
Gma-miR408 Enhances Soybean Cyst Nematode Susceptibility by Suppressing Reactive Oxygen Species Accumulation
 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Results
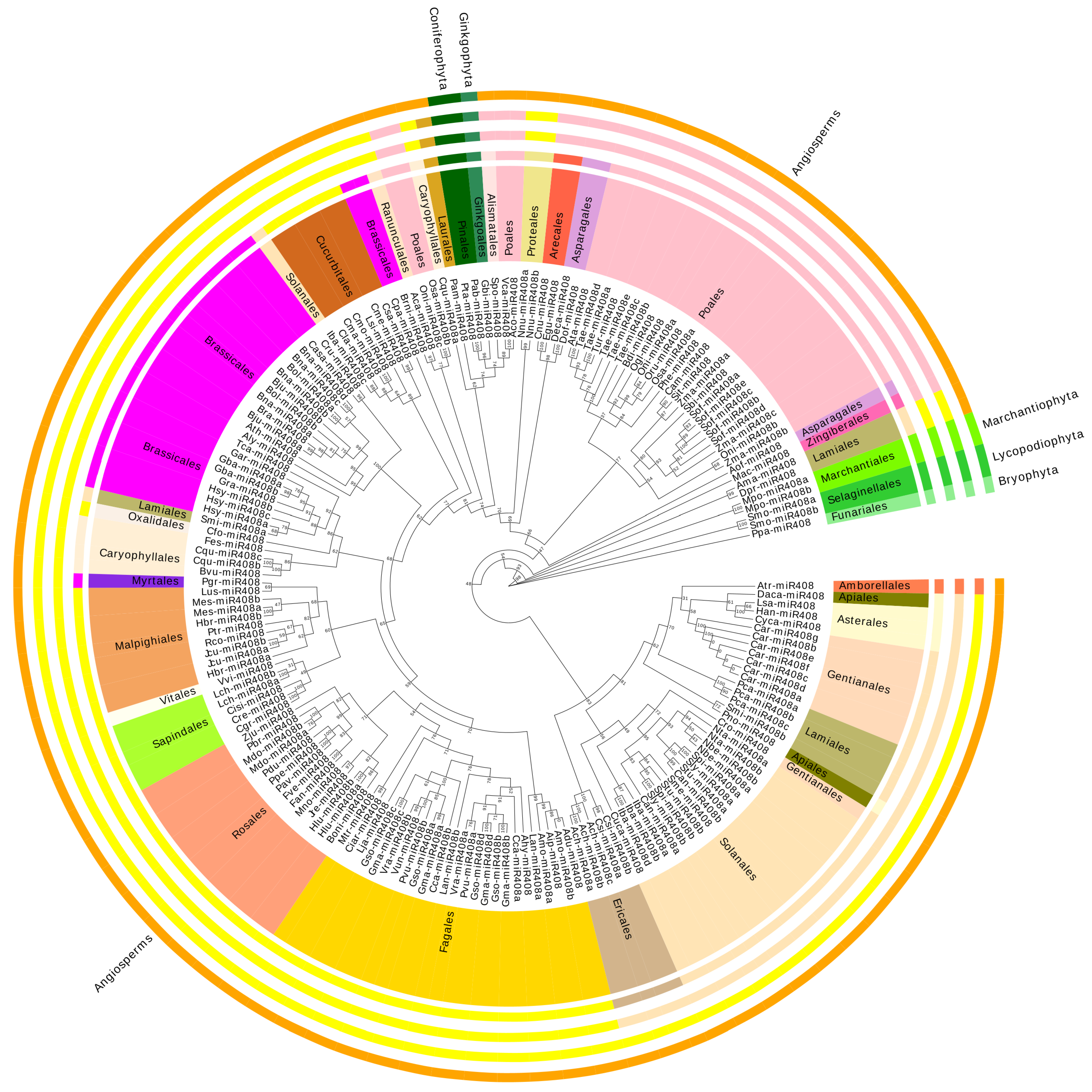
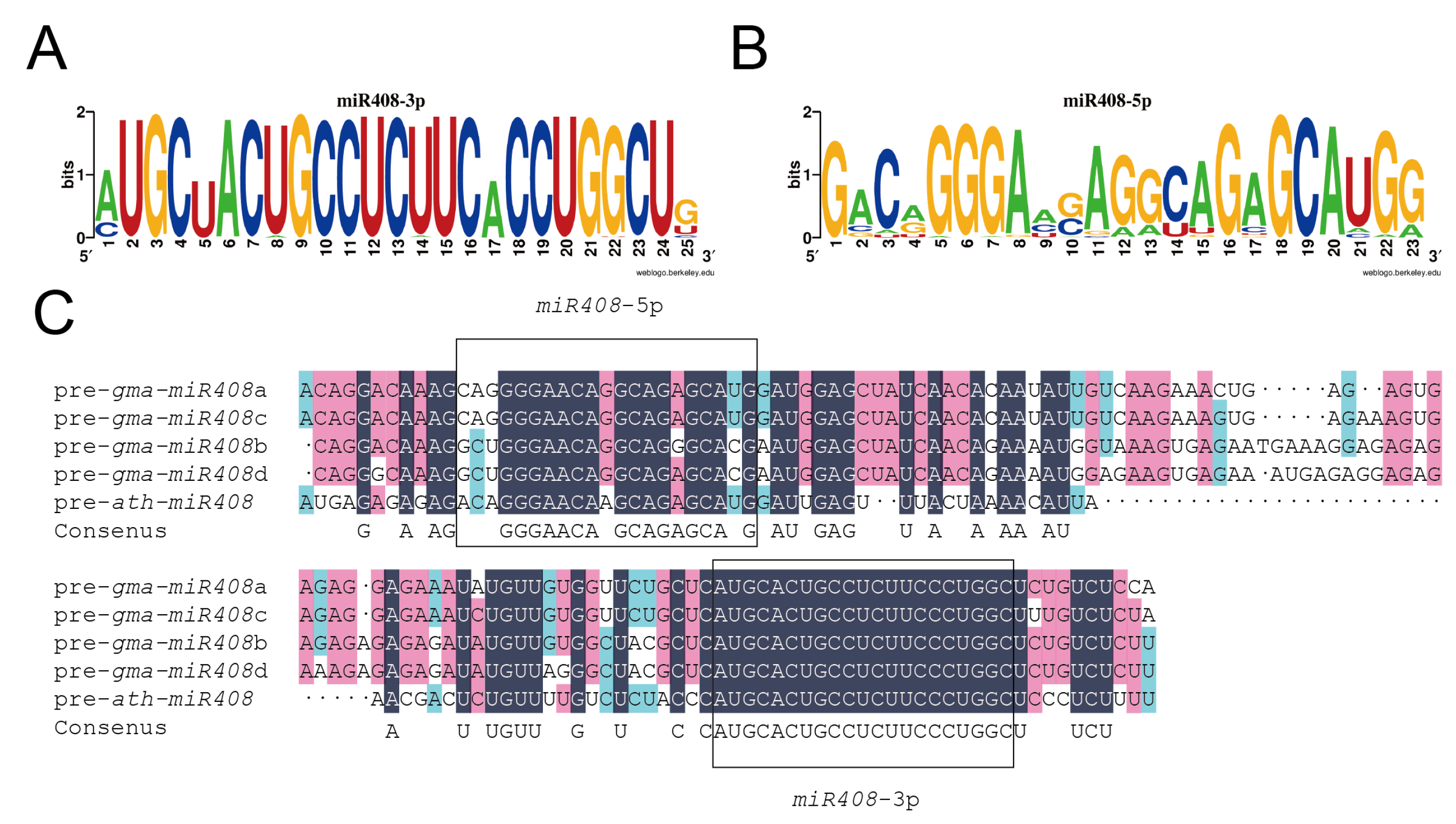
2.1. Identification of miR408 in Glycine Max
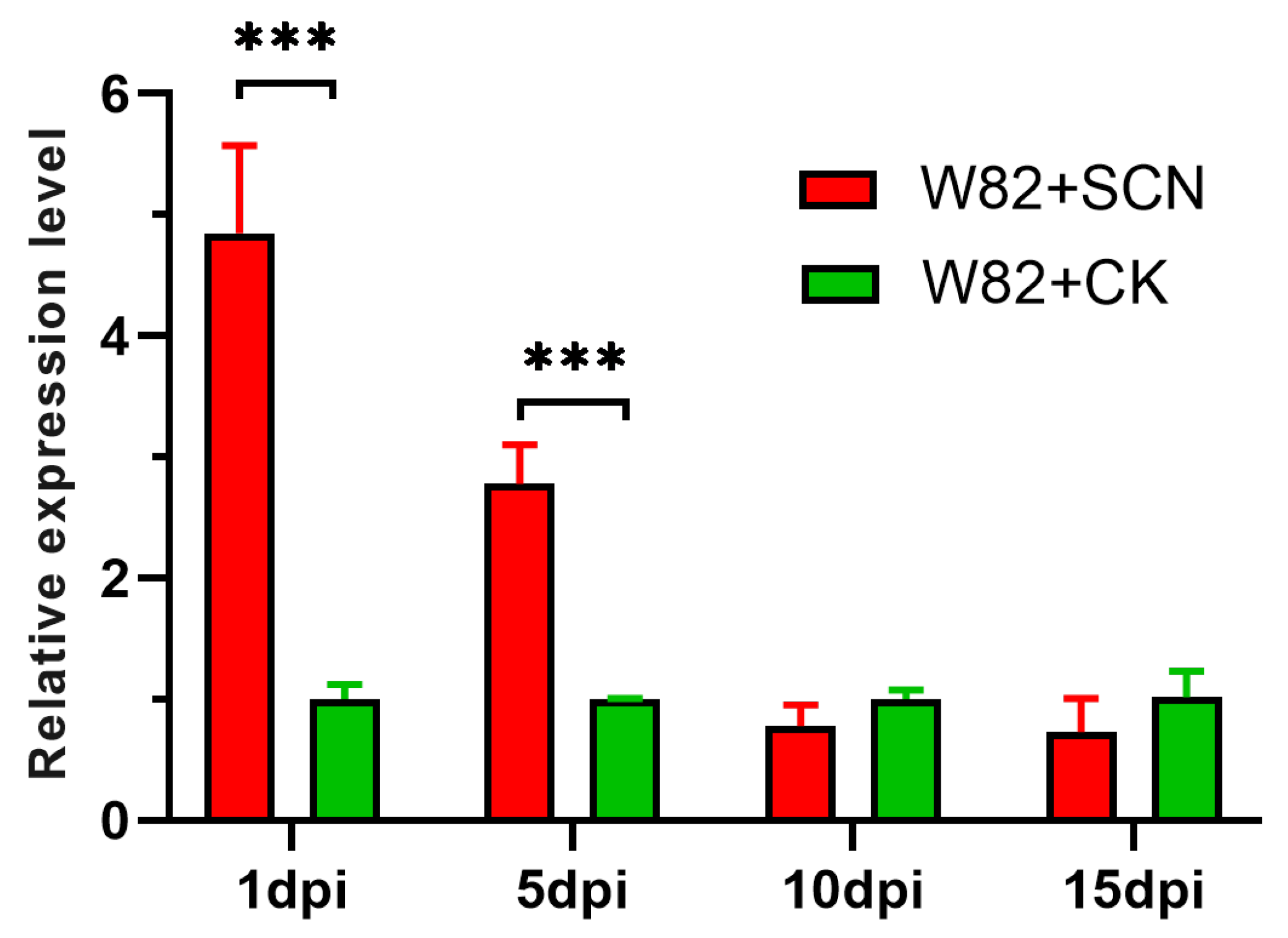
2.2. Gma-miR408 Response to Heterodera glycines Infection
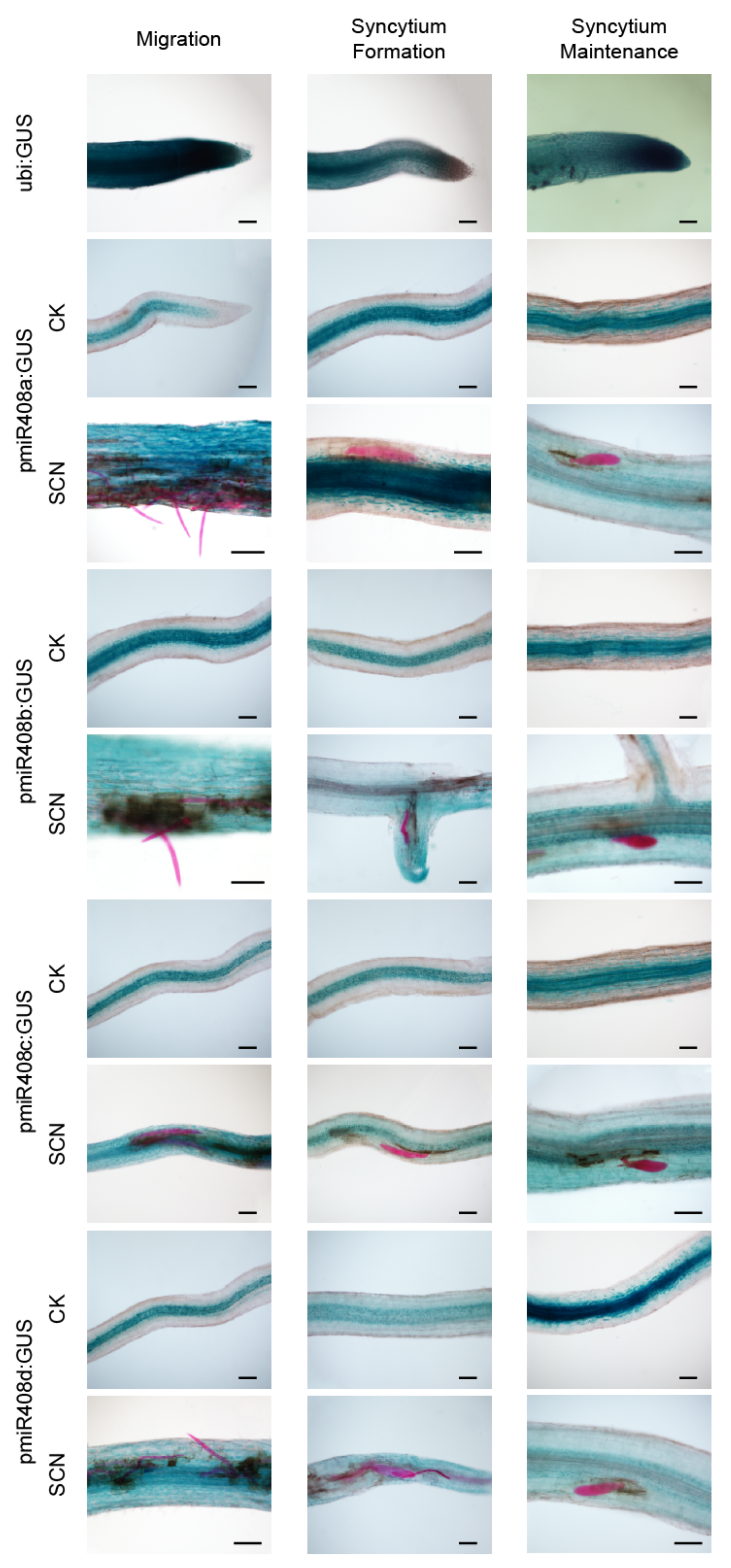
2.3. Expression Pattern of gma-miR408 in Soybean Roots
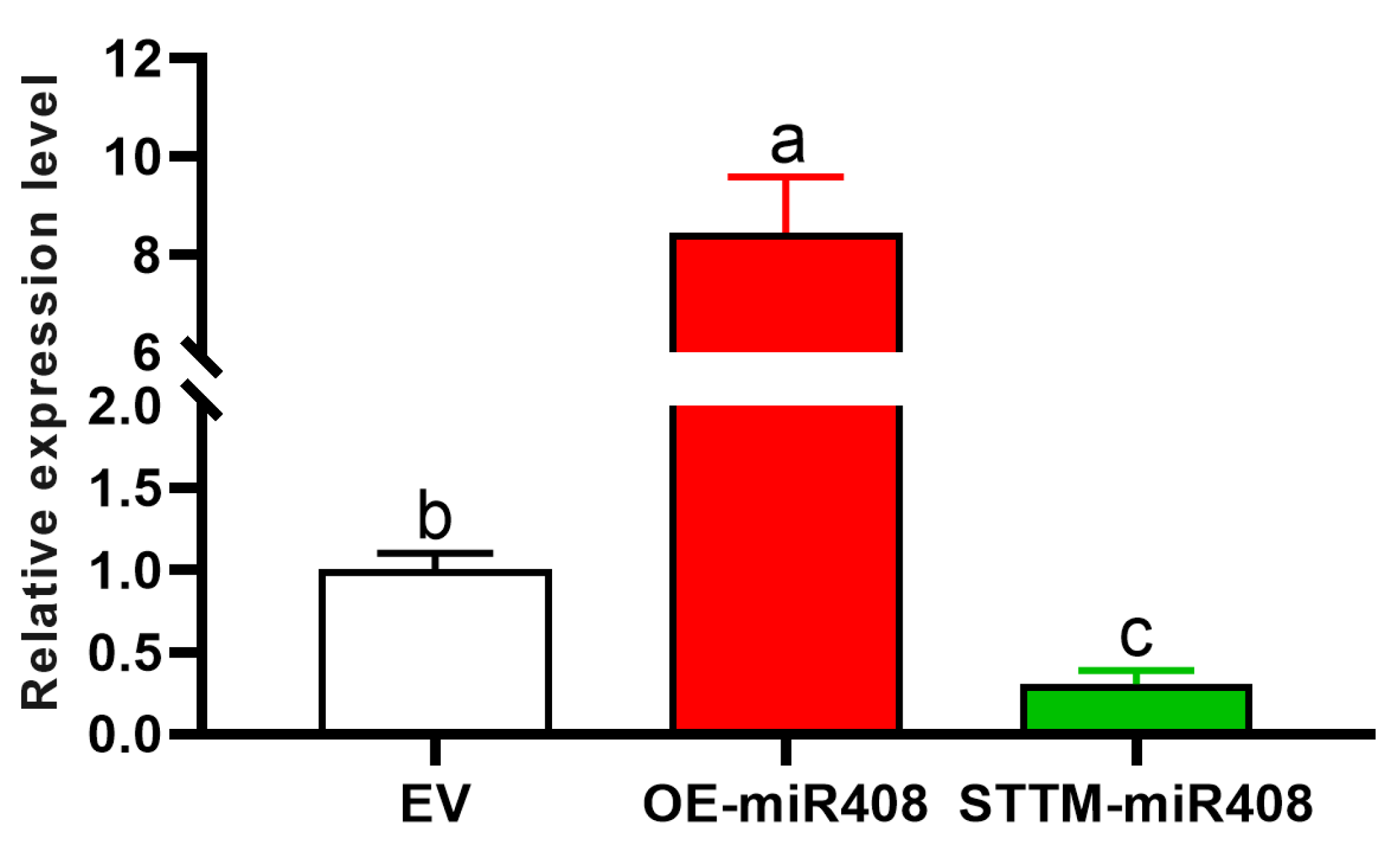
2.4. Induction of gma-miR408 Overexpression and Silencing Transgenic Soybean Hairy Roots
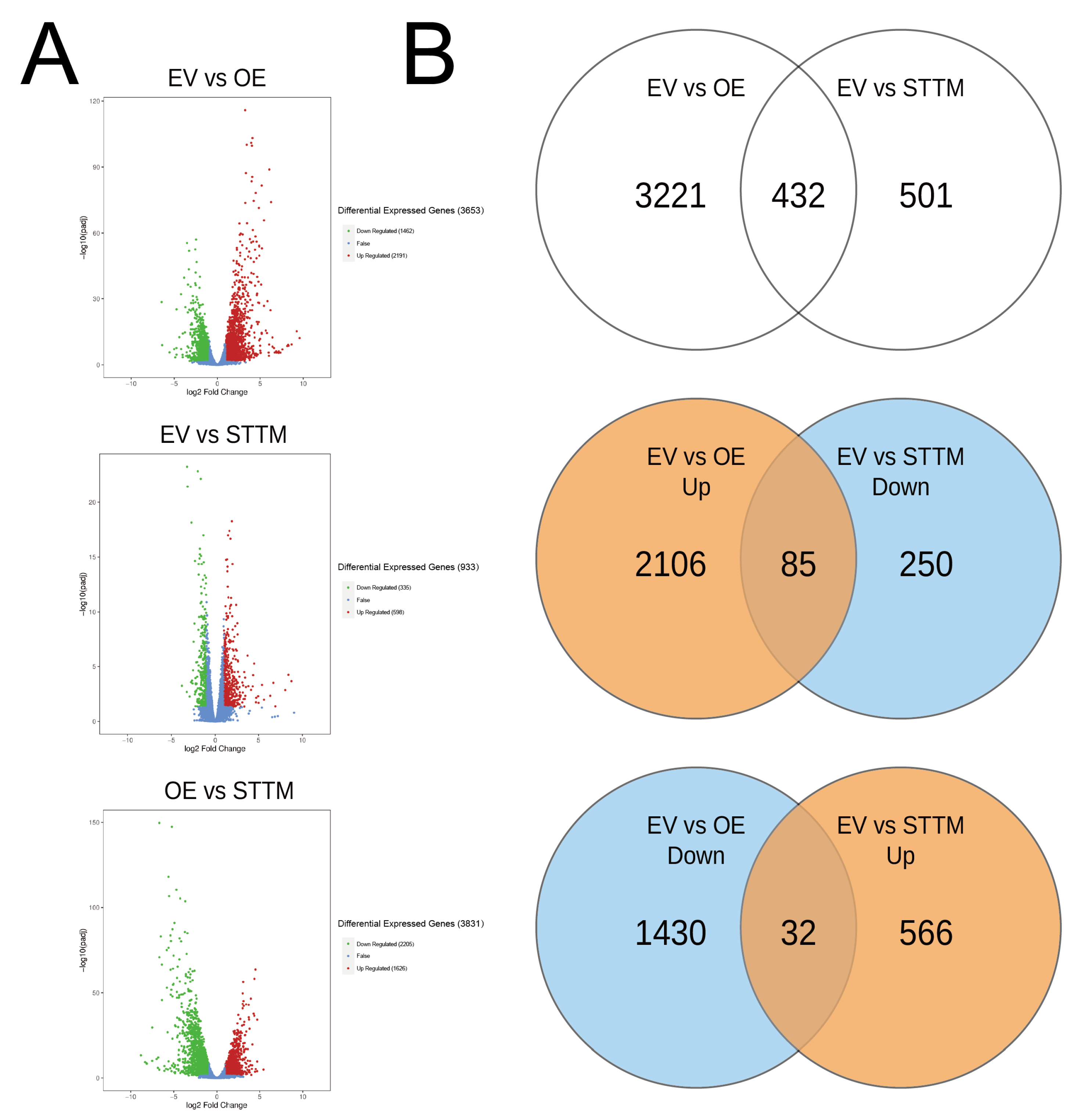
2.5. Transcriptomic Analyses for OE-miR408 and STTM-miR408
2.6. Differentially Expressed Genes for OE-miR408 and STTM-miR408
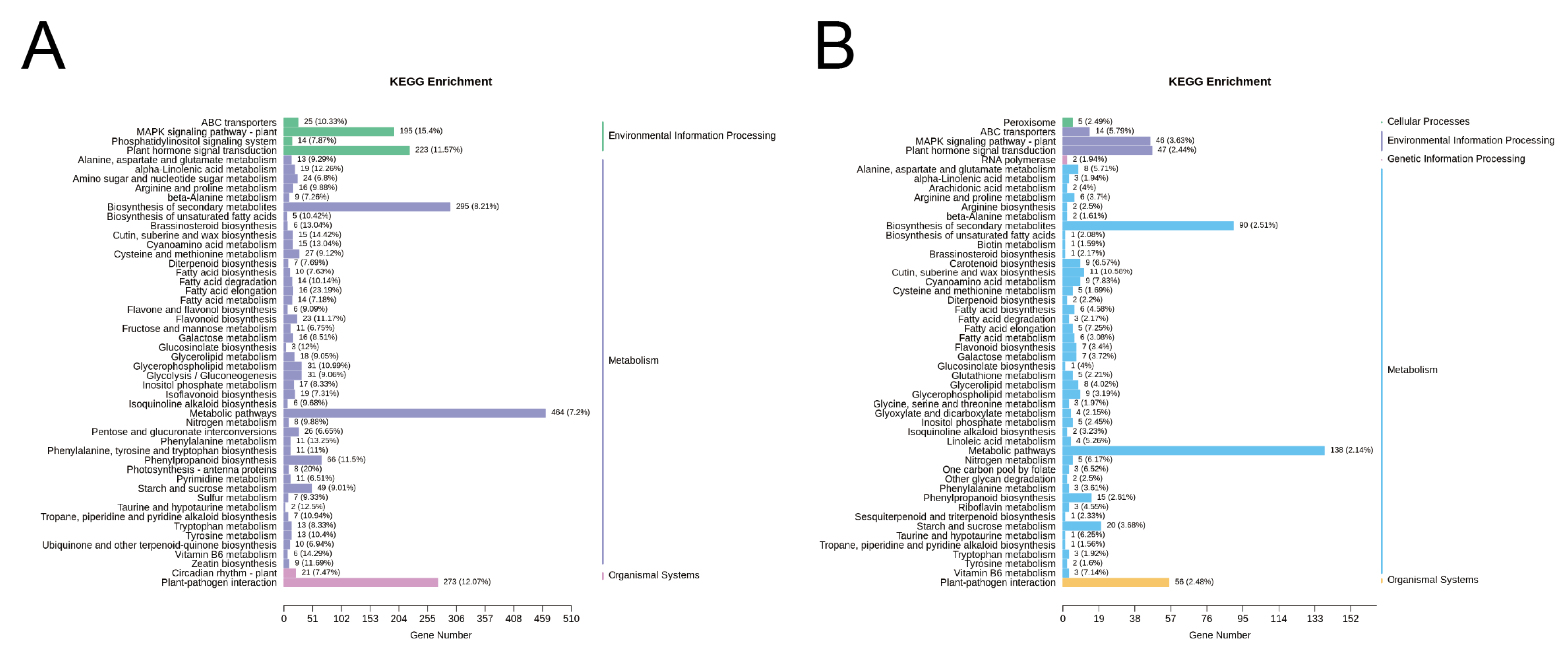
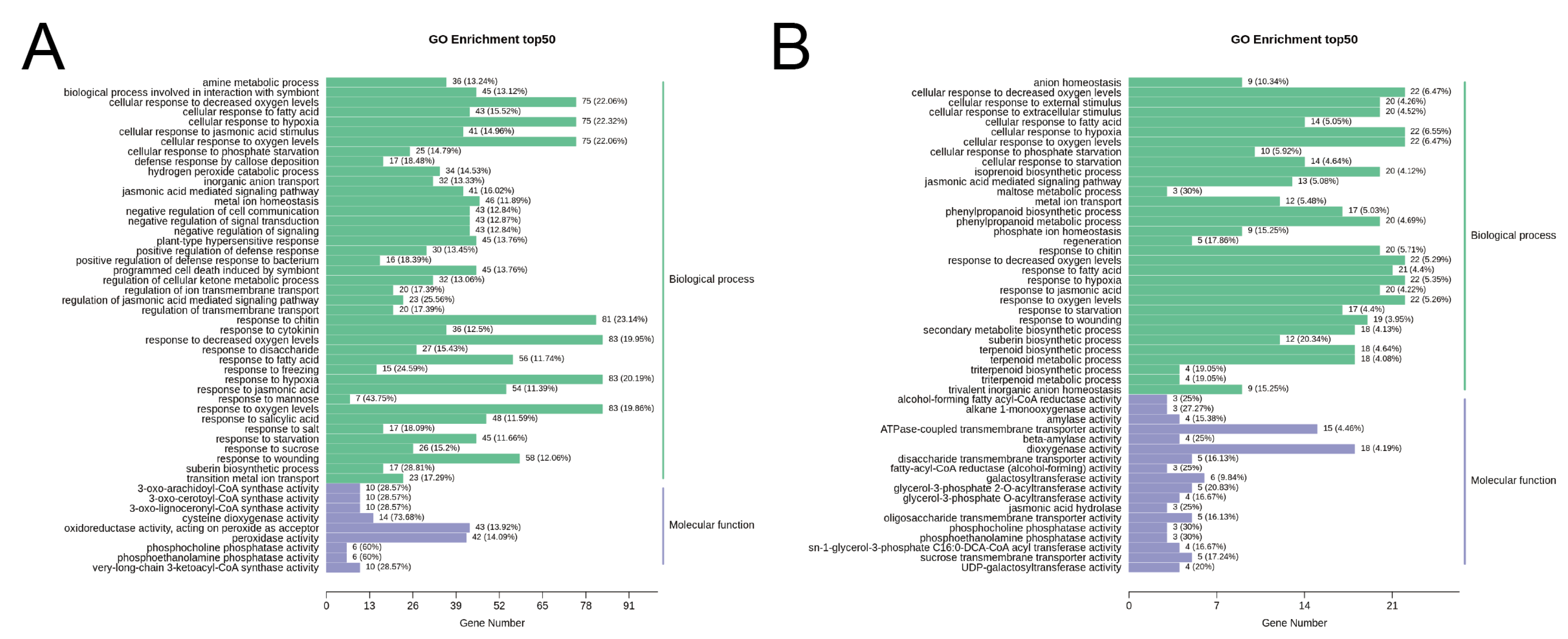
2.7. Functional Analysis and Clustering
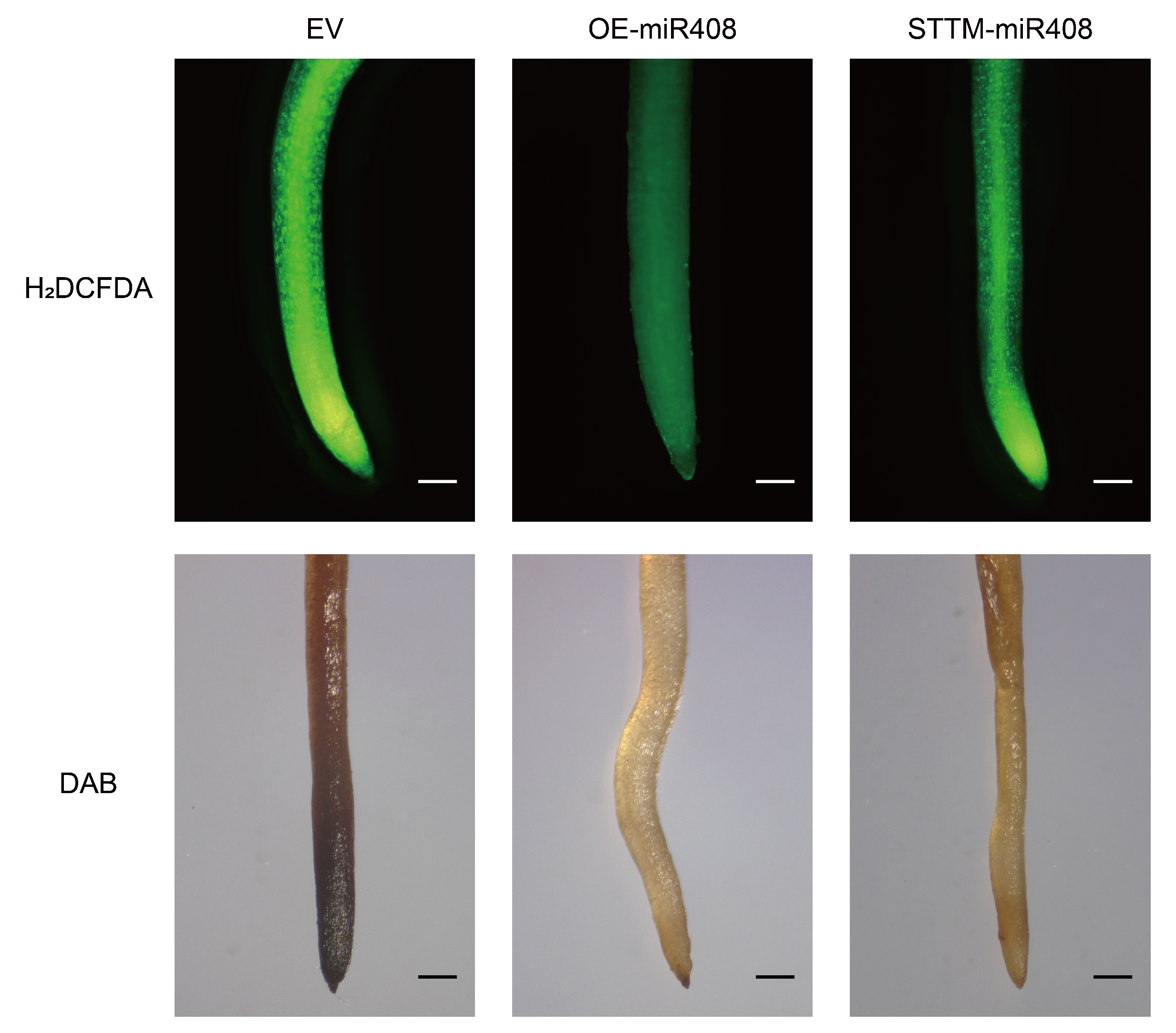
2.8. Gma-miR408 Suppressing Reactive Oxygen Species Accumulation
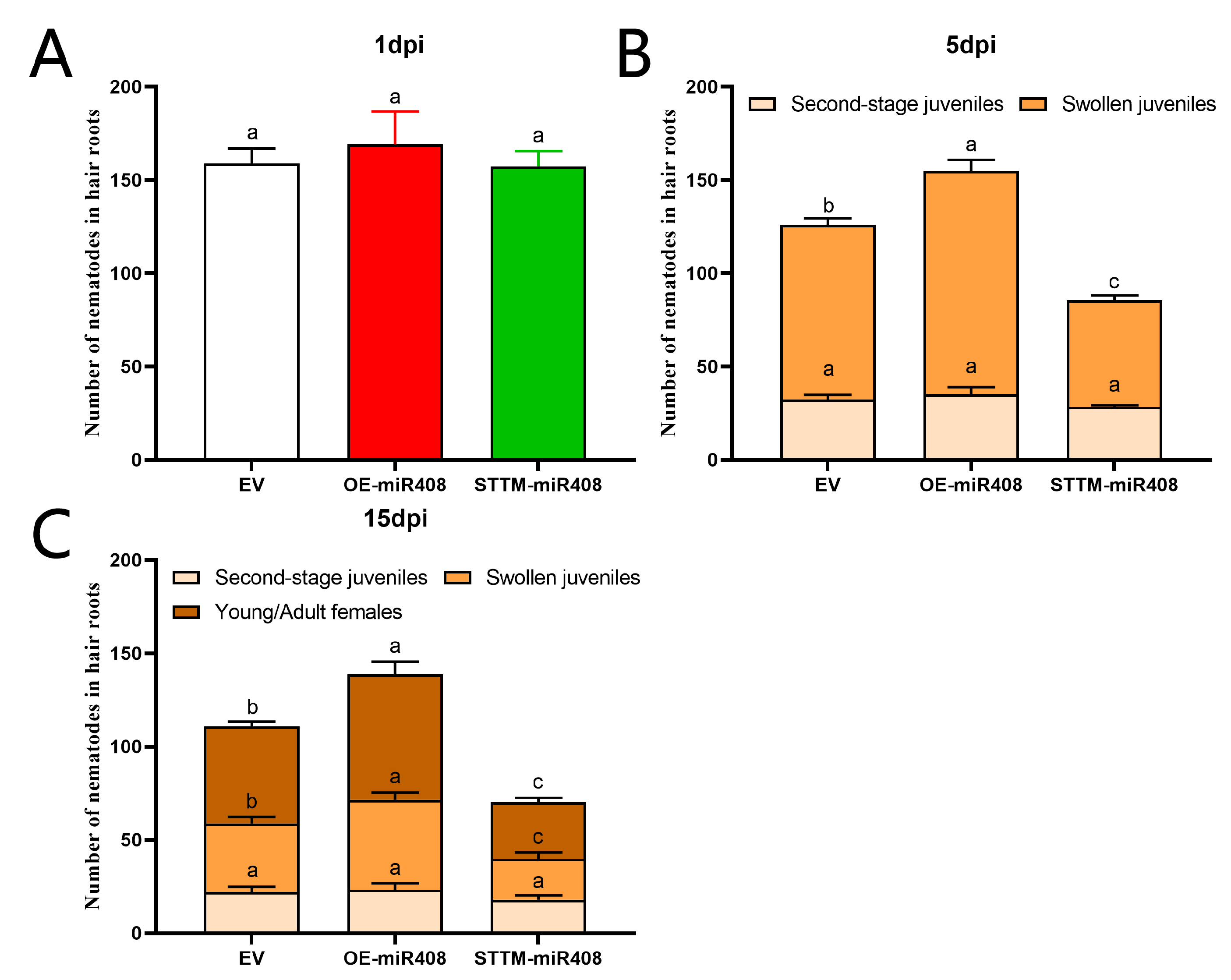
2.9. Gma-miR408 Enhances Soybean Cyst Nematode Susceptibility
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Nematode
4.3. Nematode Infection
4.4. Construction for Genetic Transformation Vectors
4.5. Transformation Process of Soybean Hairy Roots
4.6. Gene Expression Analysis of miR408
4.7. RNA Isolation, Reverse Transcription, and cDNA Libraries Sequencing
4.8. Differential Gene Analysis and Enrichment
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| W82 | Williams 82 |
| SCN | soybean cyst nematode |
| qRT-PCR | quantitative real-time PCR |
| dpi | day post-inoculation |
| J2s | second-stage juveniles |
| OE | overexpressing |
| STTM | Short tandem target mimic |
References
- Yu, Y.; Jia, T.; Chen, X. The ‘how’ and ‘where’ of plant micro RNAs. New Phytol. 2017, 216, 1002–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Zeng, Z.; Liu, Z.; Xia, R. Small RNAs, emerging regulators critical for the development of horticultural traits. Hortic. Res. 2018, 5, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Feng, B.; Gao, C.; Zhang, H.; Wen, F.; Tao, L.; Fu, G.; Xiong, J. The Evolution and Functional Roles of miR408 and Its Targets in Plants. Int. J. Mol. Sci. 2022, 23, 530. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, 155–162. [Google Scholar] [CrossRef]
- Chen, C.; Li, J.; Feng, J.; Liu, B.; Feng, L.; Yu, X.; Li, G.; Zhai, J.; Meyers, B.C.; Xia, R. sRNAanno—A database repository of uniformly annotated small RNAs in plants. Hortic. Res. 2021, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.P.; Yu, Y.; Feng, Y.Z.; Zhou, Y.F.; Zhang, F.; Yang, Y.W.; Lei, M.Q.; Zhang, Y.C.; Chen, Y.Q. MiR408 Regulates Grain Yield and Photosynthesis via a Phytocyanin Protein. Plant Physiol. 2017, 175, 1175–1185. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Zhang, Q.; Wang, Q.; Wang, X.; Liu, J.; Li, M.; Huang, L.; Kang, Z. Target of tae-miR408, a chemocyanin-like protein gene (TaCLP1), plays positive roles in wheat response to high-salinity, heavy cupric stress and stripe rust. Plant Mol. Biol. 2013, 83, 433–443. [Google Scholar] [CrossRef]
- Hajyzadeh, M.; Turktas, M.; Khawar, K.M.; Unver, T. miR408 overexpression causes increased drought tolerance in chickpea. Gene 2015, 555, 186–193. [Google Scholar] [CrossRef]
- Mutum, R.D.; Balyan, S.C.; Kansal, S.; Agarwal, P.; Kumar, S.; Kumar, M.; Raghuvanshi, S. Evolution of variety-specific regulatory schema for expression of osa-miR408 in indica rice varieties under drought stress. FEBS J. 2013, 280, 1717–1730. [Google Scholar] [CrossRef]
- Trindade, I.; Capitão, C.; Dalmay, T.; Fevereiro, M.P.; Santos, D.M.D. miR398 and miR408 are up-regulated in response to water deficit in Medicago truncatula. Planta 2010, 231, 705–716. [Google Scholar] [CrossRef]
- Kantar, M.; Unver, T.; Budak, H. Regulation of barley miRNAs upon dehydration stress correlated with target gene expression. Funct. Integr. Genom. 2010, 10, 493–507. [Google Scholar] [CrossRef] [PubMed]
- Eldem, V.; Çelikkol Akçay, U.; Ozhuner, E.; Bakır, Y.; Uranbey, S.; Unver, T. Genome-wide identification of miRNAs responsive to drought in peach (Prunus persica) by high-throughput deep sequencing. PLoS ONE 2012, 7, 50298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Liu, Y.; Liu, Z.; Kong, D.; Duan, M.; Luo, L. Genome-wide identification and analysis of drought-responsive microRNAs in Oryza sativa. J. Exp. Bot. 2010, 61, 4157–4168. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Burd, S.; Lers, A. miR408 is involved in abiotic stress responses in A rabidopsis. Plant J. 2015, 84, 169–187. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, Z.; Li, W.; Fang, C.; Shen, Y.; Li, C.; Wu, Y.; Tian, Z. Comprehensive analyses of microRNA gene evolution in paleopolyploid soybean genome. Plant J. 2013, 76, 332–344. [Google Scholar] [CrossRef]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Se, B. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [Green Version]
- Lei, P.; Qi, N.; Zhou, Y.; Wang, Y.; Zhu, X.; Xuan, Y.; Liu, X.; Fan, H.; Chen, L.; Duan, Y. Soybean miR159-GmMYB33 regulatory network involved in gibberellin-modulated resistance to Heterodera glycines. Int. J. Mol. Sci. 2021, 22, 13172. [Google Scholar] [CrossRef]
- Qi, N.; Yan, J.; Lei, P.; Zhu, X.; Liu, X.; Xuan, Y.; Fan, H.; Wang, Y.; Chen, L.; Duan, Y. Functional Characterization of Ubiquitination Genes in the Interaction of Soybean—Heterodera glycines. Int. J. Mol. Sci. 2022, 23, 10771. [Google Scholar] [CrossRef]
- Bybd Jr, D.; Kirkpatrick, T.; Barker, K. An improved technique for clearing and staining plant tissues for detection of nematodes. J. Nematol. 1983, 15, 142. [Google Scholar]
- Voinnet, O. Origin, biogenesis, and activity of plant microRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Axtell, M.J.; Bartel, D.P. Antiquity of microRNAs and their targets in land plants. Plant Cell 2005, 17, 1658–1673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fattash, I.; Voß, B.; Reski, R.; Hess, W.R.; Frank, W. Evidence for the rapid expansion of microRNA-mediated regulation in early land plant evolution. BMC Plant Biol. 2007, 7, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Alaba, S.; Piszczalka, P.; Pietrykowska, H.; Pacak, A.M.; Sierocka, I.; Nuc, P.W.; Singh, K.; Plewka, P.; Sulkowska, A.; Jarmolowski, A. The liverwort Pellia endiviifolia shares microtranscriptomic traits that are common to green algae and land plants. New Phytol. 2015, 206, 352–367. [Google Scholar] [CrossRef] [Green Version]
- Sunkar, R.; Zhu, J.K. Novel and stress-regulated microRNAs and other small RNAs from Arabidopsis. Plant Cell 2004, 16, 2001–2019. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Yang, J.; Cai, X.; Shen, Y.; Cui, N.; Zhu, Y.; Jia, B.; Sun, X. The opposite roles of OsmiR408 in cold and drought stress responses in Oryza sativa. Mol. Breed 2018, 38, 1–12. [Google Scholar] [CrossRef]
- Macovei, A.; Tuteja, N. microRNAs targeting DEAD-box helicases are involved in salinity stress response in rice (Oryza sativa L.). BMC Plant Biol. 2012, 12, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Niu, J.; Cao, X. Heterologous expression of Salvia miltiorrhiza microRNA408 enhances tolerance to salt stress in Nicotiana benthamiana. Int. J. Mol. Sci. 2018, 19, 3985. [Google Scholar] [CrossRef] [Green Version]
- Djami-Tchatchou, A.T.; Dubery, I.A. Lipopolysaccharide perception leads to dynamic alterations in the microtranscriptome of Arabidopsis thaliana cells and leaf tissues. BMC Plant Biol. 2015, 15, 79. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Liu, J.; Triplett, L.; Leach, J.E.; Wang, G.-L. Novel insights into rice innate immunity against bacterial and fungal pathogens. Annu. Rev. Phytopathol. 2014, 52, 213–241. [Google Scholar] [CrossRef] [PubMed]
- Gupta, O.P.; Permar, V.; Koundal, V.; Singh, U.D.; Praveen, S. MicroRNA regulated defense responses in Triticum aestivum L. during Puccinia graminis f. sp. tritici infection. Mol. Biol. Rep. 2012, 39, 817–2824. [Google Scholar] [CrossRef] [PubMed]
- Kereszt, A.; Li, D.; Indrasumunar, A.; Nguyen, C.D.; Nontachaiyapoom, S.; Kinkema, M.; Gresshoff, P.M. Agrobacterium rhizogenes-mediated transformation of soybean to study root biology. Nat. Protoc. 2007, 2, 948–952. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA Name | Location | Mature Sequence-5p | Mature Sequence-3p |
|---|---|---|---|
| gma-miR408a | Chr2:837419-837550 | CAGGGGAACAGGCAGAGCAUG | AUGCACUGCCUCUUCCCUGGC |
| gma-miR408b | Chr3:44626692-44626830 | CUGGGAACAGGCAGGGCACGA | AUGCACUGCCUCUUCCCUGGC |
| gma-miR408c | Chr10:36557001-36557134 | CAGGGGAACAGGCAGAGCAUG | AUGCACUGCCUCUUCCCUGGC |
| gma-miR408d | Chr19:47163684-47163821 | CUGGGAACAGGCAGAGCACGA | AUGCACUGCCUCUUCCCUGGC |
| miRNA Name | Precursor Sequence |
|---|---|
| gma-miR408a | ACAGGACAAAGCAGGGGAACAGGCAGAGCAUGGAUGGAGCUAUCAA |
| CACAAUAUUGUCAAGAAACUGAGAGUGAGAGGAGAAAUAUGUUGUG | |
| GUUCUGCUCAUGCACUGCCUCUUCCCUGGCUCUGUCUCCA | |
| gma-miR408b | CAGGACAAAGGCUGGGAACAGGCAGGGCACGAAUGGAGCUAUCAAC |
| AGAAAAUGGUAAAGUGAGAAUGAAAGGAGAGAGAGAGAGAGAGAUC | |
| UGUUGUGGCUACGCUCAUGCACUGCCUCUUCCCUGGCUCUGUCUCUU | |
| gma-miR408c | ACAGGACAAAGCAGGGGAACAGGCAGAGCAUGGAUGGAGCUAUCAA |
| CACAAUAUUGUCAAGAAAGUGAGAAAGUGAGAGGAGAAAUCUGUUG | |
| UGGUUCUGCUCAUGCACUGCCUCUUCCCUGGCUUUGUCUCUA | |
| gma-miR408d | CAGGGCAAAGGCUGGGAACAGGCAGAGCACGAAUGGAGCUAUCAAC |
| AGAAAAUGGAGAAGUGAGAAAUGAGAGGAGAGAAAGAGAGAGAUC | |
| UGUUAGGGCUACGCUCAUGCACUGCCUCUUCCCUGGCUCUGUCUCUU |
| ID | Raw Reads | Clean Reads | Mapped Reads (%) | Uniquely M | Multiple M | Q30(%) |
|---|---|---|---|---|---|---|
| EV-1 | 46,804,494 | 39,360,542 | 36,609,992 (93.01) | 35,970,124 | 639,868 | 90.89 |
| EV-2 | 48,333,980 | 41,035,690 | 38,123,699 (92.90) | 37,471,922 | 651,777 | 91.34 |
| EV-3 | 48,613,286 | 40,137,396 | 37,364,825 (93.09) | 36,697,461 | 667,364 | 91.25 |
| OE-1 | 48,044,828 | 38,188,248 | 35,289,502 (92.41) | 34,698,477 | 591,025 | 91.20 |
| OE-2 | 56,847,288 | 45,955,306 | 42,646,102 (92.80) | 41,945,756 | 700,346 | 91.84 |
| OE-3 | 57,874,234 | 52,788,600 | 49,278,779 (93.35) | 48,478,778 | 800,001 | 92.68 |
| STTM-1 | 47,124,578 | 38,307,708 | 35,646,187 (93.05) | 35,042,320 | 603,867 | 91.42 |
| STTM-2 | 50,267,440 | 40,530,212 | 37,142,081 (91.64) | 36,466,073 | 676,008 | 91.55 |
| STTM-3 | 48,159,886 | 38,251,754 | 34,301,873 (89.67) | 33,70,2718 | 599,155 | 91.13 |
| Total | 452,070,014 | 374,555,456 | 346,403,040 | 340,473,629 | 5,929,411 |
| Group | Total | Down Regulated | Up Regulate |
|---|---|---|---|
| EV vs OE | 3653 | 1462 | 2191 |
| EV vs STTM | 933 | 335 | 598 |
| OE vs STTM | 3831 | 2205 | 1626 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, Y.; Qi, N.; Lei, P.; Wang, Y.; Xuan, Y.; Liu, X.; Fan, H.; Chen, L.; Duan, Y.; Zhu, X. Gma-miR408 Enhances Soybean Cyst Nematode Susceptibility by Suppressing Reactive Oxygen Species Accumulation. Int. J. Mol. Sci. 2022, 23, 14022. https://doi.org/10.3390/ijms232214022
Feng Y, Qi N, Lei P, Wang Y, Xuan Y, Liu X, Fan H, Chen L, Duan Y, Zhu X. Gma-miR408 Enhances Soybean Cyst Nematode Susceptibility by Suppressing Reactive Oxygen Species Accumulation. International Journal of Molecular Sciences. 2022; 23(22):14022. https://doi.org/10.3390/ijms232214022
Chicago/Turabian StyleFeng, Yaxing, Nawei Qi, Piao Lei, Yuanyuan Wang, Yuanhu Xuan, Xiaoyu Liu, Haiyan Fan, Lijie Chen, Yuxi Duan, and Xiaofeng Zhu. 2022. "Gma-miR408 Enhances Soybean Cyst Nematode Susceptibility by Suppressing Reactive Oxygen Species Accumulation" International Journal of Molecular Sciences 23, no. 22: 14022. https://doi.org/10.3390/ijms232214022