
Aspalathin and Other Rooibos Flavonoids Trapped α-Dicarbonyls and Inhibited Formation of Advanced Glycation End Products In Vitro

Abstract
:1. Introduction
2. Results and Discussion
2.1. Phytochemical Profile of Rooibos Extracts
2.2. Quantification of Flavonoids
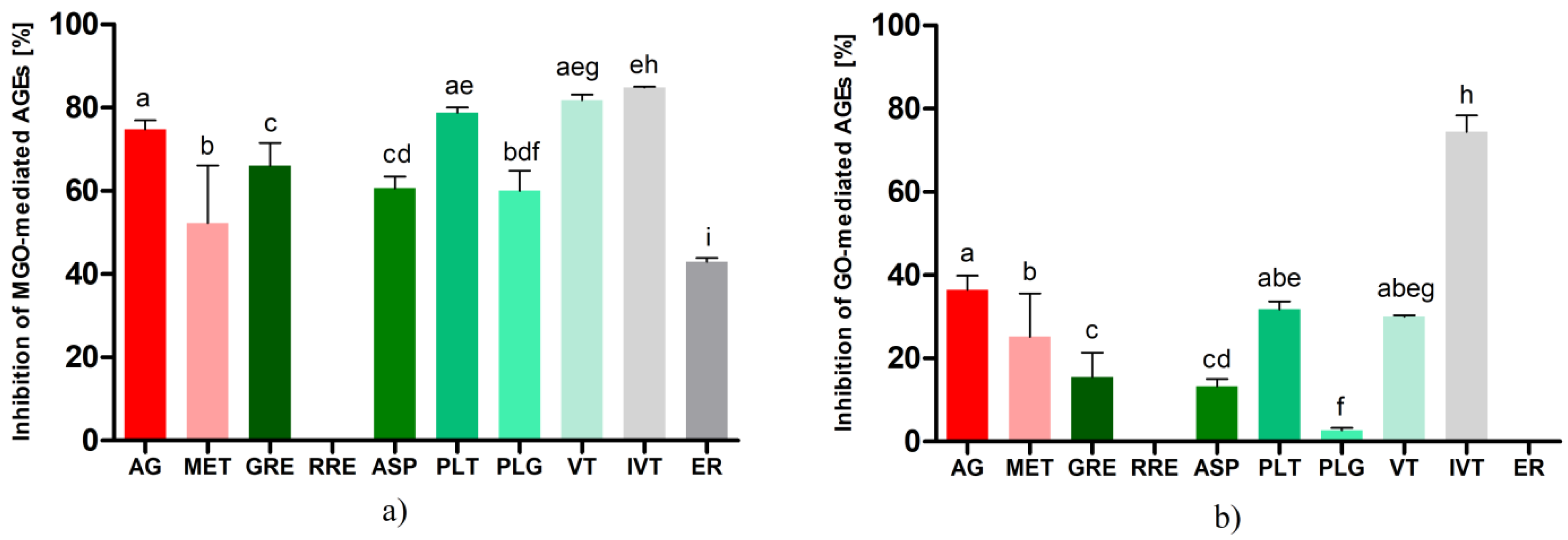
2.3. Inhibition of Glycation by Rooibos Flavonoids and Related Compounds
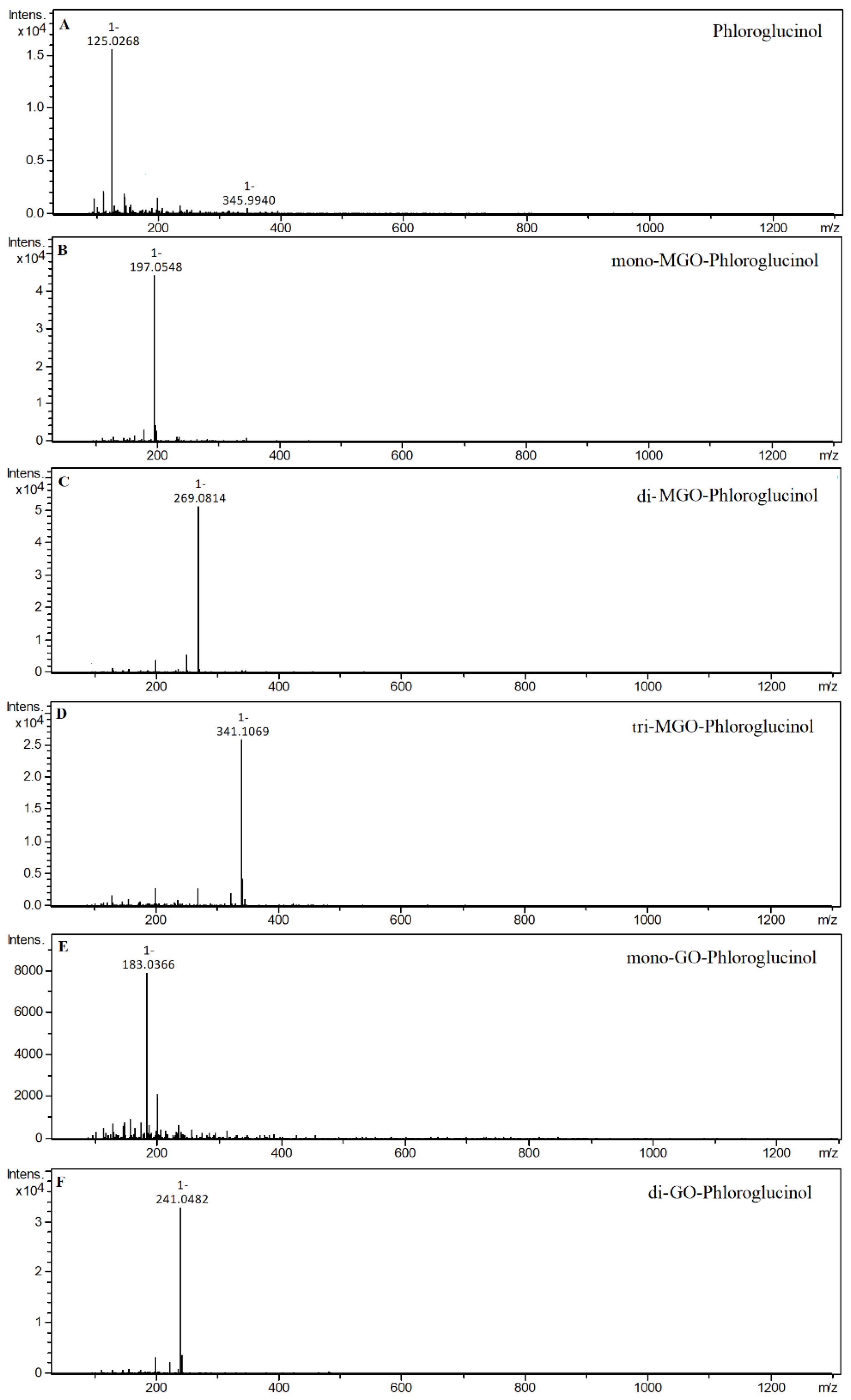
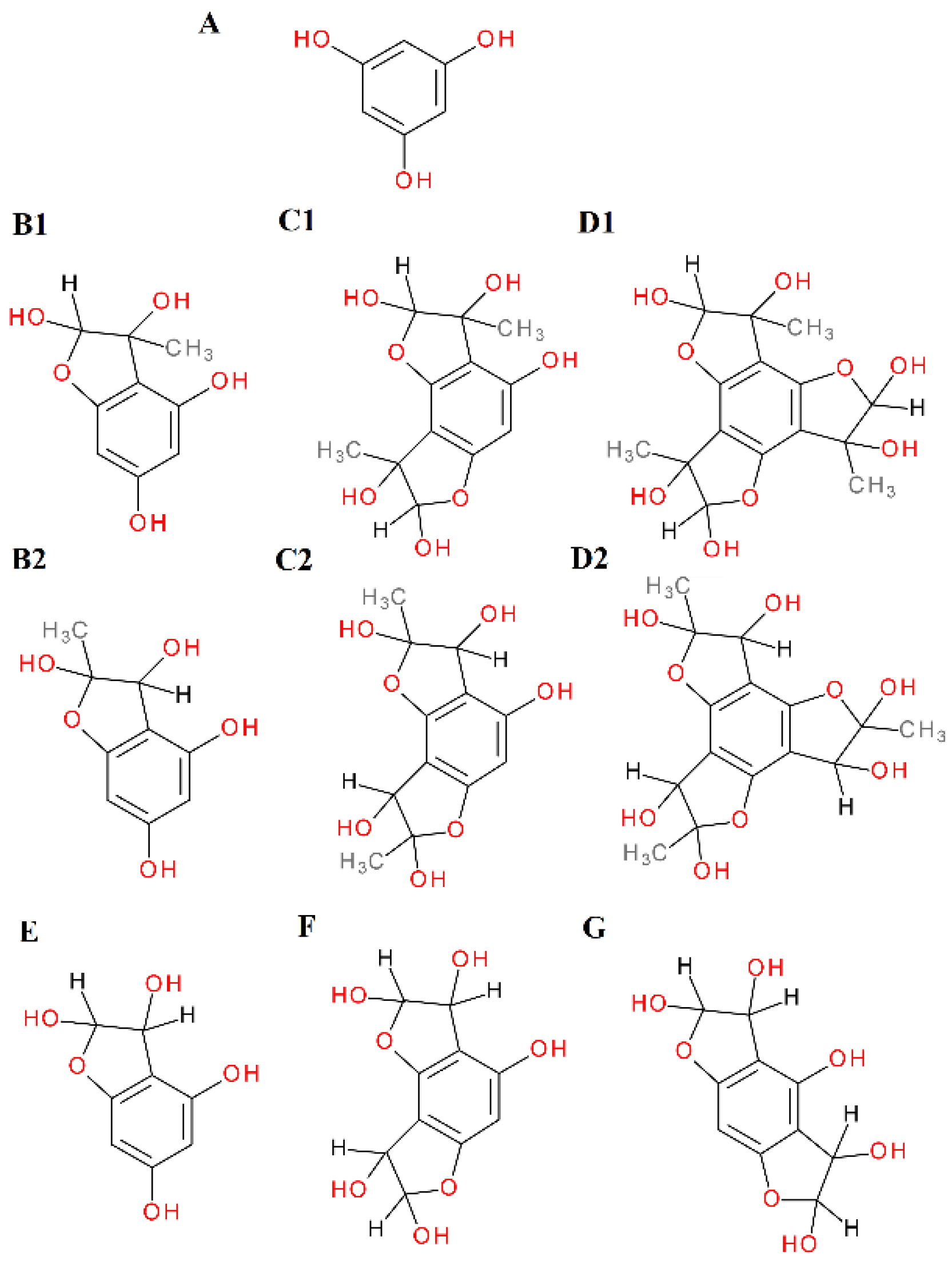
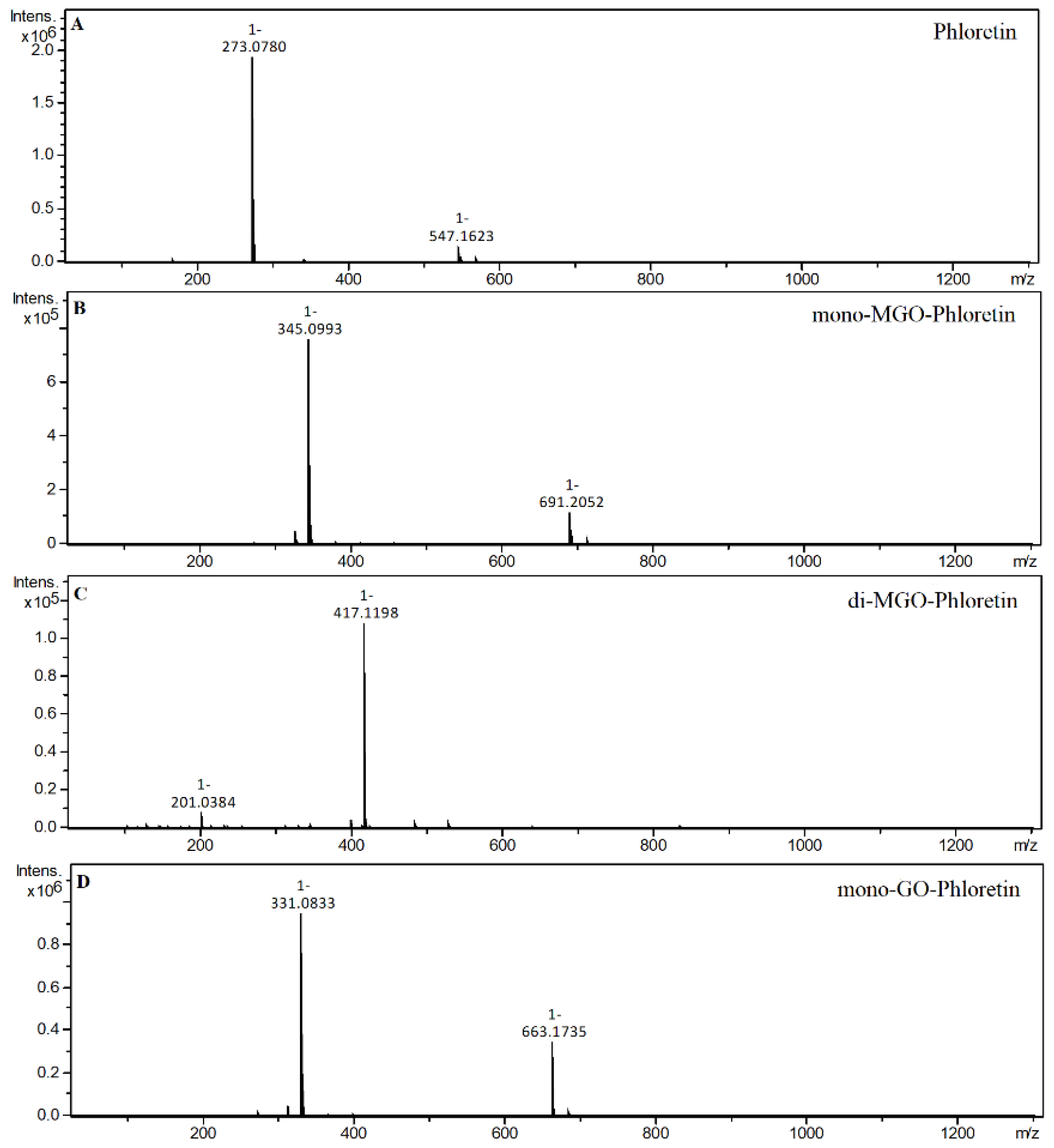
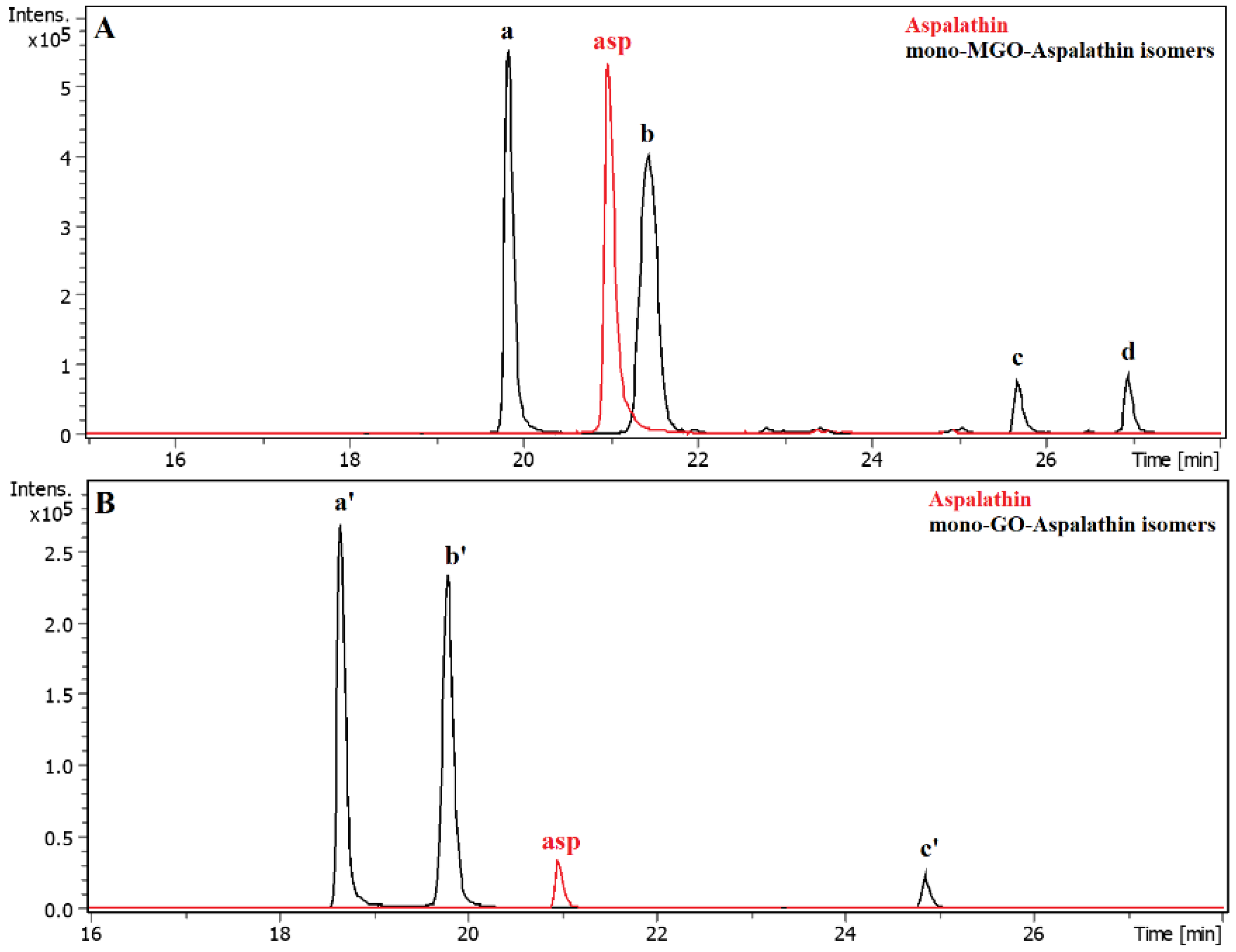
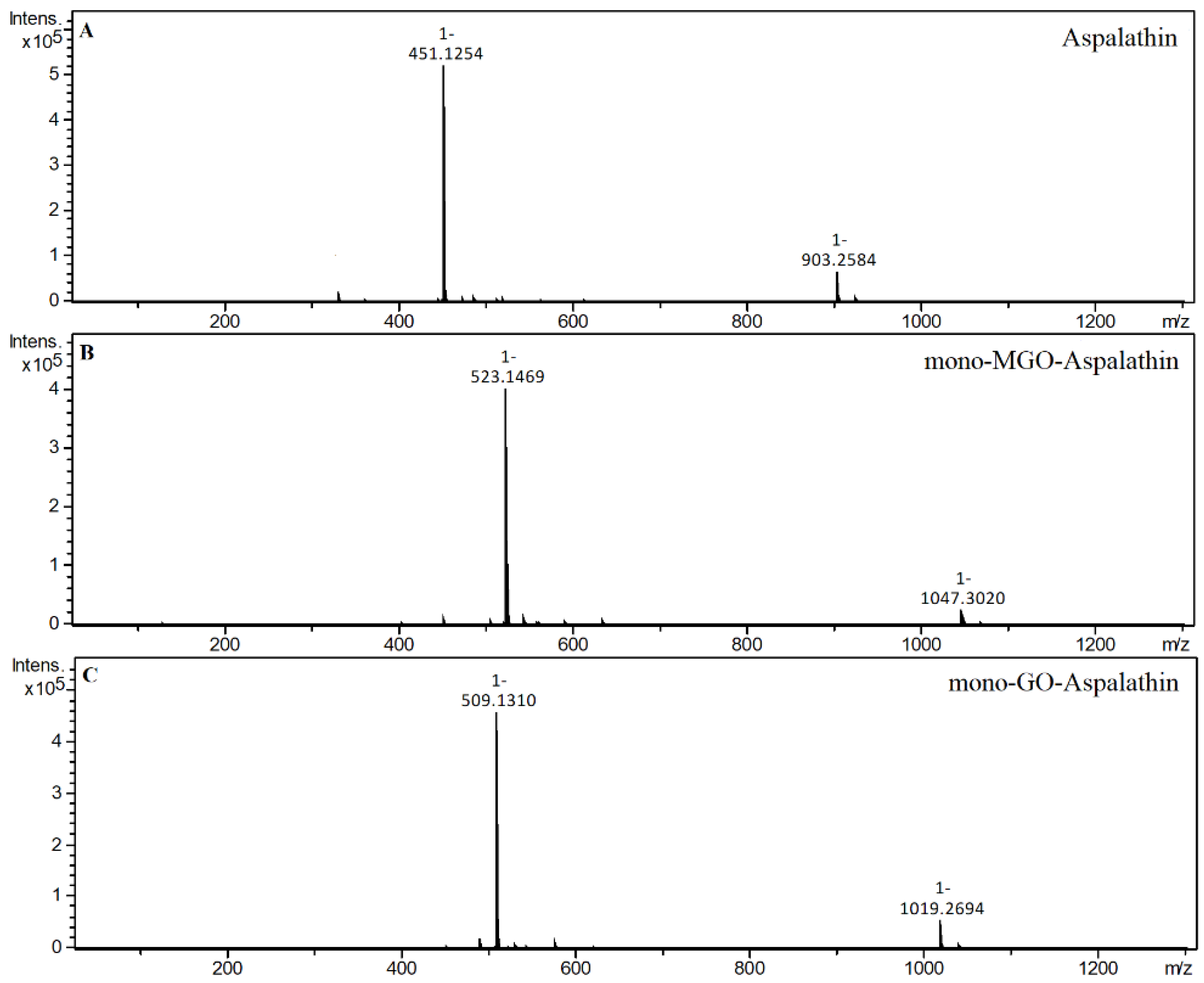
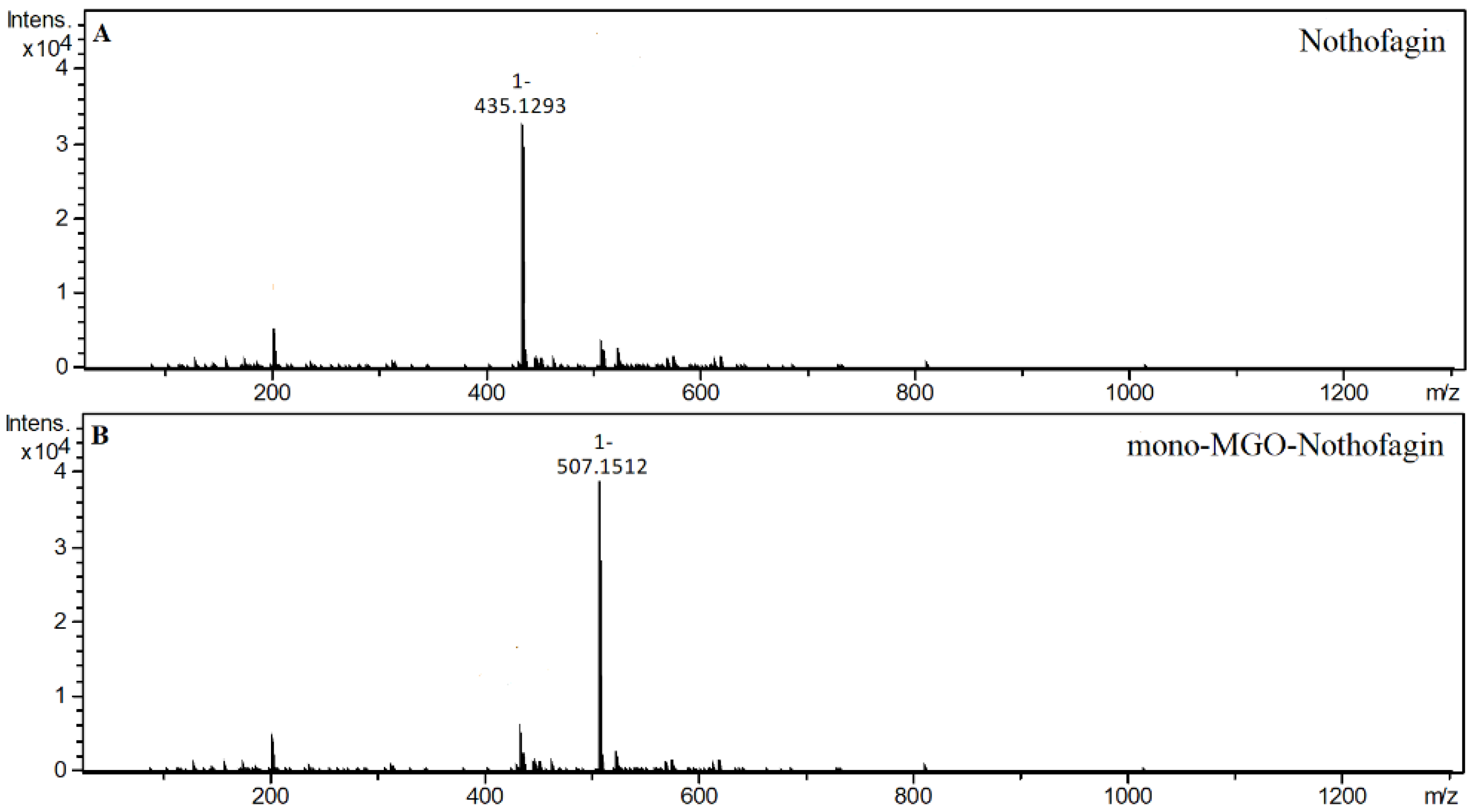
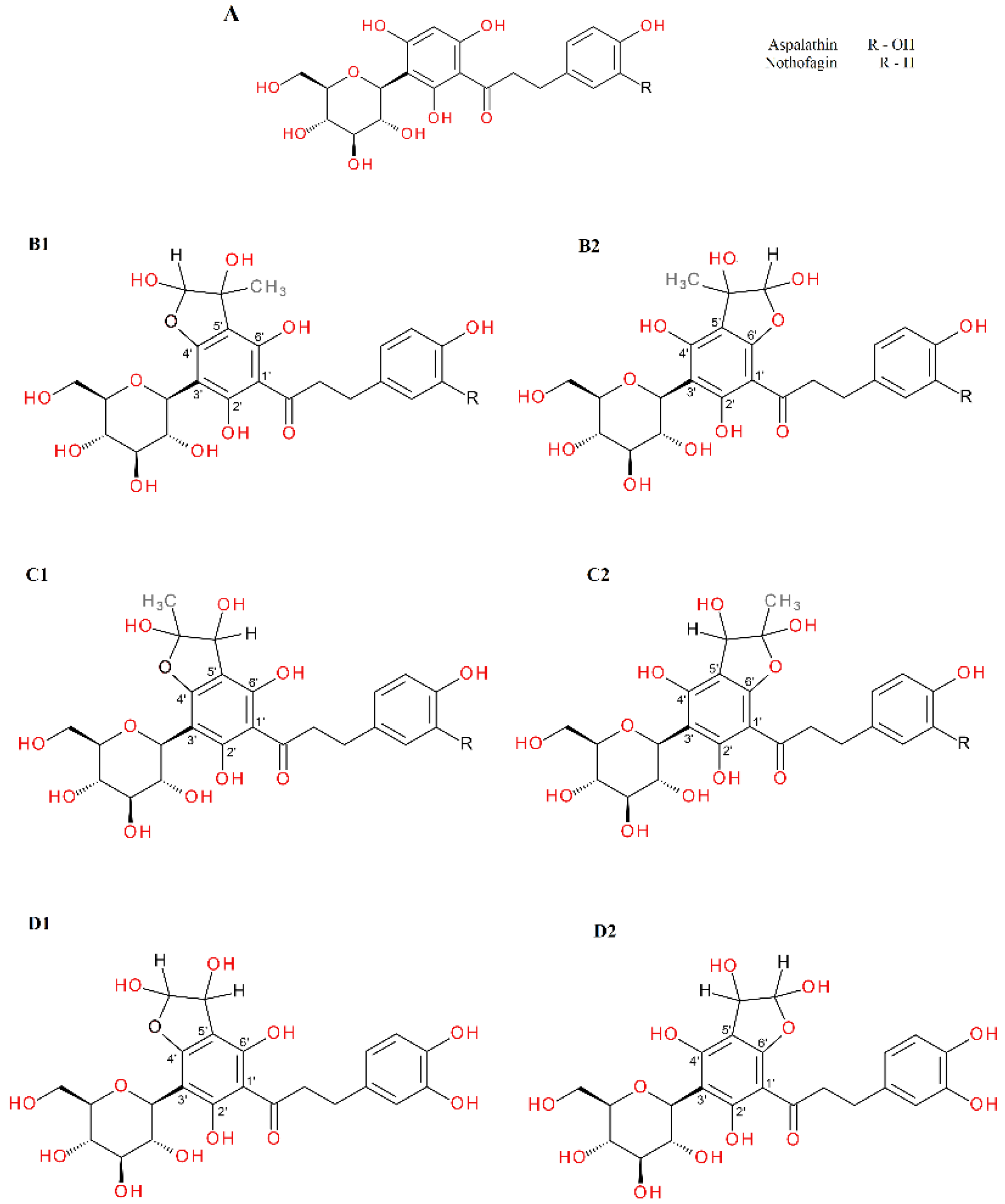
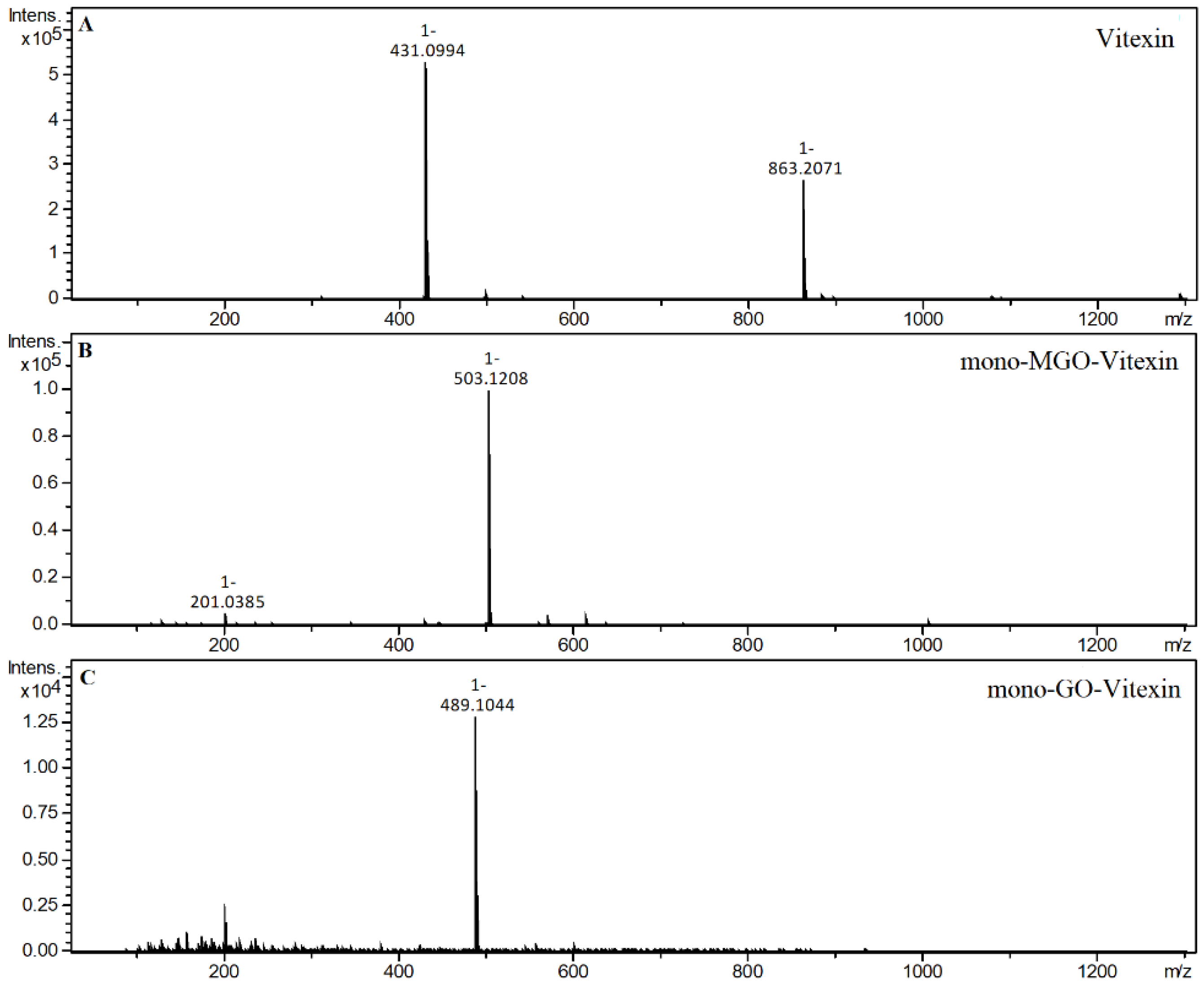
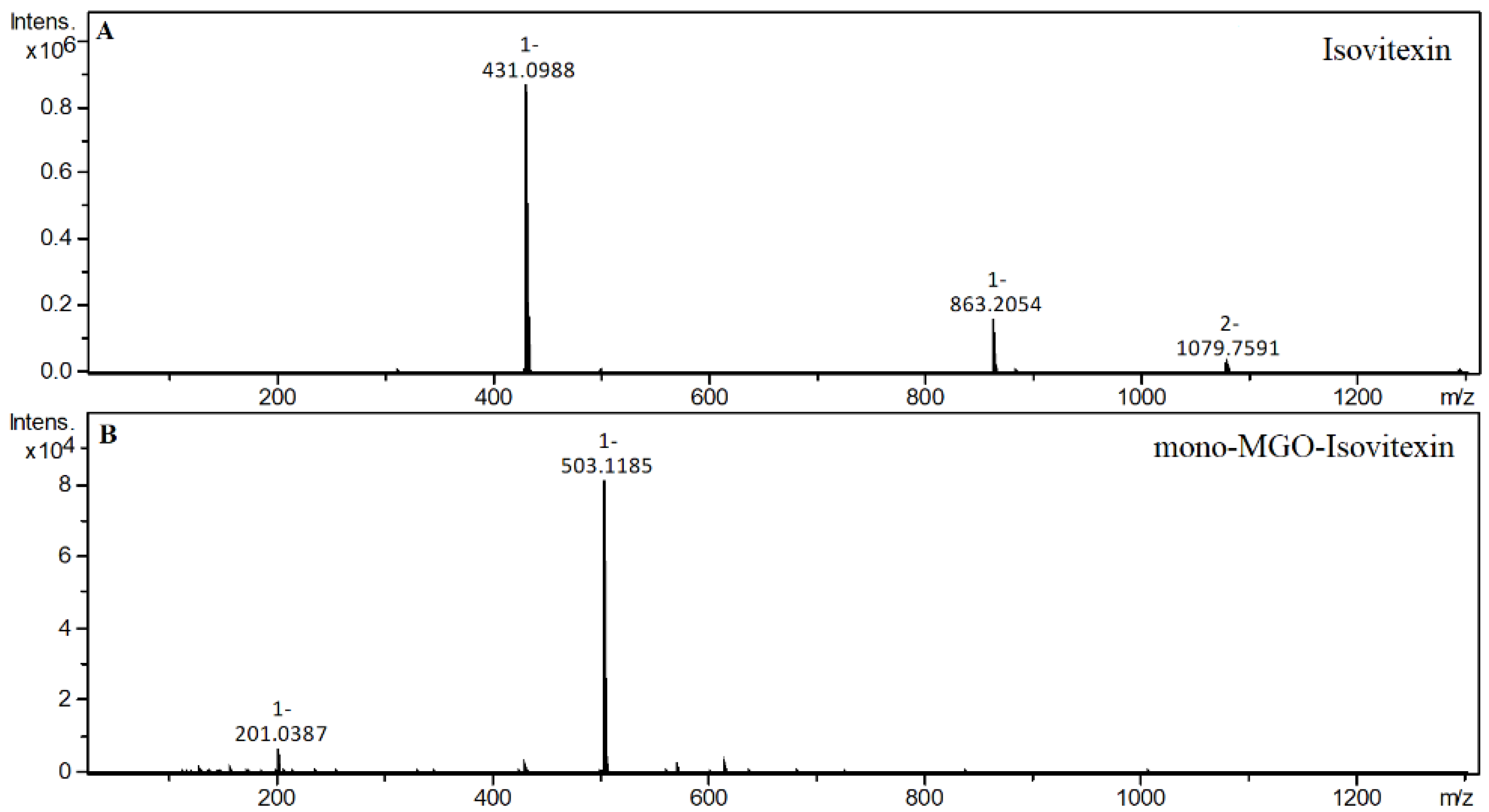
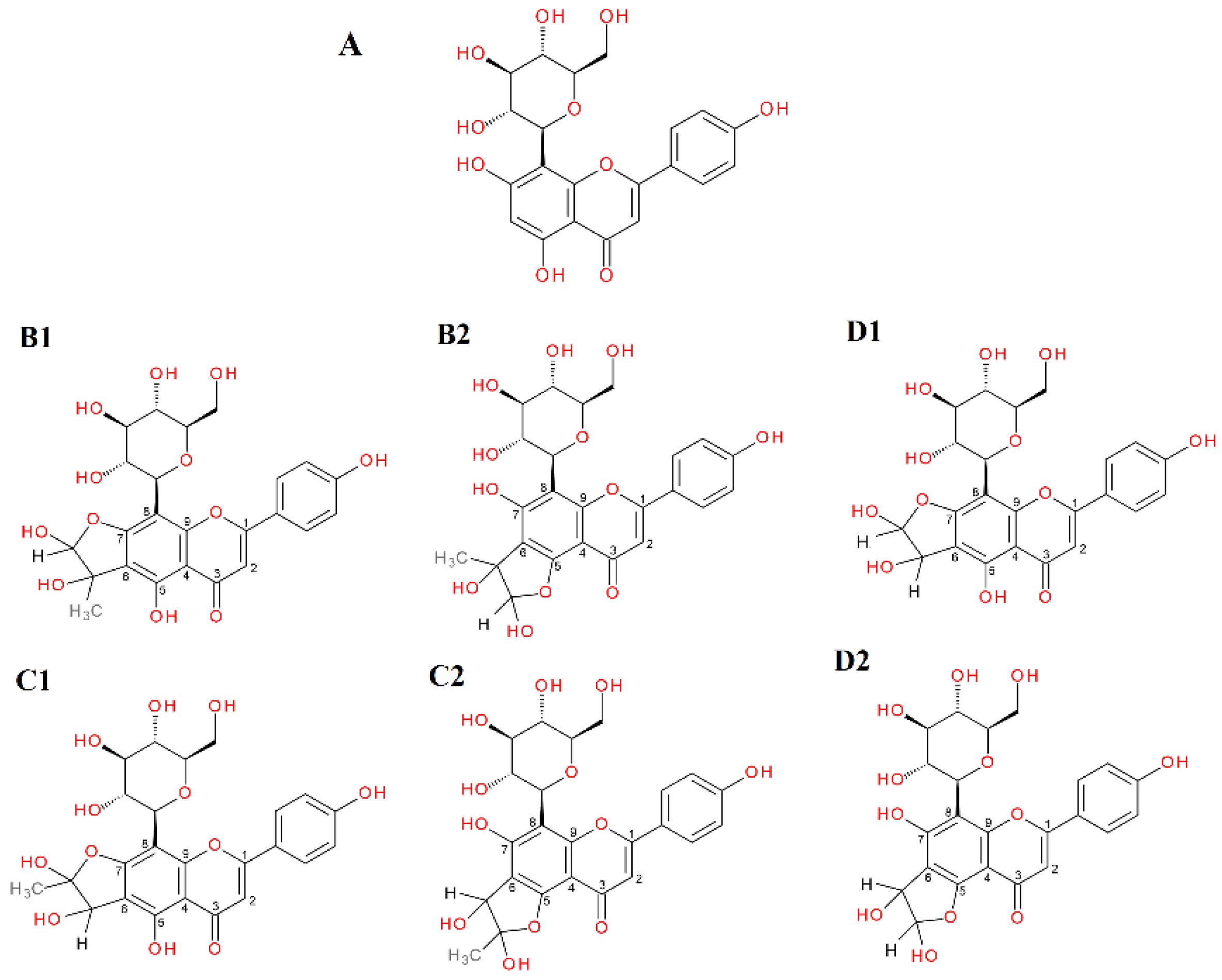
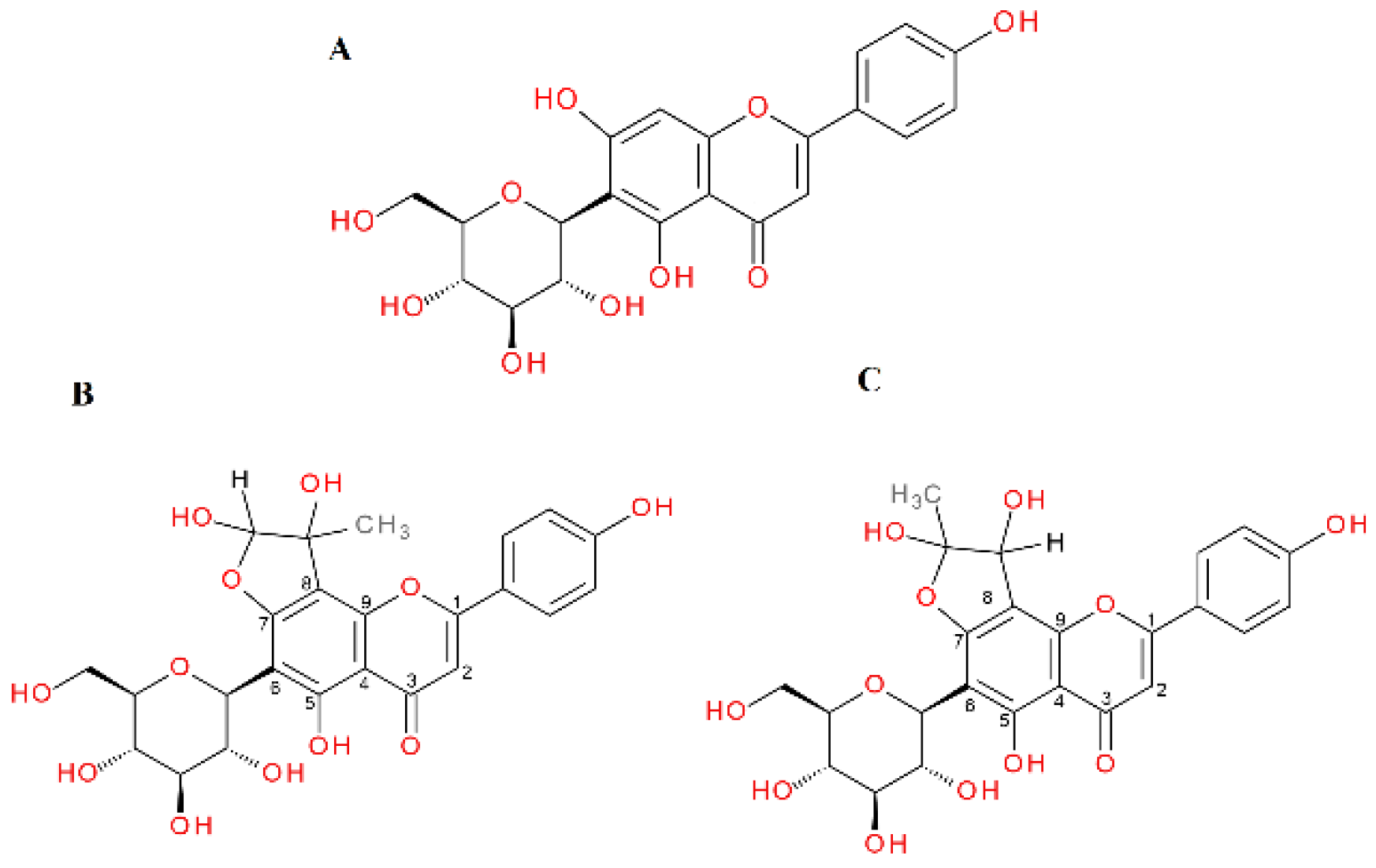
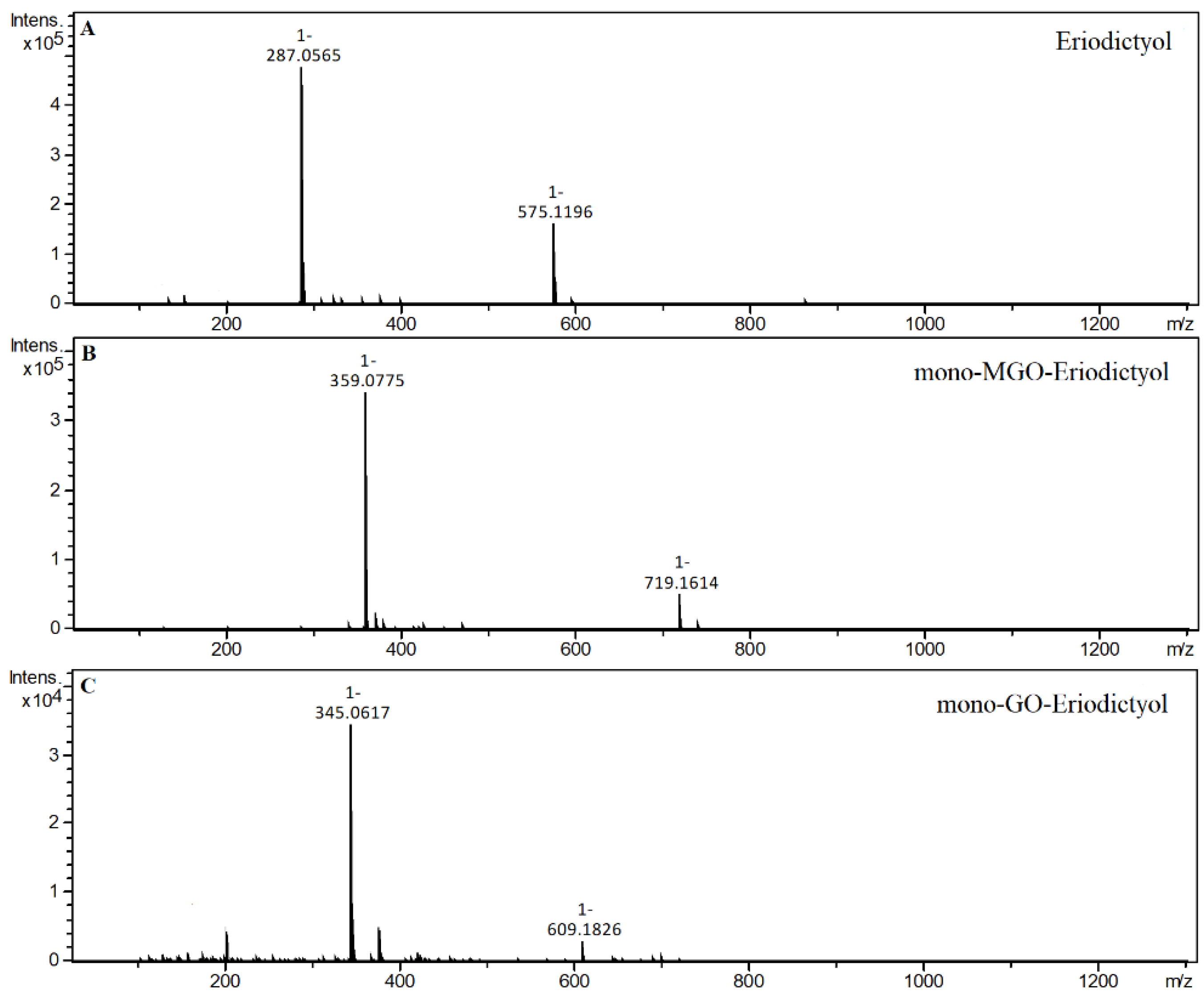
2.4. α-Dicarbonyl Compound Trapping and Adduct Analysis
3. Materials and Methods
3.1. Plant Material
3.2. Preparation of Extracts
3.3. Chemicals
3.4. Phytochemical Profile of Extracts
3.5. Quantification of Flavonoids
3.6. Glycation Process and Fluorescence Measurement of AGEs
3.6.1. In Vitro Glycation Model
3.6.2. Antiglycation Assay
3.7. α-Dicarbonyl Trapping and Adduct Analysis
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ye, S.; Matthan, N.R.; Lamon-Fava, S.; Aguilar, G.S.; Turner, J.R.; Walker, M.E.; Chai, Z.; Lakshman, S.; Urban, J.F.; Lichtenstein, A.H. Western and heart healthy dietary patterns differentially affect the expression of genes associated with lipid metabolism, interferon signaling and inflammation in the jejunum of Ossabaw pigs. J. Nutr. Biochem. 2021, 90, 108577. [Google Scholar] [CrossRef] [PubMed]
- Rippe, J.M.; Angelopoulos, T.J. Relationship between added sugars consumption and chronic disease risk factors: Current understanding. Nutrients 2016, 8, 697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aragno, M.; Mastrocola, R. Dietary sugars and endogenous formation of advanced glycation endproducts: Emerging mechanisms of disease. Nutrients 2017, 9, 385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masterjohn, C.; Park, Y.; Lee, J.; Noh, S.K.; Koo, S.I.; Bruno, R.S. Dietary fructose feeding increases adipose methylglyoxal accumulation in rats in association with low expression and activity of glyoxalase-2. Nutrients 2013, 5, 3311–3328. [Google Scholar] [CrossRef] [Green Version]
- Mizukami, H.; Osonoi, S. Pathogenesis and molecular treatment strategies of diabetic neuropathy collateral glucose-utilizing pathways in diabetic polyneuropathy. Int. J. Mol. Sci. 2021, 22, 94. [Google Scholar] [CrossRef] [PubMed]
- Goudarzi, M.; Kalantari, H.; Rezaei, M. Glyoxal toxicity in isolated rat liver mitochondria. Hum. Exp. Toxicol. 2018, 37, 532–539. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, D.; Moran, G.; Estrada, A.; Pagliassotti, M.J. Fructose-induced stress signaling in the liver involves methylglyoxal. Nutr. Metab. 2013, 10, 32. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, U.; Nagarajan, D. All-Trans Retinoic Acid supplementation prevents cardiac fibrosis and cytokines induced by Methylglyoxal. Glycoconj. J. 2017, 34, 255–265. [Google Scholar] [CrossRef]
- de Oliveira, M.G.; de Medeiros, M.L.; Tavares, E.B.G.; Mónica, F.Z.; Antunes, E. Methylglyoxal, a Reactive Glucose Metabolite, Induces Bladder Overactivity in Addition to Inflammation in Mice. Front. Physiol. 2020, 11, 290. [Google Scholar] [CrossRef] [Green Version]
- Younis, N.; Sharma, R.; Soran, H.; Charlton-Menys, V.; Elseweidy, M.; Durrington, P.N. Glycation as an atherogenic modification of LDL. Curr. Opin. Lipidol. 2008, 19, 378–384. [Google Scholar] [CrossRef]
- Kim, Y.S.; Kim, J.; Kim, K.M.; Jung, D.H.; Choi, S.; Kim, C.S.; Kim, J.S. Myricetin inhibits advanced glycation end product (AGE)-induced migration of retinal pericytes through phosphorylation of ERK1/2, FAK-1, and paxillin in vitro and in vivo. Biochem. Pharmacol. 2015, 93, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Rezaeinezhad, A.; Eslami, P.; Mirmiranpour, H.; Ghomi, H. The effect of cold atmospheric plasma on diabetes-induced enzyme glycation, oxidative stress, and inflammation; in vitro and in vivo. Sci. Rep. 2019, 9, 19958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yusufoğlu, B.; Yaman, M.; Karakuş, E. Determination of the most potent precursors of advanced glycation end products in some high-sugar containing traditional foods using high-performance liquid chromatography. J. Food Process. Preserv. 2020, 44, e14708. [Google Scholar] [CrossRef]
- Bellia, C.; Zaninotto, M.; Cosma, C.; Agnello, L.; Bivona, G.; Marinova, M.; Lo Sasso, B.; Plebani, M.; Ciaccio, M. Clinical usefulness of Glycated Albumin in the diagnosis of diabetes: Results from an Italian study. Clin. Biochem. 2018, 54, 68–72. [Google Scholar] [CrossRef]
- Giglio, R.V.; Lo Sasso, B.; Agnello, L.; Bivona, G.; Maniscalco, R.; Ligi, D.; Mannello, F.; Ciaccio, M. Recent updates and advances in the use of glycated albumin for the diagnosis and monitoring of diabetes and renal, cerebro-and cardio-metabolic diseases. J. Clin. Med. 2020, 9, 3634. [Google Scholar] [CrossRef]
- Raghav, A.; Ahmad, J.; Alam, K.; Khan, A.U. New insights into non-enzymatic glycation of human serum albumin biopolymer: A study to unveil its impaired structure and function. Int. J. Biol. Macromol. 2017, 101, 84–99. [Google Scholar] [CrossRef]
- Borg, D.J.; Forbes, J.M. Targeting advanced glycation with pharmaceutical agents: Where are we now? Glycoconj. J. 2016, 33, 653–670. [Google Scholar] [CrossRef]
- Thornalley, P.J. Use of aminoguanidine (Pimagedine) to prevent the formation of advanced glycation endproducts. Arch. Biochem. Biophys. 2003, 149, 31–40. [Google Scholar] [CrossRef]
- Chen, X.Y.; Huang, I.M.; Hwang, L.S.; Ho, C.T.; Li, S.; Lo, C.Y. Anthocyanins in blackcurrant effectively prevent the formation of advanced glycation end products by trapping methylglyoxal. J. Funct. Foods 2014, 8, 259–268. [Google Scholar] [CrossRef]
- Kim, J.H.; Zhang, K.; Lee, J.; Gao, E.M.; Lee, Y.J.; Son, R.H.; Syed, A.S.; Kim, C.Y. Trapping of Methylglyoxal by Sieboldin from Malus baccata L. and Identification of Sieboldin-Methylglyoxal Adducts Forms. Nat. Prod. Sci. 2021, 27, 245–250. [Google Scholar] [CrossRef]
- Gao, X.; Ho, C.T.; Li, X.; Lin, X.; Zhang, Y.; Chen, Z.; Li, B. Phytochemicals, Anti-Inflammatory, Antiproliferative, and Methylglyoxal Trapping Properties of Zijuan Tea. J. Food Sci. 2018, 83, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.; Swart, A. Aspalathus linearis (Rooibos)-a functional food targeting cardiovascular disease. Food Funct. 2018, 9, 5041–5058. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Sudji, I.R.; Wang, E.; Joubert, E.; Van Wyk, B.E.; Wink, M. Ameliorative effect of aspalathin from rooibos (Aspalathus linearis) on acute oxidative stress in Caenorhabditis elegans. Phytomedicine 2013, 20, 380–386. [Google Scholar] [CrossRef]
- Son, M.J.; Minakawa, M.; Miura, Y.; Yagasaki, K. Aspalathin improves hyperglycemia and glucose intolerance in obese diabetic ob/ob mice. Eur. J. Nutr. 2013, 52, 1607–1619. [Google Scholar] [CrossRef] [PubMed]
- Marnewick, J.L.; Rautenbach, F.; Venter, I.; Neethling, H.; Blackhurst, D.M.; Wolmarans, P.; MacHaria, M. Effects of rooibos (Aspalathus linearis) on oxidative stress and biochemical parameters in adults at risk for cardiovascular disease. J. Ethnopharmacol. 2011, 133, 46–52. [Google Scholar] [CrossRef]
- Dludla, P.V.; Muller, C.J.F.; Joubert, E.; Louw, J.; Essop, M.F.; Gabuza, K.B.; Ghoor, S.; Huisamen, B.; Johnson, R. Aspalathin protects the heart against hyperglycemia-induced oxidative damage by up-regulating Nrf2 expression. Molecules 2017, 22, 129. [Google Scholar] [CrossRef]
- Suchal, K.; Malik, S.; Khan, S.I.; Malhotra, R.K.; Goyal, S.N.; Bhatia, J.; Kumari, S.; Ojha, S.; Arya, D.S. Protective effect of mangiferin on myocardial ischemia-reperfusion injury in streptozotocin-induced diabetic rats: Role of AGE-RAGE/MAPK pathways. Sci. Rep. 2017, 7, 42027. [Google Scholar] [CrossRef] [Green Version]
- Mazibuko-Mbeje, S.E.; Dludla, P.V.; Roux, C.; Johnson, R.; Ghoor, S.; Joubert, E.; Louw, J.; Opoku, A.R.; Muller, C.J.F. Aspalathin-enriched green rooibos extract reduces hepatic insulin resistance by modulating PI3K/AKT and AMPK pathways. Int. J. Mol. Sci. 2019, 20, 633. [Google Scholar] [CrossRef] [Green Version]
- Uličná, O.; Vančová, O.; Božek, P.; Čársky, J.; Šebeková, K.; Boor, P.; Nakano, M.; Greksák, M. Rooibos tea (Aspalathus linearis) partially prevents oxidative stress in streptozotocin-induced diabetic rats. Physiol. Res. 2006, 55, 157–164. [Google Scholar] [CrossRef]
- Moens, C.; Muller, C.J.F.; Bouwens, L. In vitro comparison of various antioxidants and flavonoids from Rooibos as beta cell protectants against lipotoxicity and oxidative stress-induced cell death. PLoS ONE 2022, 17, e0268551. [Google Scholar] [CrossRef]
- Shao, X.; Bai, N.; He, K.; Ho, C.T.; Yang, C.S.; Sang, S. Apple polyphenols, phloretin and phloridzin: New trapping agents of reactive dicarbonyl species. Chem. Res. Toxicol. 2008, 21, 2042–2050. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, H.H.; Ramasamy, R.; Schmidt, A.M. Advanced glycation end products: Building on the concept of the “common soil” in metabolic disease. Endocrinology 2020, 161, bqz006. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, M.; Yamagishi, S. Possible Involvement of Advanced Glycation End-Products (AGEs) in the Pathogenesis of Alzheimers Disease. Curr. Pharm. Des. 2008, 14, 973–978. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Liu, J.; Dong, L.; Wang, X.; Zhang, X. Novel advances in inhibiting advanced glycation end product formation using natural compounds. Biomed. Pharmacother. 2021, 140, 111750. [Google Scholar] [CrossRef]
- Cordell, G.A. Sixty challenges—A 2030 perspective on natural products and medicines security. Nat. Prod. Commun. 2017, 12, 1371–1379. [Google Scholar] [CrossRef] [Green Version]
- Ramlagan, P.; Rondeau, P.; Planesse, C.; Neergheen-Bhujun, V.S.; Bourdon, E.; Bahorun, T. Comparative suppressing effects of black and green teas on the formation of advanced glycation end products (AGEs) and AGE-induced oxidative stress. Food Funct. 2017, 8, 4194–4209. [Google Scholar] [CrossRef]
- Babu, P.V.A.; Sabitha, K.E.; Shyamaladevi, C.S. Therapeutic effect of green tea extract on oxidative stress in aorta and heart of streptozotocin diabetic rats. Chem. Biol. Interact. 2006, 162, 114–120. [Google Scholar] [CrossRef]
- Nakagawa, T.; Yokozawa, T.; Kim, Y.A.; Kang, K.S.; Tanaka, T. Activity of Wen-Pi-Tang, and purified constituents of Rhei Rhizoma and glycyrrhizae radix against glucose-mediated protein damage. Am. J. Chin. Med. 2005, 33, 817–829. [Google Scholar] [CrossRef]
- Perez Gutierrez, R.M. Inhibition of advanced glycation end-product formation by Origanum majorana L. in vitro and in streptozotocin-induced diabetic rats. Evid.-Based Complement. Altern. Med. 2012, 2012, 598638. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.Y.; Kim, K. Protein glycation inhibitory and antioxidative activities of some plant extracts in vitro. J. Agric. Food Chem. 2003, 51, 1586–1591. [Google Scholar] [CrossRef]
- Lee, C.C.; Lee, B.H.; Lai, Y.J. Antioxidation and antiglycation of Fagopyrum tataricum ethanol extract. J. Food Sci. Technol. 2015, 52, 1110–1116. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.; Zheng, Z.; Cheng, K.W.; Shan, F.; Ren, G.X.; Chen, F.; Wang, M. Inhibitory effect of mung bean extract and its constituents vitexin and isovitexin on the formation of advanced glycation endproducts. Food Chem. 2008, 106, 475–481. [Google Scholar] [CrossRef]
- Grzegorczyk-Karolak, I.; Gołab, K.; Gburek, J.; Wysokińska, H.; Matkowski, A. Inhibition of advanced glycation end-product formation and antioxidant activity by extracts and polyphenols from Scutellaria alpina L. and S. altissima L. Molecules 2016, 21, 739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zheng, T.; Sang, S.; Lv, L. Quercetin inhibits advanced glycation end product formation by trapping methylglyoxal and glyoxal. J. Agric. Food Chem. 2014, 62, 12152–12158. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.-n.; Choi, E.-h.; Shim, S.M. Inhibitory activities of kaempferol against methylglyoxal formation, intermediate of advanced glycation end products. Appl. Biol. Chem. 2017, 60, 57–62. [Google Scholar] [CrossRef]
- Bhuiyan, M.N.I.; Mitsuhashi, S.; Sigetomi, K.; Ubukata, M. Quercetin inhibits advanced glycation end product formation via chelating metal ions, trapping methylglyoxal, and trapping reactive oxygen species. Biosci. Biotechnol. Biochem. 2017, 81, 882–890. [Google Scholar] [CrossRef] [Green Version]
- Bernacka, K.; Bednarska, K.; Starzec, A.; Mazurek, S.; Fecka, I. Antioxidant and Antiglycation Effects of Cistus × incanus Water Infusion, Its Phenolic Components, and Respective Metabolites. Molecules 2022, 27, 2432. [Google Scholar] [CrossRef]
- Bednarska, K. Potential of Vasoprotectives to Inhibit Non-Enzymatic Protein Glycation, and Reactive Carbonyl and Oxygen Species Uptake. Int. J. Mol. Sci. 2021, 22, 10026. [Google Scholar] [CrossRef]
- Bednarska, K.; Kuś, P.; Fecka, I. Investigation of the phytochemical composition, antioxidant activity, and methylglyoxal trapping effect of Galega officinalis L. Herb in vitro. Molecules 2020, 25, 5810. [Google Scholar] [CrossRef]
- Stander, M.A.; Van Wyk, B.E.; Taylor, M.J.C.; Long, H.S. Analysis of Phenolic Compounds in Rooibos Tea (Aspalathus linearis) with a Comparison of Flavonoid-Based Compounds in Natural Populations of Plants from Different Regions. J. Agric. Food Chem. 2017, 65, 10270–10281. [Google Scholar] [CrossRef]
- Joubert, E.; Jolley, B.; Koch, I.S.; Muller, M.; Van der Rijst, M.; de Beer, D. Major production areas of rooibos (Aspalathus linearis) deliver herbal tea of similar phenolic and phenylpropenoic acid glucoside content. S. Afr. J. Bot. 2016, 103, 162–169. [Google Scholar] [CrossRef]
- Walters, N.A.; de Villiers, A.; Joubert, E.; de Beer, D. Improved HPLC method for rooibos phenolics targeting changes due to fermentation. J. Food Compos. Anal. 2017, 55, 20–29. [Google Scholar] [CrossRef]
- Iswaldi, I.; Arráez-Román, D.; Rodríguez-Medina, I.; Beltrán-Debón, R.; Joven, J.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Identification of phenolic compounds in aqueous and ethanolic rooibos extracts (Aspalathus linearis) by HPLC-ESI-MS (TOF/IT). Anal. Bioanal. Chem. 2011, 400, 3643–3654. [Google Scholar] [CrossRef] [PubMed]
- Gattuso, G.; Caristi, C.; Gargiulli, C.; Bellocco, E.; Toscano, G.; Leuzzi, U. Flavonoid glycosides in bergamot juice (Citrus bergamia Risso). J. Agric. Food Chem. 2006, 54, 3929–3935. [Google Scholar] [CrossRef]
- Stalmach, A.; Mullen, W.; Pecorari, M.; Serafini, M.; Crozier, A. Bioavailability of C-linked dihydrochalcone and flavanone glucosides in humans following ingestion of unfermented and fermented rooibos teas. J. Agric. Food Chem. 2009, 57, 7104–7111. [Google Scholar] [CrossRef]
- Krafczyk, N.; Heinrich, T.; Porzel, A.; Glomb, M.A. Oxidation of the dihydrochalcone aspalathin leads to dimerization. J. Agric. Food Chem. 2009, 57, 6838–6843. [Google Scholar] [CrossRef]
- Bolton, W.K.; Cattran, D.C.; Williams, M.E.; Adler, S.G.; Appel, G.B.; Cartwright, K.; Foiles, P.G.; Freedman, B.I.; Raskin, P.; Ratner, R.E.; et al. Randomized Trial of an Inhibitor of Formation of Advanced Glycation End Products in Diabetic Nephropathy. Am. J. Nephrol. 2004, 24, 32–40. [Google Scholar] [CrossRef]
- Kiho, T.; Kato, M.; Usui, S.; Hirano, K. Effect of buformin and metformin on formation of advanced glycation end products by methylglyoxal. Clin. Chim. Acta 2005, 358, 139–145. [Google Scholar] [CrossRef]
- Beisswenger, P.J.; Ruggiero-Lopez, D. Metformin inhibition of glycation processes. Diabetes Metab. 2003, 29, S95–S96. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, S.; Minami, Y.; Taniguchi, R.; Tanaka, R.; Miyake, H.; Mori, T.; Ueda, M.; Shibata, T. Evaluation of anti-glycation activities of phlorotannins in human and bovine serum albumin-methylglyoxal models. Nat. Prod. Commun. 2017, 12, 1793–1796. [Google Scholar] [CrossRef]
- Kamakura, R.; Son, M.J.; de Beer, D.; Joubert, E.; Miura, Y.; Yagasaki, K. Antidiabetic effect of green rooibos (Aspalathus linearis) extract in cultured cells and type 2 diabetic model KK-Ay mice. Cytotechnology 2015, 67, 699–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pringle, N.A.; Koekemoer, T.C.; Holzer, A.; Young, C.; Venables, L.; Van De Venter, M. Potential Therapeutic Benefits of Green and Fermented Rooibos (Aspalathus linearis) in Dermal Wound Healing. Planta Med. 2018, 84, 645–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinae, N.; Shimoi, K.; Masumori, S.; Harusawa, M.; Furugori, M. Suppression of the Formation of Advanced Glycosylation Products by Tea Extracts. ACS Publ. 1994, 547, 68–75. [Google Scholar] [CrossRef]
- Kim, J.M.; Jang, D.S.; Lee, Y.M.; Yoo, J.L.; Kim, Y.S.; Kim, J.H.; Kim, J.S. Aldose-reductase- and protein-glycation-inhibitory principles from the whole plant of Duchesnea chrysantha. Chem. Biodivers. 2008, 5, 352–356. [Google Scholar] [CrossRef]
- Drygalski, K.; Siewko, K.; Chomentowski, A.; Odrzygóźdź, C.; Zalewska, A.; Krȩtowski, A.; MacIejczyk, M. Phloroglucinol Strengthens the Antioxidant Barrier and Reduces Oxidative/Nitrosative Stress in Nonalcoholic Fatty Liver Disease (NAFLD). Oxid. Med. Cell. Longev. 2021, 2021, 8872702. [Google Scholar] [CrossRef]
- Liu, J.; Yang, Z.; Cheng, Y.; Wu, Q.; He, Y.; Li, Q.; Cao, X. Eriodictyol and naringenin inhibit the formation of AGEs: An in vitro and molecular interaction study. J. Mol. Recognit. 2020, 33, e2814. [Google Scholar] [CrossRef]
- Perrone, A.; Giovino, A.; Benny, J.; Martinelli, F. Advanced Glycation End Products (AGEs): Biochemistry, Signaling, Analytical Methods, and Epigenetic Effects. Oxid. Med. Cell. Longev. 2020, 2020, 3818196. [Google Scholar] [CrossRef] [Green Version]
- Nevin, C.; McNeil, L.; Ahmed, N.; Murgatroyd, C.; Brison, D.; Carroll, M. Investigating the Glycating Effects of Glucose, Glyoxal and Methylglyoxal on Human Sperm. Sci. Rep. 2018, 8, 9002. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.M.; Na, M.K.; Na, R.B.; Min, B.S.; Lee, H.K. Antioxidant activity of two phloroglucinol derivatives from Dryopteris crassirhizoma. Biol. Pharm. Bull. 2003, 26, 1354–1356. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Gu, L. Phlorotannins from brown algae (Fucus vesiculosus) inhibited the formation of advanced glycation endproducts by scavenging reactive carbonyls. J. Agric. Food Chem. 2012, 60, 1326–1334. [Google Scholar] [CrossRef]
- Shao, X.; Chen, H.; Zhu, Y.; Sedighi, R.; Ho, C.T.; Sang, S. Essential structural requirements and additive effects for flavonoids to scavenge methylglyoxal. J. Agric. Food Chem. 2014, 62, 3202–3210. [Google Scholar] [CrossRef] [PubMed]
- Mendes, V.; Vilaça, R.; De Freitas, V.; Ferreira, P.M.; Mateus, N.; Costa, V. Effect of myricetin, pyrogallol, and phloroglucinol on yeast resistance to oxidative stress. Oxid. Med. Cell. Longev. 2015, 2015, 782504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, M.; Song, X.; Pan, J.; Gong, D.; Zhang, G. Vitexin Inhibits Protein Glycation through Structural Protection, Methylglyoxal Trapping, and Alteration of Glycation Site. J. Agric. Food Chem. 2021, 69, 2462–2476. [Google Scholar] [CrossRef] [PubMed]
- Erlund, I. Review of the flavonoids quercetin, hesperetin, and naringenin. Dietary sources, bioactivities, bioavailability, and epidemiology. Nutr. Res. 2004, 24, 851–874. [Google Scholar] [CrossRef]
- Muller, C.J.F.; Joubert, E.; Chellan, N.; Miura, Y.; Yagasaki, K. New insights into the efficacy of aspalathin and other related phytochemicals in type 2 diabetes—A review. Int. J. Mol. Sci. 2022, 23, 356. [Google Scholar] [CrossRef]
- Kreuz, S.; Joubert, E.; Waldmann, K.H.; Ternes, W. Aspalathin, a flavonoid in Aspalathus linearis (rooibos), is absorbed by pig intestine as a C-glycoside. Nutr. Res. 2008, 28, 690–701. [Google Scholar] [CrossRef] [PubMed]
- Blaut, M.; Schoefer, L.; Braune, A. Transformation of flavonoids by intestinal microorganisms. Int. J. Vitam. Nutr. Res. 2003, 73, 79–87. [Google Scholar] [CrossRef]
- Labib, S.; Erb, A.; Kraus, M.; Wickert, T.; Richling, E. The pig caecum model: A suitable tool to study the intestinal metabolism of flavonoids. Mol. Nutr. Food Res. 2004, 48, 326–332. [Google Scholar] [CrossRef]
- Zhang, S.; Jiao, T.; Chen, Y.; Gao, N.; Zhang, L.; Jiang, M. Methylglyoxal induces systemic symptoms of irritable bowel syndrome. PLoS ONE 2014, 9, e105307. [Google Scholar] [CrossRef] [Green Version]
- van der Lugt, T.; Opperhuizen, A.; Bast, A.; Vrolijk, M.F. Dietary advanced glycation endproducts and the gastrointestinal tract. Nutrients 2020, 12, 2814. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Rt [min] | [M − H]− | MS/MS | Error [ppm] | Compound | Rooibos Type | Source |
|---|---|---|---|---|---|---|---|
| 1. | 8.47 | 255.0504 | 193, 165 | −0.31 | Pisidic acid | RR | [50] |
| 2. | 13.76 | 341.0879 | 179, 135 | 1.87 | 1-O-caffeoylglucose | RR | [50] |
| 3. | 14.75 | 325.0929 | 119 | 1.64 | (E)-PPAG | RR | [51] |
| 4. | 15.08 | 289.0713 | 203, 109 | 0.29 | Epicatechin | RR | [50] |
| 5. | 17.98 | 609.1450 | - | −0.93 | Luteolin-6,8-di-C-glucoside or galactoside | RR, GR | [52] |
| 6. | 18.12 | 449.1089 | 329, 193, 135 | 1.13 | Eriodictyol-C-glucoside 1 | RR, GR | [52] |
| 7. | 18.82 | 449.1092 | 329, 193, 135 | 1.80 | Eriodictyol-C-glucoside 2 | RR, GR | [52] |
| 8. | 18.89 | 449.1085 | 329, 193, 135 | 0.24 | Eriodictyol-C-glucoside 3 | RR, GR | [52] |
| 9. | 19.15 | 449.1074 | 329, 193, 135 | −1.98 | Eriodictyol-C-glucoside 4 | RR, GR | [52] |
| 10. | 19.40 | 579.1329 | 399, 369 | −3.62 | Luteolin-C-glucoside-C-arabinoside (carlinoside) | GR | [53] |
| 11. | 20.44 | 447.0937 | 357, 327 | 2.14 | Luteolin-C-glucoside 1 (isoorientin or orientin) | RR, GR | [52] |
| 12. | 20.62 | 447.0937 | 357, 327 | 2.14 | Luteolin-C-glucoside 2 (isoorientin or orientin) | RR, GR | [52] |
| 13. | 20.96 | 451.1254 | 331, 209 | 3.01 | Aspalathin | RR, GR | S |
| 14. | 21.80 | 431.0994 | 341, 311 | 3.65 | Apigenin-C-glucoside 1 (vitexin) | RR, GR | S |
| 15. | 21.97 | 609.1461 | 300/301 | 0.87 | Queretin-3-O-robinobioside (bioquercetin, bioquercitrin) | RR | [50] |
| 16. | 22.19 | 431.0988 | 341, 311 | 2.26 | Apigenin-C-glucoside 2 (isovitexin) | RR, GR | S |
| 17. | 22.29 | 609.1461 | 300/301 | 0.87 | Queretin-3-O-rutinoside (rutin) | RR | S |
| 18. | 22.41 | 463.0883 | 300/301, 271 | 1.39 | Quercetin-3-O-β-galactoside (hyperoside) | RR | S |
| 19. | 22.70 | 463.0878 | 300/301, 271, 255 | 0.31 | Quercetin-3-O-β-glucoside (isoquercitrin) | RR | S |
| 20. | 23.30 | 435.1293 | 345, 315 | 0.42 | Nothofagin | RR, GR | [53] |
| 21. | 24.87 | 451.1254 | 331, 209 | 3.01 | Aspalathin isomer | RR, GR | [52] |
| 22. | 25.34 | 461.1077 | 446, 298/299 283/284 | −1.49 | Methyl-luteolin-O-glucoside or galactoside (e.g., chrysoeriol-O-glucoside/ galactoside) | RR | [54] |
| 23. | 25.65 | 493.1343 | 361, 331, 209 | −0.6 | Acetylaspalathin | RR | [50] |
| 24. | 27.55 | 287.0565 | 151, 135 | 3.25 | Eriodictyol | GR | S |
| 25. | 29.13 | 285.0405 | 285 | 2.05 | Luteolin | GR | S |
| 26. | 32.82 | 299.0560 | 284 | 1.45 | Chrysoeriol | GR | S |
| Compound | GRE | RRE | ||||||
|---|---|---|---|---|---|---|---|---|
| mg/100 mL or 1 g | μM | mg/100 mL or 1 g | μM | |||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| DIHYDROCHALCONES: | 45.95 | 1.28 | 1019.42 | 28.34 | 2.93 | 0.12 | 65.16 | 2.69 |
| Aspalathin | 41.23 | 1.28 | 911.26 | 28.25 | 2.36 | 0.13 | 52.25 | 2.78 |
| Nothofagin 1 | 4.72 | 0.02 | 108.16 | 0.51 | 0.56 | 0.01 | 12.91 | 0.21 |
| FLAVANONES: | 1.86 | 0.22 | 41.39 | 4.93 | 2.49 | 0.10 | 55.27 | 2.29 |
| Eriodictyol-6-C-glucoside 2 | 1.07 | 0.14 | 23.85 | 3.04 | 1.55 | 0.10 | 34.36 | 2.31 |
| Eriodictyol-8-C-glucoside 2 | 0.79 | 0.17 | 17.54 | 3.85 | 0.94 | 0.03 | 20.91 | 0.57 |
| FLAVONES: | 13.26 | 0.14 | 301.87 | 3.31 | 7.30 | 0.35 | 184.04 | 4.53 |
| Luteolin-6-C-glucoside (isoorientin) 3 | 4.93 | 0.03 | 109.96 | 0.63 | 3.14 | 0.13 | 70.04 | 2.89 |
| Luteolin-8-C-glucoside (orientin) 4 | 4.09 | 0.07 | 91.30 | 1.64 | 2.35 | 0.08 | 52.52 | 1.72 |
| Apigenin-8-C-glucoside (vitexin) | 1.39 | 0.16 | 32.35 | 3.78 | 0.40 | 0.04 | 9.47 | 0.86 |
| Apigenin-6-C-glucoside (isovitexin) | 2.66 | 0.05 | 61.78 | 1.20 | 0.13 | 0.01 | 2.92 | 0.15 |
| Luteolin | 0.08 | 0.01 | 2.74 | 0.32 | 1.05 | 0.02 | 36.84 | 0.76 |
| Apigenin + chrysoeriol 5 | 0.10 | 0.01 | 3.74 | 0.24 | 0.33 | 0.05 | 12.25 | 1.96 |
| FLAVONOLS: | 4.44 | 0.19 | 80.44 | 3.16 | 0.91 | 0.04 | 15.73 | 0.59 |
| Queretin-3-O-robinobioside (bioquercetin, bioquercitrin) 6 | 2.98 | 0.20 | 48.95 | 3.23 | 0.82 | 0.03 | 13.44 | 0.50 |
| Quercetin-3-O-β-galactoside (hyperoside) | 0.72 | 0.04 | 15.23 | 0.76 | 0.06 | 0.01 | 1.36 | 0.23 |
| Quercetin-3-O-β-glucoside (isoquercitrin) | 0.75 | 0.02 | 16.25 | 0.43 | 0.04 | 0.00 | 0.94 | 0.04 |
| SUM OF FLAVONOIDS | 65.51 | 1.52 | 1443.13 | 33.64 | 13.63 | 0.53 | 320.20 | 7.92 |
| Compound | Source | Rt [min] | [M − H]− | Error [ppm] | MGO-Adduct/Precursor |
|---|---|---|---|---|---|
| Aspalathin | S | 20.99 | 451.1254 | 3.01 | Aspalathin |
| 19.85 | 523.1475 | 4.45 | mono-MGO-aspalathin a | ||
| 21.45 | 523.1473 | 4.07 | mono-MGO-aspalathin b | ||
| GRE | 20.99 | 451.1254 | 3.01 | Aspalathin | |
| 19.85 | 523.1475 | 4.45 | mono-MGO-aspalathin a | ||
| 21.45 | 523.1473 | 4.07 | mono-MGO-aspalathin b | ||
| 25.65 | 523.1469 | 3.31 | mono-MGO-aspalathin c | ||
| 26.93 | 523.1449 | −0.51 | mono-MGO-aspalathin d | ||
| Nothofagin | GRE | 23.33 | 435.1293 | 0.42 | Nothofagin |
| 21.95 | 507.1519 | 3.24 | mono-MGO-nothofagin a | ||
| 23.43 | 507.1512 | 1.86 | mono-MGO-nothofagin b | ||
| 28.05 | 507.1521 | 3.63 | mono-MGO-nothofagin c | ||
| Vitexin | S | 21.81 | 431.0994 | 3.65 | Vitexin |
| 20.55 | 503.1205 | 3.07 | mono-MGO-vitexin a | ||
| 20.86 | 503.1208 | 3.66 | mono-MGO-vitexin b | ||
| Isovitexin | S | 22.19 | 431.0988 | 2.26 | Isovitexin |
| 20.82 | 503.1191 | 0.29 | mono-MGO-isovitexin a | ||
| 21.50 | 503.1185 | −0.90 | mono-MGO-isovitexin b | ||
| Eriodictyol | S | 27.51 | 287.0565 | 3.25 | Eriodictyol |
| 21.50 | 359.0772 | 1.40 | mono-MGO-eriodictyol a | ||
| 21.57 | 359.0771 | 1.28 | mono-MGO-eriodictyol b | ||
| 22.46 | 359.0777 | 2.80 | mono-MGO-eriodictyol c | ||
| 24.67 | 359.0775 | 2.24 | mono-MGO-eriodictyol d | ||
| Phloretin | S | 31.59 | 273.0780 | 2.56 | Phloretin |
| 27.05 | 417.1197 | 2.73 | di-MGO-phloretin a | ||
| 27.46 | 417.1198 | 2.97 | di-MGO-phloretin b | ||
| 28.82 | 345.0993 | 1.07 | mono-MGO-phloretin | ||
| Phloroglucinol | S | 2.02 | 125.0245 | 4.79 | Phloroglucinol |
| 4.28 | 197.0549 | 2.02 | mono-MGO-phloroglucinol a | ||
| 8.43 | 269.0813 | −2.97 | di-MGO-phloroglucinol a | ||
| 14.56 | 341.1067 | −1.64 | tri-MGO-phloroglucinol a | ||
| 14.82 | 269.0814 | −2.60 | di-MGO-phloroglucinol b | ||
| 15.01 | 341.1069 | −1.05 | tri-MGO-phloroglucinol b | ||
| 16.71 | 269.0812 | −3.34 | di-MGO-phloroglucinol c | ||
| 17.32 | 197.0540 | −2.53 | mono-MGO-phloroglucinol b |
| Compound | Source | Rt [min] | [M − H]− | Error [ppm] | GO-Adduct/Precursor |
|---|---|---|---|---|---|
| Aspalathin | S | 20.99 | 451.1254 | 3.01 | Aspalathin |
| 18.68 | 509.1311 | 3.10 | mono-GO-aspalathin a | ||
| 19.81 | 509.1310 | 2.90 | mono-GO-aspalathin b | ||
| 24.86 | 509.1316 | 4.08 | mono-GO-aspalathin c | ||
| GRE | 20.99 | 451.1254 | 3.01 | Aspalathin | |
| 18.68 | 509.1314 | 3.69 | mono-GO-aspalathin a | ||
| 19.81 | 509.1320 | 4.07 | mono-GO-aspalathin b | ||
| 24.86 | 509.1318 | 4.47 | mono-GO-aspalathin c | ||
| Nothofagin | GRE | 23.33 | 435.1293 | 0.40 | Nothofagin |
| Vitexin | S | 21.81 | 431.0994 | 3.65 | Vitexin |
| 19.03 | 489.1033 | −0.01 | mono-GO-vitexin a | ||
| 19.19 | 489.1044 | 2.23 | mono-GO-vitexin b | ||
| Isovitexin | S | 22.19 | 431.0988 | 2.26 | Isovitexin |
| Eriodictyol | S | 27.51 | 287.0565 | 3.25 | Eriodictyol |
| 21.56 | 345.0813 | 0.73 | mono-GO-eriodictyol a | ||
| 22.13 | 345.0810 | −0.13 | mono-GO-eriodictyol b | ||
| Phloretin | S | 31.59 | 273.0780 | 2.56 | Phloretin |
| 26.88 | 331.0833 | 4.59 | mono-GO-phloretin | ||
| Phloroglucinol | S | 2.02 | 125.0245 | 4.79 | Phloroglucinol |
| 8.15 | 241.0482 | 2.9 | di-GO-phloroglucinol a | ||
| 13.79 | 183.0366 | 1.36 | mono-GO-phloroglucinol a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bednarska, K.; Fecka, I. Aspalathin and Other Rooibos Flavonoids Trapped α-Dicarbonyls and Inhibited Formation of Advanced Glycation End Products In Vitro. Int. J. Mol. Sci. 2022, 23, 14738. https://doi.org/10.3390/ijms232314738
Bednarska K, Fecka I. Aspalathin and Other Rooibos Flavonoids Trapped α-Dicarbonyls and Inhibited Formation of Advanced Glycation End Products In Vitro. International Journal of Molecular Sciences. 2022; 23(23):14738. https://doi.org/10.3390/ijms232314738
Chicago/Turabian StyleBednarska, Katarzyna, and Izabela Fecka. 2022. "Aspalathin and Other Rooibos Flavonoids Trapped α-Dicarbonyls and Inhibited Formation of Advanced Glycation End Products In Vitro" International Journal of Molecular Sciences 23, no. 23: 14738. https://doi.org/10.3390/ijms232314738