

γδ T Lymphocytes as a Double-Edged Sword—State of the Art in Gynecological Diseases
 , , and
, , and
Abstract
:1. Introduction—Subpopulations of Gamma Delta (γδ) T Lymphocytes
2. Variations in the Phenotype and Function of γδ T Lymphocytes in Human Cancers
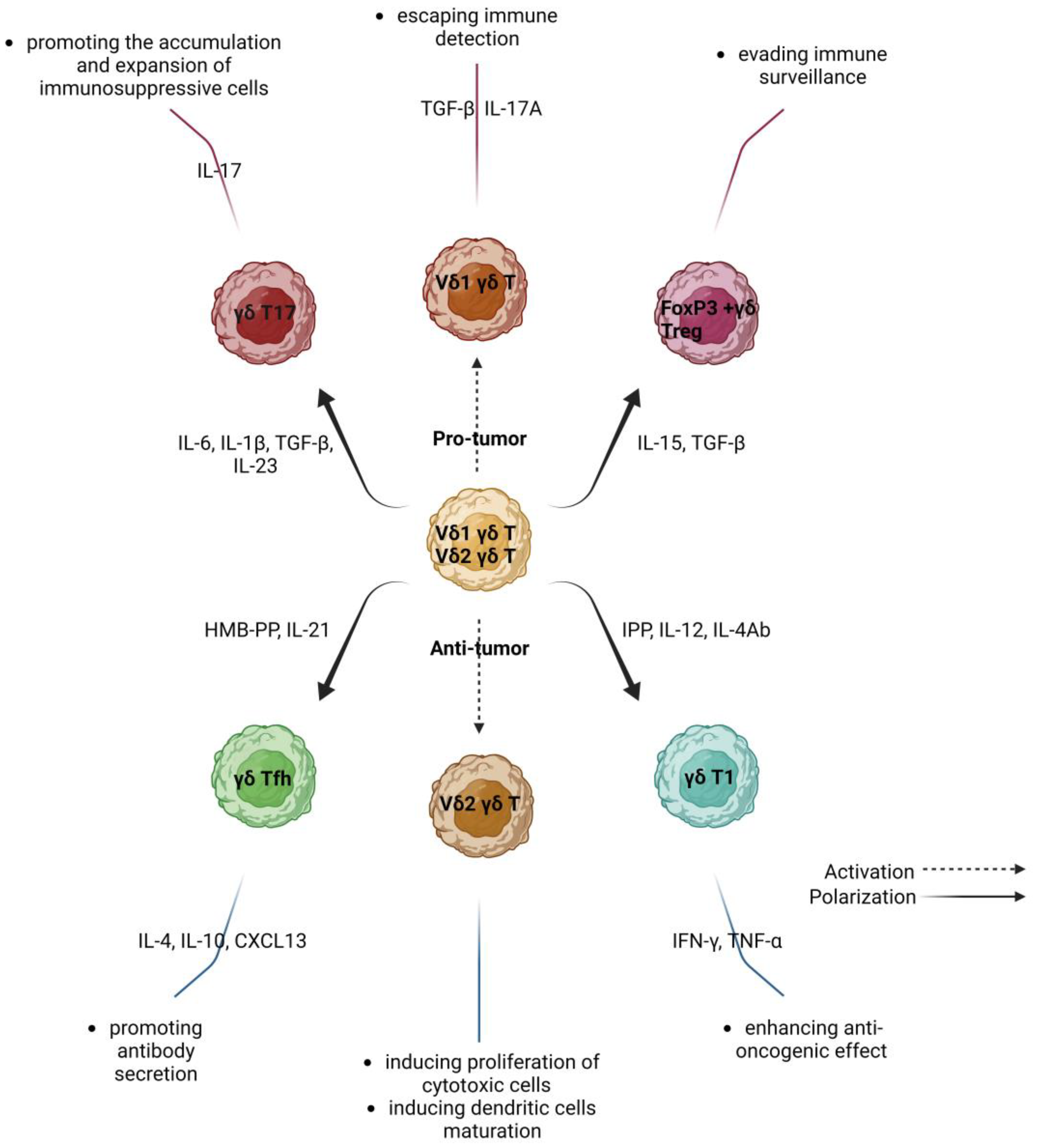
3. γδ T Lymphocytes in Cancer—Friends or Foes?
3.1. Protumor Activity of γδ T Lymphocytes
3.2. Anti-Tumor Activity of γδ T Lymphocytes
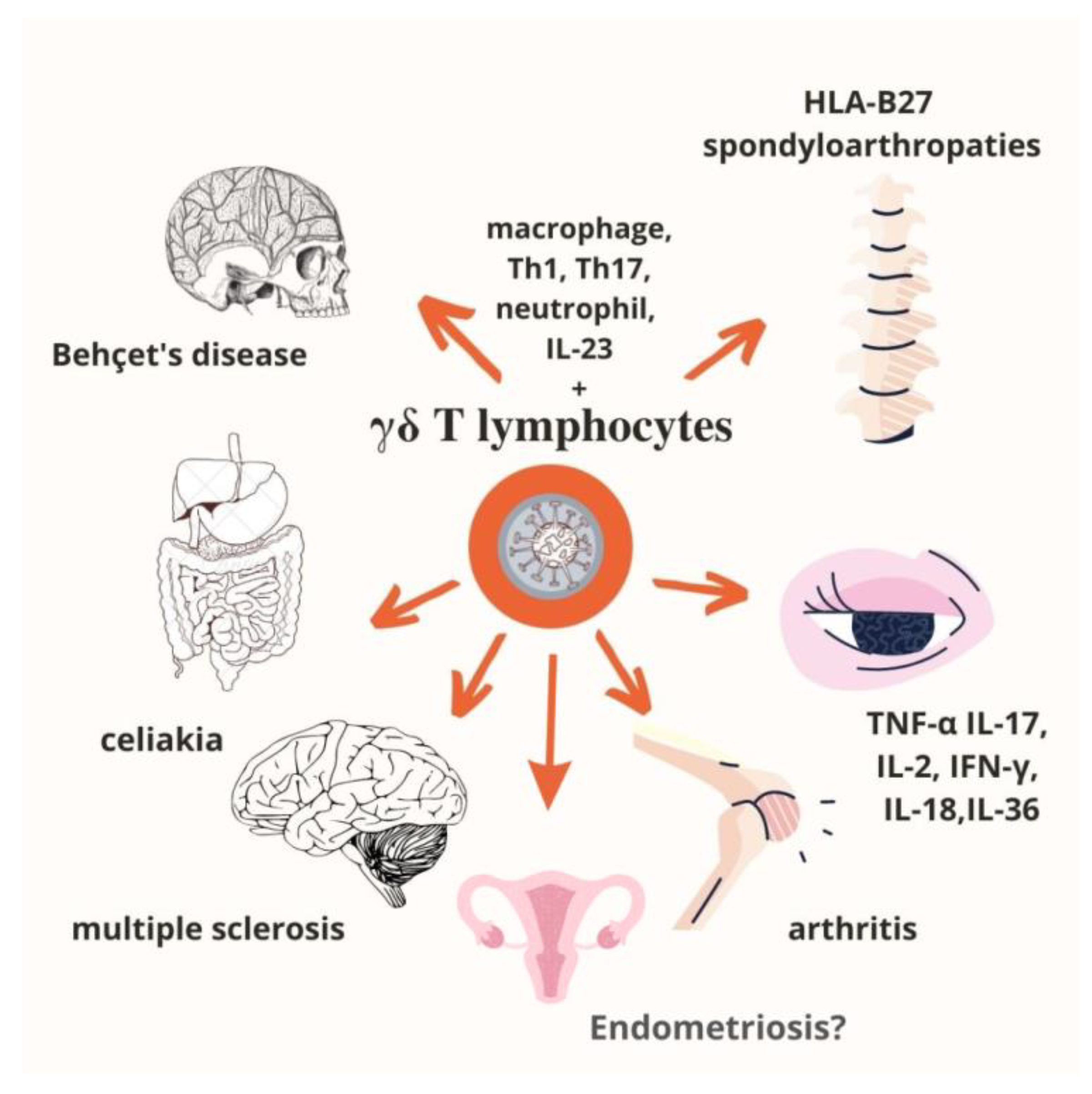
4. γδ T Lymphocytes in Autoimmune Diseases
5. γδ T Lymphocytes in Endometriosis
6. γδ T-Cell-Based Immunotherapy and Its Limitations
7. γδ T Cells—Clinical Trials
8. The Effect of Conventional Therapies on the Function of γδ T Lymphocytes
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raverdeau, M.; Cunningham, S.P.; Harmon, C.; Lynch, L. Γδ T Cells in Cancer: A Small Population of Lymphocytes with Big Implications. Clin. Transl. Immunol. 2019, 8, e01080. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Chung, Y.S.; Kim, T.J. Heterogeneity of Human Γδ T Cells and Their Role in Cancer Immunity. Immune Netw. 2020, 20, e5. [Google Scholar] [CrossRef]
- Chang, Y.C.; Chiang, Y.H.; Hsu, K.; Chuang, C.K.; Kao, C.W.; Chang, Y.F.; Chang, M.C.; Lim, K.H.; Cheng, H.I.; Hsu, Y.N.; et al. Activated Naïve Γδ T Cells Accelerate Deep Molecular Response to BCR-ABL Inhibitors in Patients with Chronic Myeloid Leukemia. Blood Cancer J. 2021, 11, 182. [Google Scholar] [CrossRef] [PubMed]
- Hudecek, R.; Kohlova, B.; Siskova, I.; Piskacek, M.; Knight, A. Blocking of EphA2 on Endometrial Tumor Cells Reduces Susceptibility to Vδ1 Gamma-Delta T-Cell-Mediated Killing. Front. Immunol. 2021, 12, 752646. [Google Scholar] [CrossRef] [PubMed]
- Vallvé-Juanico, J.; Houshdaran, S.; Giudice, L.C. The Endometrial Immune Environment of Women with Endometriosis. Hum. Reprod. Update 2019, 25, 564–591. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, G.; Zhang, J.; Wu, X.; Chen, X. The Dual Roles of Human Γδ T Cells: Anti-Tumor or Tumor-Promoting. Front. Immunol. 2020, 11, 619954. [Google Scholar] [CrossRef] [PubMed]
- D’Acquisto, F.; Crompton, T. CD3+CD4-CD8- (Double Negative) T Cells: Saviours or Villains of the Immune Response? Biochem. Pharmacol. 2011, 82, 333–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weimer, P.; Wellbrock, J.; Sturmheit, T.; Oliveira-Ferrer, L.; Ding, Y.; Menzel, S.; Witt, M.; Hell, L.; Schmalfeldt, B.; Bokemeyer, C.; et al. Tissue-Specific Expression of TIGIT, PD-1, TIM-3, and CD39 by Γδ T Cells in Ovarian Cancer. Cells 2022, 11, 964. [Google Scholar] [CrossRef] [PubMed]
- Galati, D.; Zanotta, S.; Bocchino, M.; De Filippi, R.; Pinto, A. The Subtle Interplay between Gamma Delta T Lymphocytes and Dendritic Cells: Is There a Role for a Therapeutic Cancer Vaccine in the Era of Combinatorial Strategies? Cancer Immunol. Immunother. 2021, 70, 1797–1809. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.F.; Duarte, J.D.G.; Ostrouska, S.; Behren, A. Γδ T Cells in the Tumor Microenvironment-Interactions with Other Immune Cells. Front. Immunol. 2022, 13, 894315. [Google Scholar] [CrossRef]
- Zhao, Y.; Niu, C.; Cui, J. Gamma-Delta (Γδ) T Cells: Friend or Foe in Cancer Development? J. Transl. Med. 2018, 16, 3. [Google Scholar] [CrossRef] [Green Version]
- Weulersse, M.; Asrir, A.; Pichler, A.C.; Lemaitre, L.; Braun, M.; Carrié, N.; Joubert, M.-V.; Le Moine, M.; Do Souto, L.; Gaud, G.; et al. Eomes-Dependent Loss of the Co-Activating Receptor CD226 Restrains CD8+ T Cell Anti-Tumor Functions and Limits the Efficacy of Cancer Immunotherapy. Immunity 2020, 53, 824–839.e10. [Google Scholar] [CrossRef]
- Maiorano, B.A.; Maiorano, M.F.P.; Lorusso, D.; Maiello, E. Ovarian Cancer in the Era of Immune Checkpoint Inhibitors: State of the Art and Future Perspectives. Cancers 2021, 13, 4438. [Google Scholar] [CrossRef]
- Chardin, L.; Leary, A. Immunotherapy in Ovarian Cancer: Thinking Beyond PD-1/PD-L1. Front. Oncol. 2021, 11, 795547. [Google Scholar] [CrossRef]
- Gentles, A.J.; Newman, A.M.; Liu, C.L.; Bratman, S.V.; Feng, W.; Kim, D.; Nair, V.S.; Xu, Y.; Khuong, A.; Hoang, C.D.; et al. The Prognostic Landscape of Genes and Infiltrating Immune Cells across Human Cancers. Nat. Med. 2015, 21, 938–945. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Shang, W.; Xu, R.; Wu, M.; Zhang, X.; Huang, P.; Wang, F.; Pan, S. Distribution and Functions of Γδ T Cells Infiltrated in the Ovarian Cancer Microenvironment. J. Transl. Med. 2019, 17, 144. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zhang, X.; Xu, R.; Shang, W.; Ming, W.; Wang, F.; Wang, J. Implication of IL-17 Producing ⱭβT and ΓδT Cells in Patients with Ovarian Cancer. Hum. Immunol. 2020, 81, 244–248. [Google Scholar] [CrossRef]
- Almeida-Nunes, D.L.; Mendes-Frias, A.; Silvestre, R.; Dinis-Oliveira, R.J.; Ricardo, S. Immune Tumor Microenvironment in Ovarian Cancer Ascites. Int. J. Mol. Sci. 2022, 23, 10692. [Google Scholar] [CrossRef]
- Foord, E.; Arruda, L.C.M.; Gaballa, A.; Klynning, C.; Uhlin, M. Characterization of Ascites- and Tumor-Infiltrating Γδ T Cells Reveals Distinct Repertoires and a Beneficial Role in Ovarian Cancer. Sci. Transl. Med. 2021, 13, eabb0192. [Google Scholar] [CrossRef]
- Raspollini, M.R.; Castiglione, F.; Rossi Degl’Innocenti, D.; Amunni, G.; Villanucci, A.; Garbini, F.; Baroni, G.; Taddei, G.L. Tumour-Infiltrating Gamma/Delta T-Lymphocytes Are Correlated with a Brief Disease-Free Interval in Advanced Ovarian Serous Carcinoma. Ann. Oncol. 2005, 16, 590–596. [Google Scholar] [CrossRef]
- Rei, M.; Gonçalves-Sousa, N.; Lança, T.; Thompson, R.G.; Mensurado, S.; Balkwill, F.R.; Kulbe, H.; Pennington, D.J.; Silva-Santos, B. Murine CD27(−) Vγ6(+) Γδ T Cells Producing IL-17A Promote Ovarian Cancer Growth via Mobilization of Protumor Small Peritoneal Macrophages. Proc. Natl. Acad. Sci. USA 2014, 111, E3562–E3570. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.; Wu, D.; Ni, C.; Ye, J.; Chen, W.; Hu, G.; Wang, Z.; Wang, C.; Zhang, Z.; Xia, W.; et al. ΓδT17 Cells Promote the Accumulation and Expansion of Myeloid-Derived Suppressor Cells in Human Colorectal Cancer. Immunity 2014, 40, 785–800. [Google Scholar] [CrossRef] [Green Version]
- Chabab, G.; Barjon, C.; Bonnefoy, N.; Lafont, V. Pro-Tumor Γδ T Cells in Human Cancer: Polarization, Mechanisms of Action, and Implications for Therapy. Front. Immunol. 2020, 11, 2186. [Google Scholar] [CrossRef] [PubMed]
- Daley, D.; Zambirinis, C.P.; Seifert, L.; Akkad, N.; Mohan, N.; Werba, G.; Barilla, R.; Torres-Hernandez, A.; Hundeyin, M.; Mani, V.R.K.; et al. Γδ T Cells Support Pancreatic Oncogenesis by Restraining Aβ T Cell Activation. Cell 2016, 166, 1485–1499.e15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, C.; Oberg, H.-H.; Kabelitz, D.; Wesch, D. Phenotype and Regulation of Immunosuppressive Vδ2-Expressing Γδ T Cells. Cell Mol. Life Sci. 2014, 71, 1943–1960. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Zhang, Q.; Ye, J.; Wang, F.; Zhang, Y.; Wevers, E.; Schwartz, T.; Hunborg, P.; Varvares, M.A.; Hoft, D.F.; et al. Tumor-Infiltrating Γδ T Lymphocytes Predict Clinical Outcome in Human Breast Cancer. J. Immunol. 2012, 189, 5029–5036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhang, C. The Role of Human Γδ T Cells in Anti-Tumor Immunity and Their Potential for Cancer Immunotherapy. Cells 2020, 9, 1206. [Google Scholar] [CrossRef] [PubMed]
- Dang, W.-Z.; Li, H.; Jiang, B.; Nandakumar, K.S.; Liu, K.-F.; Liu, L.-X.; Yu, X.-C.; Tan, H.-J.; Zhou, C. Therapeutic Effects of Artesunate on Lupus-Prone MRL/Lpr Mice Are Dependent on T Follicular Helper Cell Differentiation and Activation of JAK2-STAT3 Signaling Pathway. Phytomedicine 2019, 62, 152965. [Google Scholar] [CrossRef]
- Gu-Trantien, C.; Migliori, E.; Buisseret, L.; de Wind, A.; Brohée, S.; Garaud, S.; Noël, G.; Dang Chi, V.L.; Lodewyckx, J.-N.; Naveaux, C.; et al. CXCL13-Producing TFH Cells Link Immune Suppression and Adaptive Memory in Human Breast Cancer. JCI Insight 2017, 2, 91487. [Google Scholar] [CrossRef] [Green Version]
- Curiel, T.J.; Coukos, G.; Zou, L.; Alvarez, X.; Cheng, P.; Mottram, P.; Evdemon-Hogan, M.; Conejo-Garcia, J.R.; Zhang, L.; Burow, M.; et al. Specific Recruitment of Regulatory T Cells in Ovarian Carcinoma Fosters Immune Privilege and Predicts Reduced Survival. Nat. Med. 2004, 10, 942–949. [Google Scholar] [CrossRef]
- Hu, G.; Wu, P.; Cheng, P.; Zhang, Z.; Wang, Z.; Yu, X.; Shao, X.; Wu, D.; Ye, J.; Zhang, T.; et al. Tumor-Infiltrating CD39+ γδ Tregs Are Novel Immunosuppressive T Cells in Human Colorectal Cancer. OncoImmunology 2017, 6, e1277305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo Presti, E.; Dieli, F.; Fourniè, J.J.; Meraviglia, S. Deciphering Human Γδ T Cell Response in Cancer: Lessons from Tumor-Infiltrating Γδ T Cells. Immunol. Rev. 2020, 298, 153–164. [Google Scholar] [CrossRef]
- Patil, R.S.; Shah, S.U.; Shrikhande, S.V.; Goel, M.; Dikshit, R.P.; Chiplunkar, S.V. IL17 Producing ΓδT Cells Induce Angiogenesis and Are Associated with Poor Survival in Gallbladder Cancer Patients. Int. J. Cancer 2016, 139, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Ye, W.; Lan, T.; Zhao, Y.; Liu, X.; Chen, J.; Lai, J.; Chen, S.; Zhong, X.; Wu, X. Characteristic of TIGIT and DNAM-1 Expression on Foxp3+ Γδ T Cells in AML Patients. Biomed. Res. Int. 2020, 2020, 4612952. [Google Scholar] [CrossRef] [PubMed]
- Hysa, E.; Cutolo, C.A.; Gotelli, E.; Pacini, G.; Schenone, C.; Kreps, E.O.; Smith, V.; Cutolo, M. Immunopathophysiology and Clinical Impact of Uveitis in Inflammatory Rheumatic Diseases: An Update. Eur. J. Clin. Investig. 2021, 51, e13572. [Google Scholar] [CrossRef] [PubMed]
- Tomogane, M.; Sano, Y.; Shimizu, D.; Shimizu, T.; Miyashita, M.; Toda, Y.; Hosogi, S.; Tanaka, Y.; Kimura, S.; Ashihara, E. Human Vγ9Vδ2 T Cells Exert Anti-Tumor Activity Independently of PD-L1 Expression in Tumor Cells. Biochem. Biophys. Res. Commun. 2021, 573, 132–139. [Google Scholar] [CrossRef]
- Talukdar, A.; Rai, R.; Aparna Sharma, K.; Rao, D.N.; Sharma, A. Peripheral Gamma Delta T Cells Secrete Inflammatory Cytokines in Women with Idiopathic Recurrent Pregnancy Loss. Cytokine 2018, 102, 117–122. [Google Scholar] [CrossRef]
- Girard, P.; Charles, J.; Cluzel, C.; Degeorges, E.; Manches, O.; Plumas, J.; De Fraipont, F.; Leccia, M.-T.; Mouret, S.; Chaperot, L.; et al. The Features of Circulating and Tumor-Infiltrating Γδ T Cells in Melanoma Patients Display Critical Perturbations with Prognostic Impact on Clinical Outcome. Oncoimmunology 2019, 8, 1601483. [Google Scholar] [CrossRef] [Green Version]
- Meraviglia, S.; Lo Presti, E.; Tosolini, M.; La Mendola, C.; Orlando, V.; Todaro, M.; Catalano, V.; Stassi, G.; Cicero, G.; Vieni, S.; et al. Distinctive Features of Tumor-Infiltrating Γδ T Lymphocytes in Human Colorectal Cancer. Oncoimmunology 2017, 6, e1347742. [Google Scholar] [CrossRef] [Green Version]
- Viey, E.; Fromont, G.; Escudier, B.; Morel, Y.; Rocha, S.D.; Chouaib, S.; Caignard, A. Phosphostim-Activated Γδ T Cells Kill Autologous Metastatic Renal Cell Carcinoma. J. Immunol. 2005, 174, 1338–1347. [Google Scholar] [CrossRef]
- Barros, M.d.S.; de Araújo, N.D.; Magalhães-Gama, F.; Pereira Ribeiro, T.L.; Alves Hanna, F.S.; Tarragô, A.M.; Malheiro, A.; Costa, A.G. Γδ T Cells for Leukemia Immunotherapy: New and Expanding Trends. Front. Immunol. 2021, 12, 729085. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.T.; Maverakis, E.; Eberl, M.; Adamopoulos, I.E. Γδ T Cells in Rheumatic Diseases: From Fundamental Mechanisms to Autoimmunity. Semin. Immunopathol. 2019, 41, 595–605. [Google Scholar] [CrossRef] [PubMed]
- Shiromizu, C.M.; Jancic, C.C. Γδ T Lymphocytes: An Effector Cell in Autoimmunity and Infection. Front. Immunol. 2018, 9, 2389. [Google Scholar] [CrossRef] [PubMed]
- Blink, S.E.; Miller, S.D. The Contribution of Γδ T Cells to the Pathogenesis of EAE and MS. Curr. Mol. Med. 2009, 9, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhou, Y.; Sun, X.; Bian, Y.; Wang, K.; Guo, Q.; Wang, Q.; Qiu, F. Interleukin-2 Maintains the Survival of Interleukin-17+ Gamma/Delta T Cells in Inflammation and Autoimmune Diseases. Int. Immunopharmacol. 2020, 86, 106721. [Google Scholar] [CrossRef] [PubMed]
- Roark, C.L.; Simonian, P.L.; Fontenot, A.P.; Born, W.K.; O’Brien, R.L. Γδ T Cells: An Important Source of IL-17. Curr. OpinImmunol. 2008, 20, 353–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Shen, X.; Ding, C.; Qi, C.; Li, K.; Li, X.; Jala, V.R.; Zhang, H.; Wang, T.; Zheng, J.; et al. Pivotal Role of Dermal IL-17-Producing Γδ T Cells in Skin Inflammation. Immunity 2011, 35, 596–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cua, D.J.; Tato, C.M. Innate IL-17-Producing Cells: The Sentinels of the Immune System. Nat. Rev. Immunol. 2010, 10, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Hirota, K.; Duarte, J.H.; Veldhoen, M.; Hornsby, E.; Li, Y.; Cua, D.J.; Ahlfors, H.; Wilhelm, C.; Tolaini, M.; Menzel, U.; et al. Fate Mapping of IL-17-Producing T Cells in Inflammatory Responses. Nat. Immunol. 2011, 12, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Petermann, F.; Rothhammer, V.; Claussen, M.C.; Haas, J.D.; Blanco, L.R.; Heink, S.; Prinz, I.; Hemmer, B.; Kuchroo, V.K.; Oukka, M.; et al. Γδ T Cells Enhance Autoimmunity by Restraining Regulatory T Cell Responses via an Interleukin-23-Dependent Mechanism. Immunity 2010, 33, 351–363. [Google Scholar] [CrossRef]
- Itoh, S.; Kimura, N.; Axtell, R.C.; Velotta, J.B.; Gong, Y.; Wang, X.; Kajiwara, N.; Nambu, A.; Shimura, E.; Adachi, H.; et al. Interleukin-17 Accelerates Allograft Rejection by Suppressing Regulatory T Cell Expansion. Circulation 2011, 124, S187–S196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, S.; Shilpi; Lal, G. Role of Gamma-Delta (Γδ) T Cells in Autoimmunity. J. Leukoc. Biol. 2015, 97, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Abbasova, V.; Gül, A.; Saruhan-Direskeneli, G. IL-17A and IFN-γ Are Up-Regulated in CD4 and Γδ T Cells in Active Behcet’s Disease Patients. Immunol. Lett. 2022, 242, 37–45. [Google Scholar] [CrossRef]
- Hasan, M.S.; Bergmeier, L.A.; Petrushkin, H.; Fortune, F. Gamma Delta (Γδ) T Cells and Their Involvement in Behçet’s Disease. J. Immunol. Res. 2015, 2015, 705831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kühl, A.A.; Pawlowski, N.N.; Grollich, K.; Blessenohl, M.; Westermann, J.; Zeitz, M.; Loddenkemper, C.; Hoffmann, J.C. Human Peripheral Γδ T Cells Possess Regulatory Potential. Immunology 2009, 128, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Peng, G.; Wang, H.Y.; Peng, W.; Kiniwa, Y.; Seo, K.H.; Wang, R.-F. Tumor-Infiltrating Γδ T Cells Suppress T and Dendritic Cell Function via Mechanisms Controlled by a Unique Toll-like Receptor Signaling Pathway. Immunity 2007, 27, 334–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, D.; Shen, M.; Li, X.; Sun, L. Roles of Γδ T Cells in the Pathogenesis of Autoimmune Diseases. Clin. Dev. Immunol. 2013, 2013, 985753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, G.; Shao, L.; Wang, Y.; Chen, C.Y.; Huang, D.; Yao, S.; Zhan, X.; Sicard, H.; Wang, R.; Chen, Z.W. Phosphoantigen-Activated Vγ2Vδ2 T Cells Antagonize IL-2–Induced CD4+CD25+Foxp3+ T Regulatory Cells in Mycobacterial Infection. Blood 2009, 113, 837–845. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Wu, C.-Y. CD4+CD25+ Treg Cells Inhibit Human Memory Γδ T Cells to Produce IFN-γ in Response to M Tuberculosis Antigen ESAT-6. Blood 2008, 111, 5629–5636. [Google Scholar] [CrossRef] [PubMed]
- Sinaii, N.; Cleary, S.D.; Ballweg, M.L.; Nieman, L.K.; Stratton, P. High Rates of Autoimmune and Endocrine Disorders, Fibromyalgia, Chronic Fatigue Syndrome and Atopic Diseases among Women with Endometriosis: A Survey Analysis. Hum. Reprod. 2002, 17, 2715–2724. [Google Scholar] [CrossRef] [Green Version]
- Suszczyk, D.; Skiba, W.; Jakubowicz-Gil, J.; Kotarski, J.; Wertel, I. The Role of Myeloid-Derived Suppressor Cells (MDSCs) in the Development and/or Progression of Endometriosis-State of the Art. Cells 2021, 10, 677. [Google Scholar] [CrossRef] [PubMed]
- Agostinis, C.; Balduit, A.; Mangogna, A.; Zito, G.; Romano, F.; Ricci, G.; Kishore, U.; Bulla, R. Immunological Basis of the Endometriosis: The Complement System as a Potential Therapeutic Target. Front. Immunol. 2021, 11, 599117. [Google Scholar] [CrossRef] [PubMed]
- Shigesi, N.; Kvaskoff, M.; Kirtley, S.; Feng, Q.; Fang, H.; Knight, J.C.; Missmer, S.A.; Rahmioglu, N.; Zondervan, K.T.; Becker, C.M. The Association between Endometriosis and Autoimmune Diseases: A Systematic Review and Meta-Analysis. Hum. Reprod. Update 2019, 25, 486–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gogacz, M.; Winkler, I.; Bojarska-Junak, A.; Tabarkiewicz, J.; Semczuk, A.; Rechberger, T.; Adamiak, A. Increased Percentage of Th17 Cells in Peritoneal Fluid Is Associated with Severity of Endometriosis. J. Reprod. Immunol. 2016, 117, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Izumi, G.; Koga, K.; Takamura, M.; Makabe, T.; Satake, E.; Takeuchi, A.; Taguchi, A.; Urata, Y.; Fujii, T.; Osuga, Y. Involvement of Immune Cells in the Pathogenesis of Endometriosis. J. Obstet. Gynaecol. Res. 2018, 44, 191–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, C.; Zhao, P.; Xiao, Z.; Han, X.; Fu, F.; Fu, L. Γδ T Cells in Cancer Immunotherapy. Oncotarget 2016, 8, 8900–8909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, J.; Yin, H. Gamma Delta (Γδ) T Cells in Cancer Immunotherapy; Where It Comes from, Where It Will Go? Eur. J. Pharmacol. 2022, 919, 174803. [Google Scholar] [CrossRef] [PubMed]
- Sebestyen, Z.; Prinz, I.; Déchanet-Merville, J.; Silva-Santos, B.; Kuball, J. Translating Gammadelta (Γδ) T Cells and Their Receptors into Cancer Cell Therapies. Nat. Rev. Drug Discov. 2020, 19, 169–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazdanifar, M.; Barbarito, G.; Bertaina, A.; Airoldi, I. Γδ T cells: The ideal tool for cancer Immunotherapy. Cells 2020, 9, 1305. [Google Scholar] [CrossRef] [PubMed]
- Brandes, M.; Willimann, K.; Moser, B. Professional Antigen-Presentation Function by Human Gammadelta T Cells. Science 2005, 309, 264–268. [Google Scholar] [CrossRef] [PubMed]
- IClyCO Influence of Chemotherapy (Carboplatin and Taxol) on the Ex Vivo Expansion and Functional Capacity of Gamma-Delta T Cells in Patients With Epithelial Ovarian Cancer.—ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/show/NCT01606358 (accessed on 20 November 2022).
- Immunotherapy of Epithelial Ovarian Cancer Using Autologous Gamma Delta T-Cells. Available online: https://gtr.ukri.org/projects?ref=MR%2FM024733%2F1&fbclid=IwAR0I3odEZbSusWBEUACFshdwpfA1aya2NC0txi27SQ1bfOYueBsIcrQXiVg#/tabOverview (accessed on 20 November 2022).
- Γδ T Cell Immunotherapy for Treatment of Breast Cancer —ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/show/NCT02418481 (accessed on 20 November 2022).
- Weidong, H. An Open-Label, Phase 1/2 Study of Allogeneic Γδ T Cell Therapy for the Treatment of Solid Tumors; clinicaltrials.gov. 2022. Available online: https://clinicaltrials.gov/ct2/show/NCT04765462 (accessed on 20 November 2022).
- Novel Gamma-Delta (γδ)T Cell Therapy for Treatment of Patients With Newly Diagnosed Glioblastoma (DRI)—ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/show/NCT04165941 (accessed on 20 November 2022).
- Allogeneic Γδ T Cell Therapy for the Treatment of Solid Tumors—ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/show/study/NCT04765462?cond=%CE%B3%CE%B4&draw=2&rank=10 (accessed on 20 November 2022).
- Park, J.H.; Lee, H.K. Function of Γδ T Cells in Tumor Immunology and Their Application to Cancer Therapy. Exp. Mol. Med. 2021, 53, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Tomogane, M.; Miyashita, M.; Ukimura, O.; Ashihara, E. Low Dose Gemcitabine Increases the Cytotoxicity of Human Vγ9Vδ2 T Cells in Bladder Cancer Cells in Vitro and in an Orthotopic Xenograft Model. Oncoimmunology 2018, 7, e1424671. [Google Scholar] [CrossRef]
- Beatson, R.E.; Parente-Pereira, A.C.; Halim, L.; Cozzetto, D.; Hull, C.; Whilding, L.M.; Martinez, O.; Taylor, C.A.; Obajdin, J.; Luu Hoang, K.N.; et al. TGF-Β1 Potentiates Vγ9Vδ2 T Cell Adoptive Immunotherapy of Cancer. Cell Rep. Med. 2021, 2, 100473. [Google Scholar] [CrossRef] [PubMed]
- Mattarollo, S.R.; Kenna, T.; Nieda, M.; Nicol, A.J. Chemotherapy and Zoledronate Sensitize Solid Tumour Cells to Vgamma9Vdelta2 T Cell Cytotoxicity. Cancer Immunol. Immunother. 2007, 56, 1285–1297. [Google Scholar] [CrossRef] [PubMed]
- Kabelitz, D.; Serrano, R.; Kouakanou, L.; Peters, C.; Kalyan, S. Cancer Immunotherapy with Γδ T Cells: Many Paths Ahead of Us. Cell Mol. Immunol. 2020, 17, 925–939. [Google Scholar] [CrossRef]
- Lamb, L.S.; Bowersock, J.; Dasgupta, A.; Gillespie, G.Y.; Su, Y.; Johnson, A.; Spencer, H.T. Engineered Drug Resistant Γδ T Cells Kill Glioblastoma Cell Lines during a Chemotherapy Challenge: A Strategy for Combining Chemo- and Immunotherapy. PLoS ONE 2013, 8, e51805. [Google Scholar] [CrossRef] [PubMed]
- Bustos, X.; Snedal, S.; Tordesillas, L.; Pelle, E.; Abate-Daga, D. Γδ T Cell-Based Adoptive Cell Therapies Against Solid Epithelial Tumors. Cancer J. 2022, 28, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Chiu, Y.-H.; Chiu, S.-C.; Cho, D.-Y.; Lee, L.-M.; Wen, Y.-C.; Whang-Peng, J.; Hsiao, C.-H.; Shih, P.-H. Gamma/Delta T-Cells Enhance Carboplatin-Induced Cytotoxicity Towards Advanced Bladder Cancer Cells. Anticancer Res. 2020, 40, 5221–5227. [Google Scholar] [CrossRef]
- Ma, Y.; Aymeric, L.; Locher, C.; Mattarollo, S.R.; Delahaye, N.F.; Pereira, P.; Boucontet, L.; Apetoh, L.; Ghiringhelli, F.; Casares, N.; et al. Contribution of IL-17-Producing Gamma Delta T Cells to the Efficacy of Anticancer Chemotherapy. J. Exp. Med. 2011, 208, 491–503. [Google Scholar] [CrossRef] [PubMed]
- Joalland, N.; Lafrance, L.; Oullier, T.; Marionneau-Lambot, S.; Loussouarn, D.; Jarry, U.; Scotet, E. Combined Chemotherapy and Allogeneic Human Vγ9Vδ2 T Lymphocyte-Immunotherapies Efficiently Control the Development of Human Epithelial Ovarian Cancer Cells in vivo. Oncoimmunology 2019, 8, e1649971. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, Z.; Li, S.; Li, B.; Sun, L.; Li, H.; Lin, P.; Wang, S.T.; Zhou, X.; Ye, Z. Decitabine Enhances Vγ9Vδ2 T Cell-Mediated Cytotoxic Effects on Osteosarcoma Cells via the NKG2DL–NKG2D Axis. Front. Immunol. 2018, 9, 1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capsomidis, A.; Benthall, G.; Van Acker, H.H.; Fisher, J.; Kramer, A.M.; Abeln, Z.; Majani, Y.; Gileadi, T.; Wallace, R.; Gustafsson, K.; et al. Chimeric Antigen Receptor-Engineered Human Gamma Delta T Cells: Enhanced Cytotoxicity with Retention of Cross Presentation. Mol. Ther. 2018, 26, 354–365. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Vδ T in EMS | Implications of Vδ T in EMS |
|---|---|
| The number of circulating Vδ1 T cells was significantly reduced in women suffering from endometriosis [4]. An increased proportion of Vδ1 T cells in the eutopic and ectopic layers of the endometrium in patients with EMS compared to the controls [4]. | Vδ1 T cells may be involved in:
|
| Vδ T cells are an important source of IL-17 [63] | IL-17A:
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pawłowska, A.; Natochina, Y.; Zardzewiały, W.; Skiba, W.; Włodarczyk, K.; Maciejczyk, A.; Suszczyk, D.; Wertel, I. γδ T Lymphocytes as a Double-Edged Sword—State of the Art in Gynecological Diseases. Int. J. Mol. Sci. 2022, 23, 14797. https://doi.org/10.3390/ijms232314797
Pawłowska A, Natochina Y, Zardzewiały W, Skiba W, Włodarczyk K, Maciejczyk A, Suszczyk D, Wertel I. γδ T Lymphocytes as a Double-Edged Sword—State of the Art in Gynecological Diseases. International Journal of Molecular Sciences. 2022; 23(23):14797. https://doi.org/10.3390/ijms232314797
Chicago/Turabian StylePawłowska, Anna, Yelizaveta Natochina, Witold Zardzewiały, Wiktoria Skiba, Karolina Włodarczyk, Aleksandra Maciejczyk, Dorota Suszczyk, and Iwona Wertel. 2022. "γδ T Lymphocytes as a Double-Edged Sword—State of the Art in Gynecological Diseases" International Journal of Molecular Sciences 23, no. 23: 14797. https://doi.org/10.3390/ijms232314797