
G Protein-Coupled Receptors Regulated by Membrane Potential

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Voltage Dependence of GPCRs
3. The Molecular Basis for Voltage Dependence of GPCRs
4. A Tyrosine-Based Voltage Sensor in the M2R
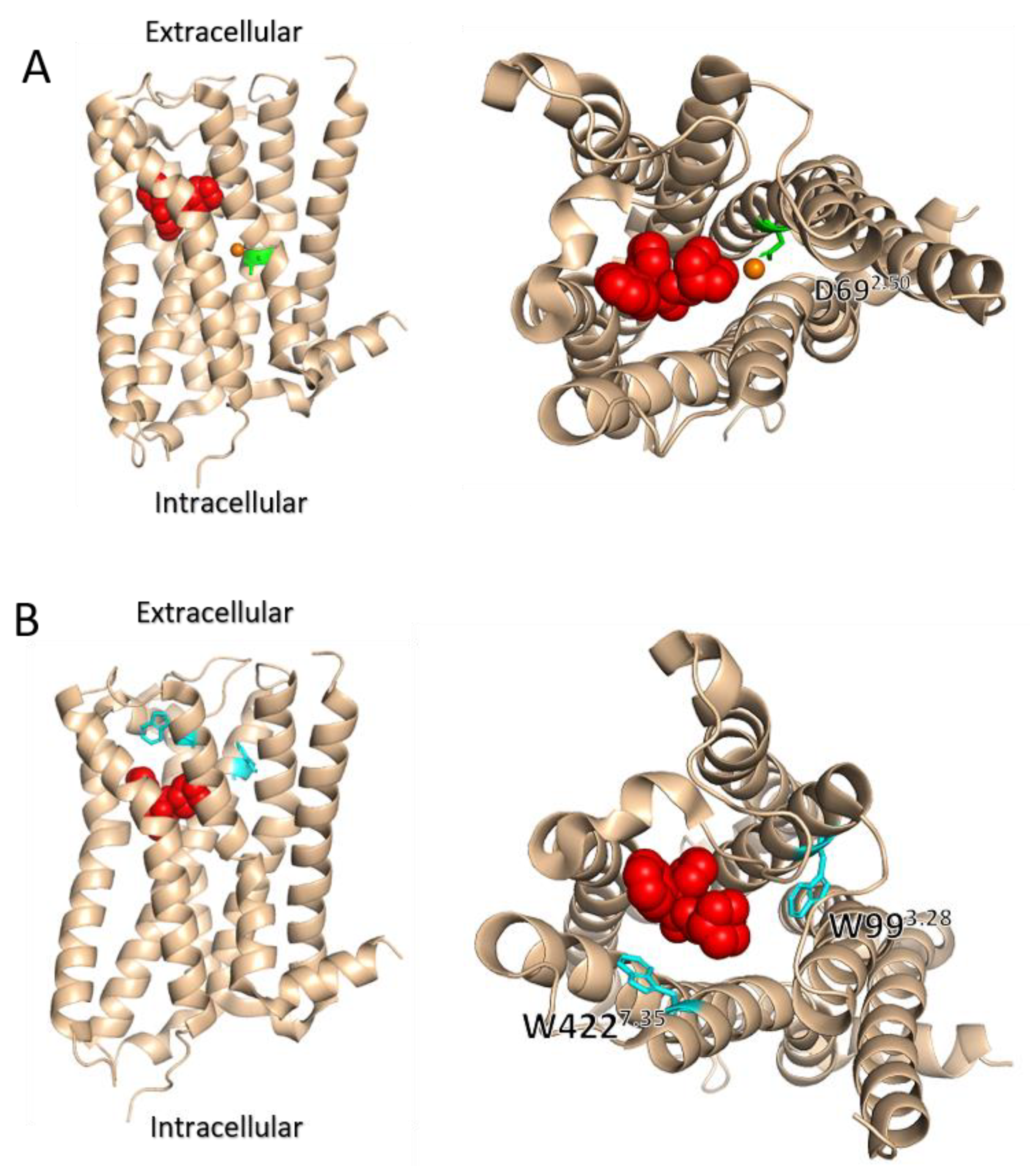
5. Role of the Ligand Binding Site in Determining the Voltage Dependence of GPCRs
6. G Protein Coupling as a Mechanism for Voltage Dependence of GPCRs
7. The Involvement of Sodium Ions in the Voltage Dependence of GPCRs
8. The Role of the Allosteric Binding Site in the Voltage Dependence of GPCRs
9. A Thermodynamics Perspective on Voltage Dependence of GPCRs
10. Physiological Roles of Voltage Dependence of GPCRs
Funding
Conflicts of Interest
References
- Hauser, A.S.; Attwood, M.M.; Rask-Andersen, M.; Schiöth, H.B.; Gloriam, D.E. Trends in GPCR Drug Discovery: New Agents, Targets and Indications. Nat. Rev. Drug Discov. 2017, 16, 829. [Google Scholar] [CrossRef] [PubMed]
- Weis, W.I.; Kobilka, B.K. The Molecular Basis of G Protein–Coupled Receptor Activation. Annu. Rev. Biochem. 2014, 87, 897–919. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Mayor, F.; Rondard, P.; Gurevich, V.V.; Gurevich, E.V. GPCR Signaling Regulation: The Role of GRKs and Arrestins. Front. Pharmacol. 2019, 10, 125. [Google Scholar] [CrossRef] [Green Version]
- Levin, M. Molecular Bioelectricity: How Endogenous Voltage Potentials Control Cell Behavior and Instruct Pattern Regulation in Vivo. Mol. Biol. Cell 2014, 25, 3835–3850. [Google Scholar] [CrossRef] [PubMed]
- Bezanilla, F.; Perozo, E. The Voltage Sensor and the Gate in Ion Channels. In Advances in Protein Chemistry; Academic Press: Cambridge, MA, USA, 2003; Volume 63, pp. 211–241. [Google Scholar] [CrossRef]
- Bezanilla, F. The Voltage Sensor in Voltage-Dependent Ion Channels. Physiol. Rev. 2000, 80, 555–592. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.L.; Westbrook, G.L. Mixed-agonist Action of Excitatory Amino Acids on Mouse Spinal Cord Neurones under Voltage Clamp. J. Physiol. 1984, 354, 29–53. [Google Scholar] [CrossRef] [Green Version]
- Nowak, L.; Bregestovski, P.; Ascher, P.; Herbet, A.; Prochiantz, A. Magnesium Gates Glutamate-Activated Channels in Mouse Central Neurones. Nature 1984, 307, 462–465. [Google Scholar] [CrossRef]
- Raman, I.M.; Trussell, L.O. Concentration-Jump Analysis of Voltage-Dependent Conductances Activated by Glutamate and Kainate in Neurons of the Avian Cochlear Nucleus. Biophys. J. 1995, 69, 1868–1879. [Google Scholar] [CrossRef] [Green Version]
- Magleby, K.L.; Stevens, C.F. The Effect of Voltage on the Time Course of End-Plate Currents. J. Physiol. 1972, 223, 151. [Google Scholar] [CrossRef]
- Okamura, Y.; Kawanabe, A.; Kawai, T. Voltage-Sensing Phosphatases: Biophysics, Physiology, and Molecular Engineering. Physiol. Rev. 2018, 98, 2097–2131. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Pinna, J.; Tolhurst, G.; Gurung, I.S.; Vandenberg, J.I.; Mahaut-Smith, M.P. Sensitivity Limits for Voltage Control of P2Y Receptor-Evoked Ca2+ Mobilization in the Rat Megakaryocyte. J. Physiol. 2004, 555, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Pinna, J.; Gurung, I.S.; Vial, C.; Leon, C.; Gachet, C.; Evans, R.J.; Mahaut-Smith, M.P. Direct Voltage Control of Signaling via P2Y1 and Other Gαq-Coupled Receptors*. J. Biol. Chem. 2005, 280, 1490–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, M.J.; Hussain, J.F.; Mahaut-Smith, M.P. A Novel Role for Membrane Potential in the Modulation of Intracellular Ca2+ Oscillations in Rat Megakaryocytes. J. Physiol. 2000, 524 Pt 2, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Bolton, T.B.; Zholos, A.V. Potential Synergy: Voltage-Driven Steps in Receptor-G Protein Coupling and Beyond. Science’s STKE 2003, 2003, pe52. [Google Scholar] [CrossRef] [PubMed]
- Zholos, A.V.; Bolton, T.B. G-Protein Control of Voltage Dependence as Well as Gating of Muscarinic Metabotropic Channels in Guinea-Pig Ileum. J. Physiol. 1994, 478 Pt 2, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Parnas, H.; Segel, L.; Dudel, J.; Parnas, I. Autoreceptors, Membrane Potential and the Regulation of Transmitter Release. Trends Neurosci. 2000, 23, 60–68. [Google Scholar] [CrossRef]
- Slutsky, I.; Rashkovan, G.; Parnas, H.; Parnas, I. Ca2+-Independent Feedback Inhibition of Acetylcholine Release in Frog Neuromuscular Junction. J. Neurosci. 2002, 22, 3426–3433. [Google Scholar] [CrossRef] [Green Version]
- Ong, B.H.; Ohsaga, A.; Sato, K.; Oshiro, T.; Shirato, K.; Maruyama, Y. G Protein Modulation of Voltage-Sensitive Muscarinic Receptor Signalling in Mouse Pancreatic Acinar Cells. Pflügers Arch. 2001, 441, 604–610. [Google Scholar] [CrossRef]
- Cohen-Armon, M.; Sokolovsky, M. Depolarization-Induced Changes in the Muscarinic Receptor in Rat Brain and Heart Are Mediated by Pertussis-Toxin-Sensitive G-Proteins. J. Biol. Chem. 1991, 266, 2595–2605. [Google Scholar] [CrossRef]
- Marty, A.; Tan, Y.P. The Initiation of Calcium Release Following Muscarinic Stimulation in Rat Lacrimal Glands. J. Physiol. 1989, 419, 665–687. [Google Scholar] [CrossRef]
- Ben-Chaim, Y.; Tour, O.; Dascal, N.; Parnas, I.; Parnas, H. The M2 Muscarinic G-Protein-Coupled Receptor Is Voltage-Sensitive. J. Biol. Chem 2003, 278, 22482–22491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahlholm, K.; Nilsson, J.; Marcellino, D.; Fuxe, K.; Århem, P. Voltage-Dependence of the Human Dopamine D2 Receptor. Synapse 2008, 62, 476–480. [Google Scholar] [CrossRef] [PubMed]
- Sahlholm, K.; Marcellino, D.; Nilsson, J.; Fuxe, K.; Århem, P. Differential Voltage-Sensitivity of D2-like Dopamine Receptors. Biochem. Biophys. Res. Commun 2008, 374, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Sahlholm, K.; Barchad-Avitzur, O.; Marcellino, D.; Gómez-Soler, M.; Fuxe, K.; Ciruela, F.; Århem, P. Agonist-Specific Voltage Sensitivity at the Dopamine D2S Receptor – Molecular Determinants and Relevance to Therapeutic Ligands. Neuropharmacology 2011, 61, 937–949. [Google Scholar] [CrossRef] [Green Version]
- Ågren, R.; Sahlholm, K. Voltage-Dependent Dopamine Potency at D1-Like Dopamine Receptors. Front. Pharmacol. 2020, 11, 1615. [Google Scholar] [CrossRef]
- Sahlholm, K.; Marcellino, D.; Nilsson, J.; Fuxe, K.; Århem, P. Voltage-Sensitivity at the Human Dopamine D2S Receptor Is Agonist-Specific. Biochem. Biophys. Res. Commun. 2008, 377, 1216–1221. [Google Scholar] [CrossRef]
- Moreno-Galindo, E.G.; Alamilla, J.; Sanchez-Chapula, J.A.; Tristani-Firouzi, M.; Navarro-Polanco, R.A. The Agonist-Specific Voltage Dependence of M2 Muscarinic Receptors Modulates the Deactivation of the Acetylcholine-Gated K+ Current (IKACh). Pflügers Arch. 2016, 468, 1207–1214. [Google Scholar] [CrossRef]
- Navarro-Polanco, R.A.; Galindo, E.G.M.; Ferrer-Villada, T.; Arias, M.; Rigby, J.R.; Sánchez-Chapula, J.A.; Tristani-Firouzi, M. Conformational Changes in the M2 Muscarinic Receptor Induced by Membrane Voltage and Agonist Binding. J. Physiol. 2011, 589, 1741–1753. [Google Scholar] [CrossRef]
- Rinne, A.; Mobarec, J.C.; Mahaut-Smith, M.; Kolb, P.; Bünemann, M. The Mode of Agonist Binding to a G Protein–Coupled Receptor Switches the Effect That Voltage Changes Have on Signaling. Sci. Signal. 2015, 8, ra110. [Google Scholar] [CrossRef]
- Ruland, J.G.; Kirchhofer, S.B.; Klindert, S.; Bailey, C.P.; Bünemann, M. Voltage Modulates the Effect of μ-Receptor Activation in a Ligand-Dependent Manner. Br. J. Pharmacol. 2020, 177, 3489–3504. [Google Scholar] [CrossRef]
- Ohana, L.; Barchad, O.; Parnas, I.; Parnas, H. The Metabotropic Glutamate G-Protein-Coupled Receptors MGluR3 and MGluR1a Are Voltage-Sensitive *. J. Biol. Chem. 2006, 281, 24204–24215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinne, A.; Birk, A.; Bünemann, M. Voltage Regulates Adrenergic Receptor Function. Proc. Natl. Acad. Sci. USA 2013, 110, 1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurz, M.; Krett, A.-L.; Bünemann, M. Voltage Dependence of Prostanoid Receptors. Mol. Pharmacol. 2020, 97, 267. [Google Scholar] [CrossRef]
- Bezanilla, F. Gating Currents. J. Gen. Physiol. 2018, 150, 911–932. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, C.M.; Bezanilla, F. Currents Related to Movement of the Gating Particles of the Sodium Channels. Nature 1973, 242, 459–461. [Google Scholar] [CrossRef]
- Ben-Chaim, Y.; Chanda, B.; Dascal, N.; Bezanilla, F.; Parnas, I.; Parnas, H. Movement of “gating Charge” Is Coupled to Ligand Binding in a G-Protein-Coupled Receptor. Nature 2006, 444, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.K.; MacKinnon, R. Contribution of the S4 Segment to Gating Charge in the Shaker K+ Channel. Neuron 1996, 16, 1169–1177. [Google Scholar] [CrossRef] [Green Version]
- Schoppa, N.E.; McCormack, K.; Tanouye, M.A.; Sigworth, F.J. The size of gating charge in wild-type and mutant Shaker potassium channels. Science 1992, 255, 1712–1715. [Google Scholar] [CrossRef]
- Dekel, N.; Priest, M.F.; Parnas, H.; Parnas, I.; Bezanilla, F. Depolarization Induces a Conformational Change in the Binding Site Region of the M2 Muscarinic Receptor. Proc. Natl. Acad. Sci. USA 2012, 109, 285–290. [Google Scholar] [CrossRef]
- Bezanilla, F. How Membrane Proteins Sense Voltage. Nat. Rev. Mol. Cell Biol. 2008, 9, 323–332. [Google Scholar] [CrossRef]
- Seoh, S.-A.; Sigg, D.; Papazian, D.M.; Bezanilla, F. Voltage-Sensing Residues in the S2 and S4 Segments of the Shaker K+ Channel. Neuron 1996, 16, 1159–1167. [Google Scholar] [CrossRef] [Green Version]
- Barchad-Avitzur, O.; Priest, M.F.; Dekel, N.; Bezanilla, F.; Parnas, H.; Ben-Chaim, Y. A Novel Voltage Sensor in the Orthosteric Binding Site of the M2 Muscarinic Receptor. Biophys. J. 2018, 111, 1396–1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ågren, R.; Sahlholm, K.; Nilsson, J.; Århem, P. Point Mutation of a Conserved Aspartate, D69, in the Muscarinic M2 Receptor Does Not Modify Voltage-Sensitive Agonist Potency. Biochem. Biophys. Res. Commun. 2018, 496, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Momany, F.A.; McGuire, R.F.; Burgess, A.W.; Scheraga, H.A. Energy Parameters in Polypeptides. VII. Geometric Parameters, Partial Atomic Charges, Nonbonded Interactions, Hydrogen Bond Interactions, and Intrinsic Torsional Potentials for the Naturally Occurring Amino Acids. J. Phys. Chem. 1975, 79, 2361–2381. [Google Scholar] [CrossRef]
- Rozenfeld, E.; Tauber, M.; Ben-Chaim, Y.; Parnas, M. GPCR Voltage Dependence Controls Neuronal Plasticity and Behavior. Nat. Commun. 2021, 12, 7252. [Google Scholar] [CrossRef]
- Kruse, A.C.; Ring, A.M.; Manglik, A.; Hu, J.; Hu, K.; Eitel, K.; Hübner, H.; Pardon, E.; Valant, C.; Sexton, P.M.; et al. Activation and Allosteric Modulation of a Muscarinic Acetylcholine Receptor. Nature 2013, 504, 101–106. [Google Scholar] [CrossRef] [Green Version]
- DeVree, B.T.; Mahoney, J.P.; Vélez-Ruiz, G.A.; Rasmussen, S.G.F.; Kuszak, A.J.; Edwald, E.; Fung, J.-J.; Manglik, A.; Masureel, M.; Du, Y.; et al. Allosteric Coupling from G Protein to the Agonist-Binding Pocket in GPCRs. Nature 2016, 535, 182. [Google Scholar] [CrossRef] [Green Version]
- Birk, A.; Rinne, A.; Bünemann, M. Membrane Potential Controls the Efficacy of Catecholamine-Induced Β1-Adrenoceptor Activity. J. Biol. Chem. 2015, 290, 27311–27320. [Google Scholar] [CrossRef] [Green Version]
- Haga, K.; Kruse, A.C.; Asada, H.; Yurugi-Kobayashi, T.; Shiroishi, M.; Zhang, C.; Weis, W.I.; Okada, T.; Kobilka, B.K.; Haga, T.; et al. Structure of the Human M2 Muscarinic Acetylcholine Receptor Bound to an Antagonist. Nature 2012, 482, 547–551. [Google Scholar] [CrossRef]
- Bezanilla, F.; Villalba-Galea, C.A. The Gating Charge Should Not Be Estimated by Fitting a Two-State Model to a Q-V Curve. J. Gen. Physiol. 2013, 142, 575–578. [Google Scholar] [CrossRef] [Green Version]
- Ben-Chaim, Y.; Broide, C.; Parnas, H. The Coupling of the M2 Muscarinic Receptor to Its G Protein Is Voltage Dependent. PLoS ONE 2019, 14, e0224367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, B.S.; Lefkowitz, R.J. Agonist-Specific Effects of Monovalent and Divalent Cations on Adenylate Cyclase-Coupled Alpha Adrenergic Receptors in Rabbit Platelets. Mol. Pharmacol. 1978, 14, 540–548. [Google Scholar] [PubMed]
- Gao, Z.-G.; Ijzerman, A.P. Allosteric Modulation of A2A Adenosine Receptors by Amiloride Analogues and Sodium Ions. Biochem. Pharmacol. 2000, 60, 669–676. [Google Scholar] [CrossRef]
- Neve, K.A. Regulation of Dopamine D2 Receptors by Sodium and PH. Mol. Pharmacol. 1991, 39, 570. [Google Scholar]
- Katritch, V.; Fenalti, G.; Abola, E.E.; Roth, B.L.; Cherezov, V.; Stevens, R.C. Allosteric Sodium in Class A GPCR Signaling. Trends Biochem. Sci. 2014, 39, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Fenalti, G.; Giguere, P.M.; Katritch, V.; Huang, X.-P.; Thompson, A.A.; Cherezov, V.; Roth, B.L.; Stevens, R.C. Molecular Control of δ-Opioid Receptor Signalling. Nature 2014, 506, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Miller-Gallacher, J.L.; Nehme, R.; Warne, T.; Edwards, P.C.; Schertler, G.F.X.; Leslie, A.G.W.; Tate, C.G. The 2.1 Å Resolution Structure of Cyanopindolol-Bound β1-Adrenoceptor Identifies an Intramembrane Na+ Ion That Stabilises the Ligand-Free Receptor. PLoS ONE 2014, 9, e92727. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Chun, E.; Thompson, A.A.; Chubukov, P.; Xu, F.; Katritch, V.; Han, G.W.; Roth, C.B.; Heitman, L.H.; IJzerman, A.P. Structural Basis for Allosteric Regulation of GPCRs by Sodium Ions. Science 2012, 337, 232–236. [Google Scholar] [CrossRef] [Green Version]
- Ballesteros, J.A.; Weinstein, H. Integrated Methods for the Construction of Three-Dimensional Models and Computational Probing of Structure-Function Relations in G Protein-Coupled Receptors. In Methods in Neurosciences; Elsevier: Amsterdam, The Netherlands, 1995; Volume 25, pp. 366–428. [Google Scholar] [CrossRef]
- Horstman, D.A.; Brandon, S.; Wilson, A.L.; Guyer, C.A.; Cragoe, E.J.; Limbird, L.E. An Aspartate Conserved among G-Protein Receptors Confers Allosteric Regulation of Alpha 2-Adrenergic Receptors by Sodium. J. Biol. Chem. 1990, 265, 21590–21595. [Google Scholar] [CrossRef]
- Barbhaiya, H.; McClain, R.; Ijzerman, A.; Rivkees, S.A. Site-Directed Mutagenesis of the Human A1 Adenosine Receptor: Influences of Acidic and Hydroxy Residues in the First Four Transmembrane Domains on Ligand Binding. Mol. Pharmacol. 1996, 50, 1635–1642. [Google Scholar]
- Wilson, M.H.; Highfield, H.A.; Limbird, L.E. The Role of a Conserved Inter-Transmembrane Domain Interface in Regulating α2a-Adrenergic Receptor Conformational Stability and Cell-Surface Turnover. Mol. Pharmacol. 2001, 59, 929. [Google Scholar] [CrossRef] [PubMed]
- Neve, K.A.; Cox, B.A.; Henningsen, R.A.; Spanoyannis, A.; Neve, R.L. Pivotal Role for Aspartate-80 in the Regulation of Dopamine D2 Receptor Affinity for Drugs and Inhibition of Adenylyl Cyclase. Mol. Pharmacol. 1991, 39, 733. [Google Scholar] [PubMed]
- Vickery, O.N.; Machtens, J.-P.; Zachariae, U. Membrane Potentials Regulating GPCRs: Insights from Experiments and Molecular Dynamics Simulations. Curr. Opin. Pharmacol. 2016, 30, 44–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vickery, O.N.; Machtens, J.-P.; Tamburrino, G.; Seeliger, D.; Zachariae, U. Structural Mechanisms of Voltage Sensing in G Protein-Coupled Receptors. Structure 2016, 24, 997–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, S.; Tauber, M.; Ben-chaim, Y. Sodium Ions Allosterically Modulate the M2 Muscarinic Receptor. Sci. Rep. 2020, 10, 11177. [Google Scholar] [CrossRef]
- Tauber, M.; ben Chaim, Y. The Activity of the Serotonergic 5-HT1A Receptor Is Modulated by Voltage and Sodium Levels. J. Biol. Chem. 2022, 298, 101978. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, A.; Marti-Solano, M.; Drabek, M.; Bünemann, M.; Kolb, P.; Rinne, A. The Allosteric Site Regulates the Voltage Sensitivity of Muscarinic Receptors. Cell. Signal. 2018, 42, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.C.; Sun, K.; Zhang, L.; Li, X.; Cao, C. GPCR Activation: Protonation and Membrane Potential. Protein Cell 2013, 4, 747–760. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.C.; Zhou, Y.; Cao, C. Thermodynamics of GPCR Activation. Biophys. Rep. 2015, 1, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Palczewski, K.; Kumasaka, T.; Hori, T.; Behnke, C.A.; Motoshima, H.; Fox, B.A.; Le Trong, I.; Teller, D.C.; Okada, T.; Stenkamp, R.E.; et al. Crystal Structure of Rhodopsin: A G Protein-Coupled Receptor. Science 2000, 289, 739–745. [Google Scholar] [CrossRef]
- Roth, C.B.; Hanson, M.A.; Stevens, R.C. Stabilization of the Human β2-Adrenergic Receptor TM4–TM3–TM5 Helix Interface by Mutagenesis of Glu1223.41, A Critical Residue in GPCR Structure. J. Mol. Biol 2008, 376, 1305–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarzycka, B.; Zaidi, S.A.; Roth, B.L.; Katritch, V. Harnessing Ion-Binding Sites for GPCR Pharmacology. Pharmacol. Rev. 2019, 71, 571. [Google Scholar] [CrossRef] [PubMed]
- Illergård, K.; Kauko, A.; Elofsson, A. Why Are Polar Residues within the Membrane Core Evolutionary Conserved? Proteins Struct. Funct. Bioinform. 2011, 79, 79–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angel, T.E.; Chance, M.R.; Palczewski, K. Conserved Waters Mediate Structural and Functional Activation of Family A (Rhodopsin-like) G Protein-Coupled Receptors. Proc. Natl. Acad. Sci. USA 2009, 106, 8555–8560. [Google Scholar] [CrossRef] [Green Version]
- Decoursey, T.E. Voltage-Gated Proton Channels and Other Proton Transfer Pathways. Physiol. Rev. 2003, 83, 475–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parnas, H.; Parnas, I. The Chemical Synapse Goes Electric: Ca2+- and Voltage-Sensitive GPCRs Control Neurotransmitter Release. Trends Neurosci. 2007, 30, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Parnas, I.; Parnas, H. Control of Neurotransmitter Release: From Ca2+ to Voltage Dependent G-Protein Coupled Receptors. Pflugers Arch. 2010, 460, 975–990. [Google Scholar] [CrossRef]
- Linial, M.; Ilouz, N.; Parnas, H. Voltage-Dependent Interaction Between the Muscarinic ACh Receptor and Proteins of the Exocytic Machinery. J. Physiol. 1997, 504, 251–258. [Google Scholar] [CrossRef]
- Ilouz, N.; Branski, L.; Parnis, J.; Parnas, H.; Linial, M. Depolarization affects the binding properties of muscarinic acetylcholine receptors and their interaction with proteins of the exocytic apparatus. J. Biol. Chem. 1999, 274, 29519–29528. [Google Scholar] [CrossRef] [Green Version]
- Kupchik, Y.M.; Barchad-Avitzur, O.; Wess, J.; Ben-Chaim, Y.; Parnas, I.; Parnas, H. A Novel Fast Mechanism for GPCR-Mediated Signal Transduction—Control of Neurotransmitter Release. J. Cell Biol. 2011, 192, 137–151. [Google Scholar] [CrossRef]
- Moreno-Galindo, E.G.; Sánchez-Chapula, J.A.; Sachse, F.B.; Rodríguez-Paredes, J.A.; Tristani-Firouzi, M.; Navarro-Polanco, R.A. Relaxation Gating of the Acetylcholine-Activated Inward Rectifier K+ Current Is Mediated by Intrinsic Voltage Sensitivity of the Muscarinic Receptor. J. Physiol. 2011, 589, 1755–1767. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Fajardo, P.D.; Aréchiga-Figueroa, I.A.; López-Serrano, A.L.; Rodriguez-Elias, J.C.; Alamilla, J.; Sánchez-Chapula, J.A.; Tristani-Firouzi, M.; Navarro-Polanco, R.A.; Moreno-Galindo, E.G. The Voltage-Sensitive Cardiac M2 Muscarinic Receptor Modulates the Inward Rectification of the G Protein-Coupled, ACh-Gated K+ Current. Pflugers Arch. 2018, 470, 1765–1776. [Google Scholar] [CrossRef] [PubMed]
- Goldberger, E.; Tauber, M.; Ben-Chaim, Y. Voltage Dependence of the Cannabinoid CB1 Receptor. Front. Pharmacol. 2022, 13, 1022275. [Google Scholar] [CrossRef]
- Luchicchi, A.; Pistis, M. Anandamide and 2-Arachidonoylglycerol: Pharmacological Properties, Functional Features, and Emerging Specificities of the Two Major Endocannabinoids. Mol. Neurobiol. 2012, 46, 374–392. [Google Scholar] [CrossRef] [PubMed]
- Straiker, A.; Mackie, K. Cannabinoid Signaling in Inhibitory Autaptic Hippocampal Neurons. Neuroscience 2009, 163, 190–201. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
David, D.; Bentulila, Z.; Tauber, M.; Ben-Chaim, Y. G Protein-Coupled Receptors Regulated by Membrane Potential. Int. J. Mol. Sci. 2022, 23, 13988. https://doi.org/10.3390/ijms232213988
David D, Bentulila Z, Tauber M, Ben-Chaim Y. G Protein-Coupled Receptors Regulated by Membrane Potential. International Journal of Molecular Sciences. 2022; 23(22):13988. https://doi.org/10.3390/ijms232213988
Chicago/Turabian StyleDavid, Dekel, Ziv Bentulila, Merav Tauber, and Yair Ben-Chaim. 2022. "G Protein-Coupled Receptors Regulated by Membrane Potential" International Journal of Molecular Sciences 23, no. 22: 13988. https://doi.org/10.3390/ijms232213988