R.; Gadea, J.; Munoz, M.; Hunter, R.H.; Coy, P. Oviductal transcriptome is modified after insemination during spontaneous ovulation in the sow. PLoS ONE 2015, 10, e0130128. [Google Scholar] [CrossRef] [Green Version]
- Usami, F.M.; Arata, M.; Shi, D.; Oka, S.; Higuchi, Y.; Tissir, F.; Takeichi, M.; Fujimori, T. Intercellular and intracellular cilia orientation is coordinated by CELSR1 and CAMSAP3 in oviduct multi-ciliated cells. J. Cell Sci. 2021, 134, jcs257006. [Google Scholar] [CrossRef] [PubMed]
- Awamleh, Z.; Han, V.K.M. Identification of miR-210-5p in human placentae from pregnancies complicated by preeclampsia and intrauterine growth restriction, and its potential role in the pregnancy complications. Pregnancy. Hypertens. 2020, 19, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.Y.; Yang, F.S. Circular RNA circRNA-0039459 promotes the migration, invasion, and proliferation of liver cancer cells through the adsorption of miR-432. Bioengineered 2022, 13, 11810–11821. [Google Scholar] [CrossRef]
- Wu, X.L.; Sandhu, S.; Patel, N.; Triggs-Raine, B.; Ding, H. EMG1 is essential for mouse pre-implantation embryo development. BMC Dev. Biol. 2010, 10, 99. [Google Scholar] [CrossRef] [Green Version]
- Tatone, C.; Di Emidio, G. Mitochondria biology in reproductive function. Antioxidants 2022, 11, 1978. [Google Scholar] [CrossRef]
- Maezawa, S.; Hasegawa, K.; Yukawa, M.; Sakashita, A.; Alavattam, K.G.; Andreassen, P.R.; Vidal, M.; Koseki, H.; Barski, A.; Namekawa, S.H. Polycomb directs timely activation of germline genes in spermatogenesis. Genes Dev. 2017, 31, 1693–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokobayashi, S.; Liang, C.Y.; Kohler, H.; Nestorov, P.; Liu, Z.; Vidal, M.; van Lohuizen, M.; Roloff, T.C.; Peters, A.H. PRC1 coordinates timing of sexual differentiation of female primordial germ cells. Nature 2013, 495, 236–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.M.; Park, J.E.; Yoo, I.; Han, J.; Kim, N.; Lim, W.J.; Cho, E.S.; Choi, B.; Choi, S.; Kim, T.H.; et al. Integrated transcriptomes throughout swine oestrous cycle reveal dynamic changes in reproductive tissues interacting networks. Sci. Rep. 2018, 8, 5436. [Google Scholar] [CrossRef] [Green Version]
- Sheibani, L.; Lechuga, T.J.; Zhang, H.; Hameed, A.; Wing, D.A.; Kumar, S.; Rosenfeld, C.R.; Chen, D.B. Augmented H2S production via cystathionine-beta-synthase upregulation plays a role in pregnancy-associated uterine vasodilation. Biol. Reprod. 2017, 96, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Qi, Q.R.; Lechuga, T.J.; Patel, B.; Nguyen, N.A.; Yang, Y.H.; Li, Y.; Sarnthiyakul, S.; Zhang, Q.W.; Bai, J.; Makhoul, J.; et al. Enhanced stromal cell CBS-H2S production promotes estrogen-stimulated human endometrial angiogenesis. Endocrinology 2020, 161, bqaa176. [Google Scholar] [CrossRef] [PubMed]
- Fujii, D.T.; Yohannes, E.; Por, E.D.; Gillette, L.; Beesley, R.D.; Heitmann, R.J.; Chow, G.E.; Burney, R.O. The proteome of human Fallopian tube lavages during the phase of embryo transit reveals candidate proteins for the optimization of preimplantation embryo culture. Hum. Reprod. 2021, 36, 367–380. [Google Scholar] [CrossRef]
- Cheng, L.; Yuan, B.; Ying, S.Y.; Niu, C.; Mai, H.; Guan, X.X.; Yang, X.H.; Teng, Y.; Lin, J.; Huang, J.J.; et al. PES1 is a critical component of telomerase assembly and regulates cellular senescence. Sci. Adv. 2019, 5, eaav1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, H.; Son, D.; Jang, Y.J.; Hong, K. Indispensable role for mouse ELP3 in embryonic stem cell maintenance and early development. Biochem. Biophys. Res. Commun. 2016, 478, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Adams, E.J.; Khoriaty, R.; Kiseleva, A.; Cleuren, A.C.A.; Tomberg, K.; van der Ent, M.A.; Gergics, P.; Tang, V.T.; Zhu, G.J.; Hoenerhoff, M.J.; et al. Murine SEC24D can substitute functionally for SEC24C during embryonic development. Sci. Rep. 2021, 11, 21100. [Google Scholar] [CrossRef] [PubMed]
- Kramer, A.C.; Erikson, D.W.; McLendon, B.A.; Seo, H.; Hayashi, K.; Spencer, T.E.; Bazer, F.W.; Burghardt, R.C.; Johnson, G.A. SPP1 expression in the mouse uterus and placenta: Implications for implantation. Biol. Reprod. 2021, 105, 892–904. [Google Scholar] [CrossRef]
- Elliott, R.M.; Lloyd, R.E.; Fazeli, A.; Sostaric, E.; Georgiou, A.S.; Satake, N.; Watson, P.F.; Holt, W.V. Effects of HSPA8, an evolutionarily conserved oviductal protein, on boar and bull spermatozoa. Reproduction 2009, 137, 191–203. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.Y.; Leushkin, E.; Liechti, A.; Ovchinnikova, S.; Mossinger, K.; Bruning, T.; Rummel, C.; Grutzner, F.; Cardoso-Moreira, M.; Janich, P.; et al. Transcriptome and translatome co-evolution in mammals. Nature 2020, 588, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.H.; Wang, M.; Li, S.; Jian, R.Q.; Xiao Li, X.; Chan, J.; Dong, G.L.; Fang, H.Y.; Robinson, A.E.; Consortium, G.; et al. A quantitative proteome map of the human body. Cell 2020, 183, 269.e19–283.e19. [Google Scholar] [CrossRef]
- Zhao, L.M.; Li, F.D.; Zhang, X.X.; Zhang, D.Y.; Li, X.L.; Zhang, Y.K.; Zhao, Y.; Song, Q.Z.; Huang, K.; Xu, D.; et al. Integrative analysis of transcriptomics and proteomics of longissimus thoracis of the Hu sheep compared with the Dorper sheep. Meat. Sci. 2022, 193, 108930. [Google Scholar] [CrossRef]
- Sun, Z.P.; Zhang, Z.J.; Liu, Y.F.; Ren, C.H.; He, X.Y.; Jiang, Y.T.; Ouyang, Y.N.; Hong, Q.H.; Chu, M.X. Integrated analysis of mRNAs and long non-coding RNAs expression of oviduct that provides novel insights into the prolificacy mechanism of goat (Capra. hircus). Genes 2022, 13, 1031. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.P.; Hong, Q.H.; Liu, Y.F.; Ren, C.H.; He, X.Y.; Jiang, Y.T.; Ouyang, Y.N.; Chu, M.X.; Zhang, Z.J. Oviduct transcriptomic reveals the regulation of mRNAs and lncRNAs related to goat prolificacy in the luteal phase. Animals 2022, 12, 2823. [Google Scholar] [CrossRef]
- Garg, R.; Shankar, R.; Thakkar, B.; Kudapa, H.; Krishnamurthy, L.; Mantri, N.; Varshney, R.K.; Bhatia, S.; Jain, M. Transcriptome analyses reveal genotype-and developmental stage-specific molecular responses to drought and salinity stresses in chickpea. Sci. Rep. 2016, 6, 19228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.Y.; Lei, G.; Zhou, H.W.; He, C.; Liao, J.L.; Huang, Y.J. Quantitative iTRAQ-based proteomic analysis of rice grains to assess high night temperature stress. Proteomics 2017, 17, 1600365. [Google Scholar] [CrossRef] [Green Version]
- Luyao Zhang, L.Y.; Liu, K.X.; Qingrui Zhuan, Q.R.; Liu, Z.Q.; Lin Meng, L.; Fu, X.W.; Jia, G.X.; Hou, Y.P. Mitochondrial calcium disorder affects early embryonic development in mice through regulating the ERK/MAPK pathway. Oxid. Med. Cell. Longev. 2022, 2022, 8221361. [Google Scholar] [CrossRef]
- Li, S.; O’Neill, S.R.S.; Zhang, Y.; Holtzman, M.J.; Takemaru, K.I.; Korach, K.S.; Winuthayanon, W. Estrogen receptor α is required for oviductal transport of embryos. FASEB J. 2017, 31, 1595–1607. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Rosenberger, G.; Koh, C.C.; Guo, T.; Rost, H.L.; Kouvonen, P.; Collins, B.C.; Heusel, M.; Liu, Y.S.; Caron, E.; Vichalkovski, A.; et al. A repository of assays to quantify 10,000 human proteins by SWATH-MS. Sci. Data 2014, 1, 140031. [Google Scholar] [CrossRef] [Green Version]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.L.; Zhou, H.Z.; Wu, Y.R.; Zhang, H.; Lin, J.; Jiang, X.Y.; He, Q.J.; Zhu, J.S.; Li, Y.; Yu, H.; et al. OsSPL3, an SBP-Domain protein, regulates crown root development in rice. Plant Cell. 2019, 31, 1257–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovaka, S.; Zimin, A.V.; Pertea, G.M.; Razaghi, R.; Salzberg, S.L.; Pertea, M. Transcriptome assembly from long-read RNA-seq alignments with StringTie2. Genome Biol. 2019, 20, 278. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacLean, B.; Tomazela, D.M.; Shulman, N.; Chambers, M.; Finney, G.L.; Frewen, B.; Kern, R.; Tabb, D.L.; Liebler, D.C.; MacCoss, M.J. Skyline: An open source document editor for creating and analyzing targeted proteomics experiments. Bioinformatics 2010, 26, 966–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence (5′-3′) | Product Size (bp) |
|---|---|---|
| JAK1 | F: CGGAGAGTACACAGCGGAGGAG | 124 |
| R: GCGATTCGGAGCATACCAGAGC | ||
| HOOK2 | F: GCCGACAAGCACTGATGGATCTC | 115 |
| R: GGCAGAGACTGTGACTTCAAGGTTC | ||
| SRSF5 | F: GCTAAGTGCGTCAGTTGTGGAGAG | 90 |
| R: GAGAGGTCCGTAGTCTGCGATAGAG | ||
| MATR3 | F: GCAGTCTACAAACCCAGCACCAG | 107 |
| R: TTCCATTTCCAGCACCCAGATTTCC | ||
| RPL21 | F: GAGCCGAGATAGCTTCCTGAAACG | 145 |
| R: GGGTTCCTTTCCATTGGTCCTCAC | ||
| GPT2 | F: GCCAGCAGCCAATCACCTTCC | 131 |
| R: GTTCCCACCGCAAGCCTGTAG | ||
| SPOCK1 | F: AGTGACGGAGGATGACGAGGATG | 132 |
| R: AGACAAATGCAGGAACGGGAAGTG | ||
| APOD | F: AGAAGGGAAATGTGGCAGCAGAAG | 98 |
| R: TCAGACACGCAAGGTAACAGAATGG | ||
| RPL19 | F: ATCGCCAATGCCAACTC | 154 |
| R: CCTTTCGCTTACCTATACC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Z.; Liu, Y.; He, X.; Di, R.; Wang, X.; Ren, C.; Zhang, Z.; Chu, M. Integrative Proteomics and Transcriptomics Profiles of the Oviduct Reveal the Prolificacy-Related Candidate Biomarkers of Goats (Capra hircus) in Estrous Periods. Int. J. Mol. Sci. 2022, 23, 14888. https://doi.org/10.3390/ijms232314888
Sun Z, Liu Y, He X, Di R, Wang X, Ren C, Zhang Z, Chu M. Integrative Proteomics and Transcriptomics Profiles of the Oviduct Reveal the Prolificacy-Related Candidate Biomarkers of Goats (Capra hircus) in Estrous Periods. International Journal of Molecular Sciences. 2022; 23(23):14888. https://doi.org/10.3390/ijms232314888
Chicago/Turabian StyleSun, Zhipeng, Yufang Liu, Xiaoyun He, Ran Di, Xiangyu Wang, Chunhuan Ren, Zijun Zhang, and Mingxing Chu. 2022. "Integrative Proteomics and Transcriptomics Profiles of the Oviduct Reveal the Prolificacy-Related Candidate Biomarkers of Goats (Capra hircus) in Estrous Periods" International Journal of Molecular Sciences 23, no. 23: 14888. https://doi.org/10.3390/ijms232314888