1. Introduction
Vaginal infections, including vulvovaginal candidiasis (VVC), have gained increased relevance in everyday medical practice, which is also reflected in the scientific literature. Being the second most commonly diagnosed gynecological infection, VVC affects over 75% of women of reproductive age worldwide and is often associated with therapeutic failures and the risk of recurrence. The most common and relatively easily recognized etiological factors of this type of infection are yeast-like fungi of the genus
Candida. Physicians and scientists have been paying more and more attention to mucosal candidiasis—including vulvovaginal candidiasis. Since it is not an infection that requires hospitalization, and its symptoms, as well as possible complications and relapses, are classified as relatively mild, it can be easily underestimated. In recent years, there has been a growing interest in VVC among clinicians and scientists, as well as among patients themselves [
1,
2,
3,
4,
5,
6]. However, despite the increasing amount of research conducted on the pathogenesis of VVC and its relapsed form, as well as on effective treatment, it turns out that more remains unknown than known. Given the difficulties associated with establishing the detailed mechanism of the development of fungal infection on the vaginal mucosa, it becomes virtually impossible to identify a specific factor as a target for potential new antimicrobials [
6,
7,
8,
9,
10]. It is, therefore, not surprising that, on the one hand, intensive research is being carried out to identify the specific pathogenesis of these infections (which would facilitate the fight against and prevention of these infections [
2,
3,
7]), and on the other hand, many research studies have focused on the search for new substances or therapeutic regimens with potential antifungal activity in the vagina. Because it has been known for years that
Candida biofilm plays a role (if not the key role) in treatment resistance and VVC relapses, the newly synthesized agents are also tested for anti-biofilm activity [
2,
3,
7,
8,
11,
12,
13].
Short cationic compounds are probably the largest group among the compounds that can be an alternative to mycobiotics conventionally used against yeast-like fungi. Among them, ultrashort cationic lipopeptides (USCLs) deserve special attention, as they are especially active against fungi. Their structure consists of short positively charged peptides (seven amino acid residues at most), mainly due to lysine (K) or arginine (R) residues, linked to a fatty acid chain [
14,
15,
16,
17]. As a result, these compounds are amphiphilic and easily react with the negatively charged components of the
Candida cell membrane—sialic acid and phosphatidylinositol [
18,
19]. All of this results in the permeabilization of these membranes and, in effect, cell death [
15,
16,
17,
20]. In addition to antifungal activity, USCLs also show many other properties as antimicrobial and anti-adhesive activity, translating to their potent anti-biofilm effect [
21].
In recent years, commonly used laboratory methods to determine the effect of such compounds have been considered a significant obstacle in the research on new antifungal and antibiofilm agents. Researchers often point out that in vitro methods, which primarily determine the MIC (minimum inhibitory concentration) and MBEC (minimum biofilm eradication concentration) values, may turn out to be inadequate in relation to the actual in vivo activity of the substance. Hence, various models are increasingly used, including in vivo and ex vivo models, which are mainly animal (mouse and rat) models [
12,
22]. So far, the uniqueness of the vaginal microenvironment and the crucial importance of the as yet unexplained (but definitely existing) interactions between
Candida and vaginal epithelium cells (VECs) have been proven. While there are already relatively well-described models of mucosal candidiasis in the oral cavity or the gastrointestinal tract in research, a reliable and universal research model for VVC remains a challenge [
12,
22]. In 2010, Harriot et al. were the first to obtain
Candida biofilm on the surface of vaginal tissue in in vivo and ex vivo animal models. Their comparison showed that in both types of models, the kinetics of biofilm formation and its structure are the same and could be used in further research, as well as that they most likely correspond to biofilms obtained in vitro, e.g., on silicone disks [
12]. In 2016, Krom and Willems also compared other methods of testing
Candida biofilm and its drug susceptibility in vitro, indicating an interesting method for studying this structure under dynamic and realistic microfluidic conditions using the BioFlux-based system [
23]. The BioFlux system has many advantages, including the most important two: observing the behavior of microorganisms (biofilms) under microfluidic conditions, thanks to the performance of all experiments in the microcapillaries of this system and the possibility of real-time observation of the processes taking place. The study of fungal biofilm in the flow may have some practical impact depending on the site of infection—it is obvious that this model will be much more useful for imitating conditions prevailing in, for example, blood vessels than in mucosal infections. Referring to the strains isolated from VVC, BioFlux enables continuous 24-hour observation of the kinetics of biofilm development, including
Candida adhesion and germination, the production of germ tubes and filaments, as well as the subsequent detachment of fragments of the mature structure and its “migration” further. The advantage of BioFlux over standard in vitro methods in drug susceptibility testing is manifested in the assessment of the results—the possibility of observing and assessing the formation of microbial aggregates, their developmental form, and increasing or decreasing coverage of microcapillaries—without the need to take into account potential errors and problems resulting, for example, from manual handling of microscope slides or rinsing of the biofilm. Apart from the evaluation of dynamic processes, BioFlux also enables molecular mechanisms, and in the adjacent microcapillaries, one can simultaneously conduct many other experiments [
23,
24,
25].
In our previous work, we compared the antifungal activity of two pairs of cationic lipopeptides against
Candida strains isolated from vulvovaginal candidiasis. These compounds effectively eradicated the mature structure of the biofilm [
26]. In in vitro studies using polystyrene plates, we have demonstrated the advantage of the newly synthesized cyclic analogs C
16-CKKKKC-NH
2 and C
16-CKRKKC-NH
2 over their linear counterparts in the antimicrobial activity against
Candida in planktonic and biofilm form [
26]. At the same time, we have already proven that the use of combination therapy in the form of the simultaneous action of antifungal fluconazole that is conventionally used in VVC and the tested lipopeptides could be equally promising—this time indicating the advantage of the linear compounds C
16-KKKK-NH
2 and C
16-KRKK-NH
2 [
26]. As we have mentioned before, such an approach could solve the problem of the relatively high toxicity of this type of compound in relation to eukaryotic cells, because the combination of USCLs with fluconazole turned out to be effective at concentrations several times lower than when these substances were used separately [
26].
The aim of the study was to verify the results obtained previously in classic in vitro tests using the ex vivo VVC mouse model, and in the in vitro biofilm model under microfluidic conditions with the BioFlux-based system. The most promising pair of lipopeptides selected on the basis of previous work were used: linear C16-KKKK-NH2 and cyclic C16-CKKKKC-NH2. Their effect on mature fungal biofilm was tested against two selected clinical strains of Candida albicans isolated from VVC, for which the drug susceptibility of the biofilm was previously determined. Using effective in vitro concentrations, the activity of the tested lipopeptides was verified both directly on the mouse vaginal tissue (ex vivo model) and under microfluidic conditions (BioFlux model).
3. Discussion
Lipopeptides are a group of substances with a wide range of antimicrobial activity already known to researchers. In our previous work, using classical in vitro methods (determination of MIC, MBEC, and FIC values), we proved the effectiveness of the newly synthesized ultra-short cyclic lipopeptides (USCLs) against yeast-like fungi, both in planktonic and biofilm form [
15,
26]. Among other properties, USCLs have recognized anti-adhesive properties and, thus, anti-biofilm and antimicrobial action, including against
Candida [
14,
15,
16,
17]. For this reason, we are investigating the effect of these compounds on
Candida strains isolated from vulvovaginal candidiasis (VVC). Both in the scientific literature and in everyday clinical practice, attention is increasingly being paid to this infection for several reasons, including its extremely common occurrence, the still unknown etiology, and the increasingly frequent and difficult to explain treatment failures and recurrences of VVC. Based on our previous research, two (linear and cyclic) of the most promising USCLs were selected for further study. Their structure consists of a palmitic acid residue (C16, hexadecenoic acid) conjugated to a positively charged peptide with L-lysine residues and C-terminal amide. In the case of the cyclic analog, cyclization was achieved with two cysteine (C-cysteine) residues linked by a disulfide bridge [
16,
19]. In our study, the compounds C
16-KKKK-NH
2 (L1) and C
16-CKKKKC-NH
2 (C1) were able to inhibit the plankton growth of various
Candida strains isolated from VVC, as well as to eradicate the mature biofilm structure formed by these isolates at lower concentrations than the other tested USCLs [
26]. At the same time, they showed relatively low toxicity towards HaCaT and the lowest hemolytic capacity [
15]. In addition, they showed the most favorable (synergistic or additive) effect when using combination therapy—the simultaneous use of these lipopeptides in combination with antifungal fluconazole, which is conventionally used in VVC [
26].
The aim of this study was to verify the results obtained previously using standard in vitro methods for determining drug susceptibility. This was performed using two other methods of testing the effect of various compounds on
Candida biofilm—a newly developed ex vivo model, in which we used mouse vaginal tissues, and a biofilm model in microfluidic conditions, obtained by using BioFlux technology. Two
C. albicans strains isolated from VVC were selected for the study, with MIC, MBEC, and FIC values previously determined for fluconazole as well as for L1 and C1 lipopeptides [
26]. The experiments consisted of creating a mature (i.e., 24-h) biofilm using these strains, and then treating this structure with the tested compounds at concentrations considered in the course of previous studies to be the lowest which were effective in eradication. The expected result was at least a notable reduction in biofilm biomass under the influence of L1 and C1 (at concentrations equal to MBEC) and their combination with fluconazole (at concentrations equal to FIC), similarly to the RPMI 1640 microdilution method on polystyrene plates. In addition, it was decided to use sub-inhibitory concentrations of lipopeptides in the model using mouse vaginal tissues. There exist reports on the potential action of certain AMPs (antimicrobial peptides), including cationic lipopeptides, resulting in sensitization of various microorganisms to the effects of the so-called host defense peptides (HDPs) which naturally occur in the body. Although the mechanism of this action is currently unknown, it has already been observed that the addition of low (sub-inhibitory) concentrations of AMPs to, for example, serum may inhibit bacterial growth, most likely enhancing the action of complement proteins, lysozyme, or lactoferrin [
16,
27,
28]. It is known that there is a mucus layer on the vaginal mucosa and in the vagina itself, produced by the VEC, which has antimicrobial properties and contains many substances, including lactoferrin and lysozyme [
29,
30]. Unfortunately, in vitro, it is almost impossible, or at least extremely difficult, to recreate or obtain this substance in a manner similar to, for example, the collection of saliva or serum. In addition, obtaining a tissue culture of VK2 cells would not reflect the complex mechanisms between VECs, their products, or yeast-like fungi. Hence, it was decided that we would test the hypothesis of the potential supporting action of lipopeptides at sub-inhibitory concentrations against various, including unidentified, HDPs in the vaginal microenvironment using an ex vivo animal model.
The MBECs were identical for both tested
C. albicans. In the in vitro microdilution method in RPMI 1640 on polystyrene plates, the use of these concentrations resulted in no signal (MTT solution color change) when visualizing the results, which meant that no metabolically active sessile cells were present [
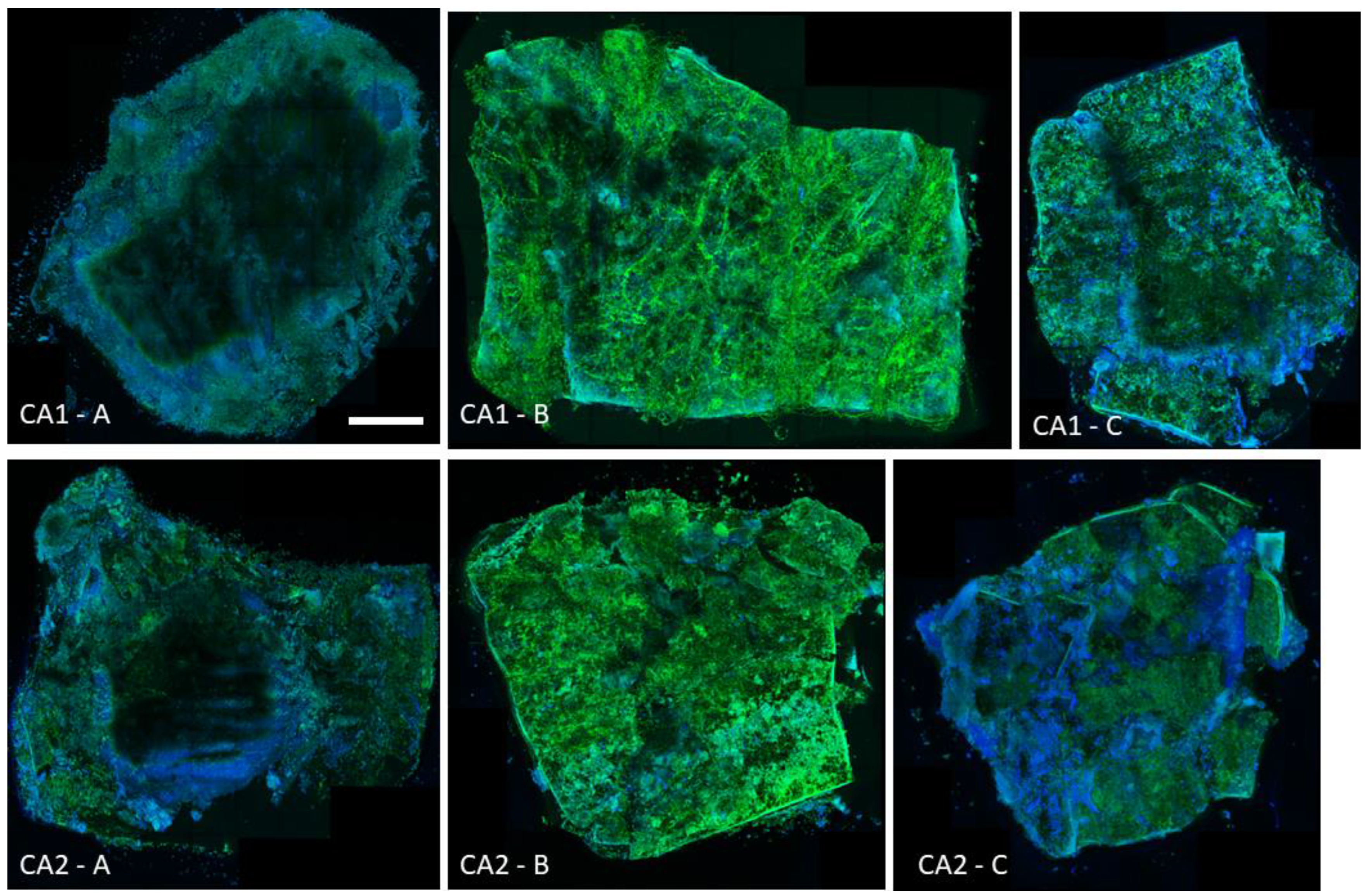
26]. In an ex vivo animal model using mouse vaginal tissue fragments, treatment of the 24 h biofilm formed by these strains with the same concentrations of L1 and C1 did not eradicate
Candida. In confocal microscopy, a thick layer of fungal hyphae stained with Calcofluor White was still visible. The failure of biofilm eradication in this model was demonstrated by the results obtained with tissue homogenization as well. For both the CA1 and CA2 strains, in the presence of L1 equal to MBEC, the mean values of CFU/g of tissue were generally higher than in positive controls—
Figure 2 and
Figures S1 and S3 in Supplementary Materials; no statistical significance (
p > 0.05). For strain CA1, when using C1, the value of CFU/g visibly exceeded the result for the positive control. A low decrease in mean value was only obtained for CA2 using C1 (again, without statistical significance). However, it should be kept in mind that the obtained results may be subject to some variations depending on various laboratory factors. Examples of such steps potentially leading to greater uncertainty include manual homogenization of tissue fragments or transfer of tissues with strain suspensions to microscope slides in CM. Furthermore, one should remember the tissue-dependent characteristics, as the tissues were obtained from successive mice. Therefore, the noticeable standard deviations shown in
Figure 2 should not be surprising either. Nevertheless, the presence of a relatively large amount of
Candida (most often exceeding the values obtained in positive controls) is certain, despite the use of eradicating lipopeptide concentrations. In no case was there a decrease in the amount of fungi on the surface of the tissue, which was comparable, for example, to the significantly lower values obtained for high concentrations of amphotericin B (
Figure 2,
Figures S1–S4 in Supplementary Materials) (
p < 0.05). On this basis, it can be concluded that the results obtained with the classical in vitro methods were not confirmed in the ex vivo method. Of course, the question remains as to which of the methods is more reliable. Microdilution methods have been recognized and approved for many years as reliable by all major institutions and scientific and clinical societies; they are repeatable and verifiable. Their translation into the clinical use of the tested compounds is also confirmed and effective [
31,
32]. However, there are some clinical exceptions, and we believe that vulvovaginal candidiasis should be considered such an exception. This is because, despite establishing the drug susceptibility of
Candida isolates in VVC therapy, failures still occur [
2,
3,
6,
8]. The discussion on what we actually know about the biofilm formed by microorganisms and what the structure which we clinically consider biofilm, and above all, in vitro, really is, has been going on for several years [
33]. When considering the interpretation of the results of our work, these issues are slightly less important—it does not seem to matter whether the 24-h structure formed by the suspension of
C. albicans plankton cells on the surface of the tissue is called biofilm. In the ex vivo model, we have the opportunity to observe how yeast-like fungi behave towards the vaginal epithelium, as well as how various compounds, including the USCLs tested, may act in this environment. In 2010, Harriot et al. conducted a comparative analysis of the
Candida biofilm itself, and of the kinetics of its formation, in the in vivo, ex vivo model as well as on polystyrene plates in RPMI 1640. They concluded that the dynamics of biofilm formation and its individual stages, as well as the structure and properties of mature (24-h) biofilm, are comparable and almost identical [
12]. If so, the reason for the differences in the results of biofilm eradication by L1 and C1 lipopeptides obtained with these methods should be seen in the interaction of fungi with the abiotic and biotic surfaces. On the surface of the vaginal tissue,
Candida undoubtedly interact with epithelial cells, deriving primarily nutrients from them (no nutrient medium for fungi is used in the ex vivo model) [
34]. According to the current knowledge,
C. albicans has the ability to actively invade tissues thanks to many virulence factors, including filamentation. Thus, the hyphae observed on the epithelium should be considered an invasive form of these strains, displaying, at the same time, tissue-destructive activity [
29,
35]. It is also known that the proliferating and filamentous
Candida blastospores also have the ability to produce a large amount of extracellular substance, or mucus (matrix–ECM in accordance with the dynamics of biofilm formation), which is a barrier impermeable to various substances [
6,
8,
12]. On the surface of a polystyrene plate, an abiotic material, yeast-like fungi also multiply intensively and have the ability to germinate and produce mucus. However, this method provides a nutrient medium—RPMI 1640—so these isolates do not have to compete for substances, and have no possibility of further invasion. Perhaps this is the reason why it is “easier” to eradicate yeast-like fungus overnight on a given surface (which we call mature biofilm) in an in vitro model than in an ex vivo tissue model. If so, the method that uses the animal equivalent of the vaginal epithelium, which undoubtedly more closely imitates the real processes of VVC, seems to be much more reliable than the polystyrene plates. If, in the course of further studies, the described observations and results are confirmed, it could indicate the need to drastically change the methods of
Candida susceptibility testing used in microbiological diagnostics towards VVC.
The use of concentrations corresponding to the lowest FIC also did not result in complete eradication of
C. albicans biofilm for either tested strain. However, the use of FIC resulted in the lowest mean number of CFU/g of tissue for the entire experiment (no statistical significance,
p > 0.05). A decrease can be seen for CA1 and for CA2, for both lipopeptide–fluconazole combinations (
Figure 2,
Figures S1–S4 in Supplementary Materials). It is worth emphasizing that the fact that the classic checkerboard method used by our team in the previous work to determine the FIC and FIC indexes refers to plankton cells and not to biofilm [
26] speaks in favor of the presented results. Thus, naturally, one would expect higher MICs of both substances in combination during eradication of this highly resistant structure than against
Candida plankton. Meanwhile, concentrations many times lower than MBEC (and MIC) turned out to be partially effective in our ex vivo method. A comparison of the results obtained in our work (
Table 1) suggests that the simultaneous use of linear USCL concentrations 16 times (isolate 1) and 32 times (isolate 2) lower (than MBEC), as well as the use of concentrations of cyclic USCL 32 times (both isolates) lower (than MBEC), with the addition of fluconazole at a concentration 64 times lower than the MIC, produces similar or even better results in eradication of
C. albicans biofilm from the surface of the murine vaginal epithelium [
26]. The above may also prove that the combination of substances with different mechanisms of action can actually be more effective, which has been postulated for a long time [
36,
37,
38,
39]. As we mentioned in our previous work, the most likely explanation of the activity of these compounds against
C. albicans is the possibility of sensitizing fungal cells by means of USCLs, which is conducted by causing at least partial damage to cell membranes, enabling faster and more effective interaction of fluconazole with its molecular target inside the cell (14 α-ergosterol demetylase). At the same time, the very interaction of this azole with the cell membrane may increase its permeabilization by lipopeptides [
26]. Proving the effectiveness of using a combination of lipopeptides with fluconazole at concentrations many times lower than, when these substances are used separately, could solve the two most serious problems that should be taken into account in research on antimicrobial activity. The use of lipopeptides at much lower concentrations could no longer be associated with the potential relatively high toxicity of these compounds, which is increasingly noted. In addition, by treating cells with substances with different mechanisms of action, the acquisition of resistance by microorganisms, including
Candida, would be significantly hampered, especially since the acquisition of resistance to AMPs, including USCLs, is already considered unlikely [
16]. Thus, the results obtained in the course of the presented studies on the eradication of
C. albicans biofilm from the tissue, using a combination of low concentrations of fluconazole and both lipopeptide analogs, are highly promising and require further confirmation.
It seems that the most diverse and difficult to interpret results of tissue biofilm eradication were obtained using sub-inhibitory concentrations (1/2 MBEC). Results were, again, statistically insignificant (
p > 0.05). With the use of L1, some reduction in mean CFU/g values was obtained for both
C. albicans strains (
Figure 2,
Figures S1 and S3 in Supplementary Materials). Regarding C1, one value indicates an eradicating effect—a decrease in mean CFU/g for CA2. For CA1, the obtained values exceeded those determined for the positive controls. It can, therefore, be concluded that the sub-inhibitory MBEC concentrations of the linear lipopeptide have a similar eradicating capacity to the use of the combination of this analog with fluconazole. This would be in contrast to the cyclic USCL, which is actually ineffective at one-half of the MBEC concentrations. At present, it is difficult to determine why a more favorable effect of linear analogs at low concentrations could be observed, since when using the lipopeptide alone, cyclic compounds turn out to be much more effective against
Candida (and their MIC values are much lower). It is worth noting, however, that both lipopeptides showed synergistic or additive activity with fluconazole, and their combinations were comparably effective in eradicating fungal biofilm from the tissue. It is likely that, in some way, a linear analog at low concentrations sensitizes
Candida cells to the effects of fluconazole when used in combination therewith, and also analogously enhances the antifungal activity of HDPs and other immune mechanisms present in the vaginal tissue [
16,
27,
28]. If the cyclic compound acts similarly in combination with fluconazole, and if it is indeed possible, for example, to synergize with HDPs, why does C1 not have this ability when L1 does? Any potential explanation must take into account the differences in the structure of both compounds, as well as their interactions with VECs and the cells of the immune system, and with the substances they produce. Even if the aforementioned hypothesis regarding supporting the action of HDPs and/or other immunological components is true, its confirmation requires many verification studies, focused primarily on determining the nature of the interaction of all the substances and components present in the vagina—especially in the presence of
Candida fungi.
Under the microfluidic conditions obtained using the BioFlux technology, it was possible to obtain a 24-h structure formed by blastospores and hyphae of both tested
C. albicans strains—which we call biofilm. The microscopic photos of the channels used in BioFlux, taken every hour, clearly showed the next stages of the development of fungal biofilm—especially visible in
Videos S1 and S2 animations and in the example in
Figure 4. These observations are consistent with the common knowledge on the kinetics of biofilm formation, and (not for the first time) they confirm the equal suitability of the microflow model and in vitro models, as well as of the in vivo and ex vivo animal models, already proven by Harriot et al. [
6,
13]. BioFlux is currently considered to be another method of testing the biofilm of microorganisms and its drug susceptibility in vitro. Thanks to the use of BioFlux technology, we were able to observe the behavior of the strains in real time. We managed both to confirm the occurrence of all the commonly described stages of fungal biofilm formation, and to visually and quantitatively verify the effect (or lack thereof) of the tested substances on the investigated
C. albicans isolates. According to the assumptions, if, for example, yeast-like fungi behave similarly/almost identically on polystyrene plates and in BioFlux channels, they should also show at least similar drug sensitivity when exposed to the same eradicating substances [
23]. Following these assumptions, we treated the 24-h structure formed by
C. albicans strains with L1 and C1 lipopeptides at concentrations equal to MBEC—again, expecting to confirm the effectiveness of these compounds in combating biofilm in vitro. Meanwhile, the results obtained by us do not prove the eradication abilities of USCLs, but the slight (and statistically insignificant) possibilities of limiting the further development of fungal biofilm. In
Figure 5 and
Figure 6 for strains CA1 and CA2, respectively, we can observe lower percentages of increasing coverage of channels in which biofilm was previously formed. Compared to the positive controls, especially for CA2, a relatively markedly lower further increase in the amount of
Candida on the surface of the channels can be seen. During the first 8 h of incubation with the L1 flow, there was almost no further multiplication of the fungi (value 0.29 vs. 2.45% in K(+)). Ultimately, after one day, the percentage of coverage increase was nearly 50 percent lower than in K(+) (4.35 vs. 8.60%). The analogous results for CA1 also show an insignificant reduction in multiplication only in the first 8 h of incubation with L1 (2.21 vs. 2.64% in K(+)).After 24 h, the final value is lower by almost ¼ (6.88 vs. 9.39% in K(+)). In both cases, compound C1 limited biofilm development of both strains in the channels to a much lesser extent—the final values for CA1 were 8.40% (vs 9.39%), and for CA2, 6.18% (vs 8.60%). In both cases, after 12 h,
C. albicans formed a very thick layer of biofilm in the channels. This physically, above all, significantly hindered the further flow of any substances. Given the fungal ability to produce a large amount of highly impermeable matrices (ECM), also most likely abundantly present in these cases, the relatively poor penetration of L1 and C1 is not surprising [
8,
33,
40]. It should also come as no surprise that action in the first hours was more effective. It is likely that during the first few flows of substances, there was the greatest mechanical effect on the hyphae present in the channels and on their detachment. These, at the same time, could expose the more sensitive cells located in the deeper layers of the biofilm to the effects of USCLs, limiting their multiplication or even their microbiotic effect (percentages are presented for the entire length of the channels observed under the microscope). The microfluidic model is, perhaps, not the best reflection of
Candida kinetics and of the interactions with test substances during VVC. However, comparing the results obtained with the use of BioFlux and with polystyrene plates, it can be concluded that for the effective action of lipopeptides against fungal biofilm, constant contact of microorganism cells with antimicrobial substances is necessary, as is the case with polystyrene plates. However, in both models, it is difficult to detect active invasion of
C. albicans during adhesion to abiotic surfaces, so this in vitro model does not seem to be appropriate for drawing conclusions regarding the behavior of yeast-like fungi on the surface of the vaginal tissue during infection.


 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}