
Conservation of Importin α Function in Apicomplexans: Ivermectin and GW5074 Target Plasmodium falciparum Importin α and Inhibit Parasite Growth in Culture


Abstract
:1. Introduction
2. Results
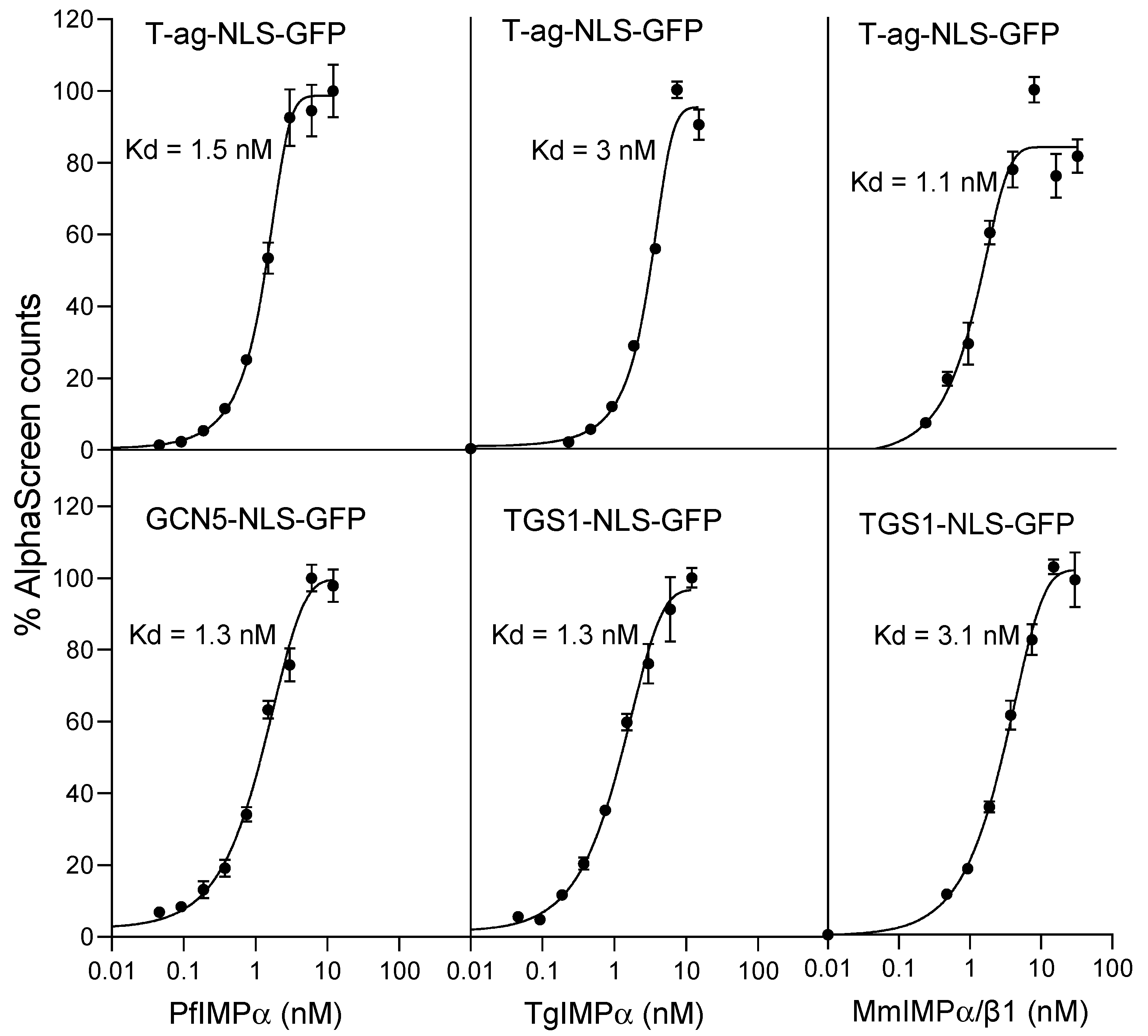
2.1. Apicomplexan NLSs Can Be Recognised with High Affinity by Mammalian IMPα/β1; Apicomplexan IMPαs Can Bind a Heterologous NLS with High Affinity
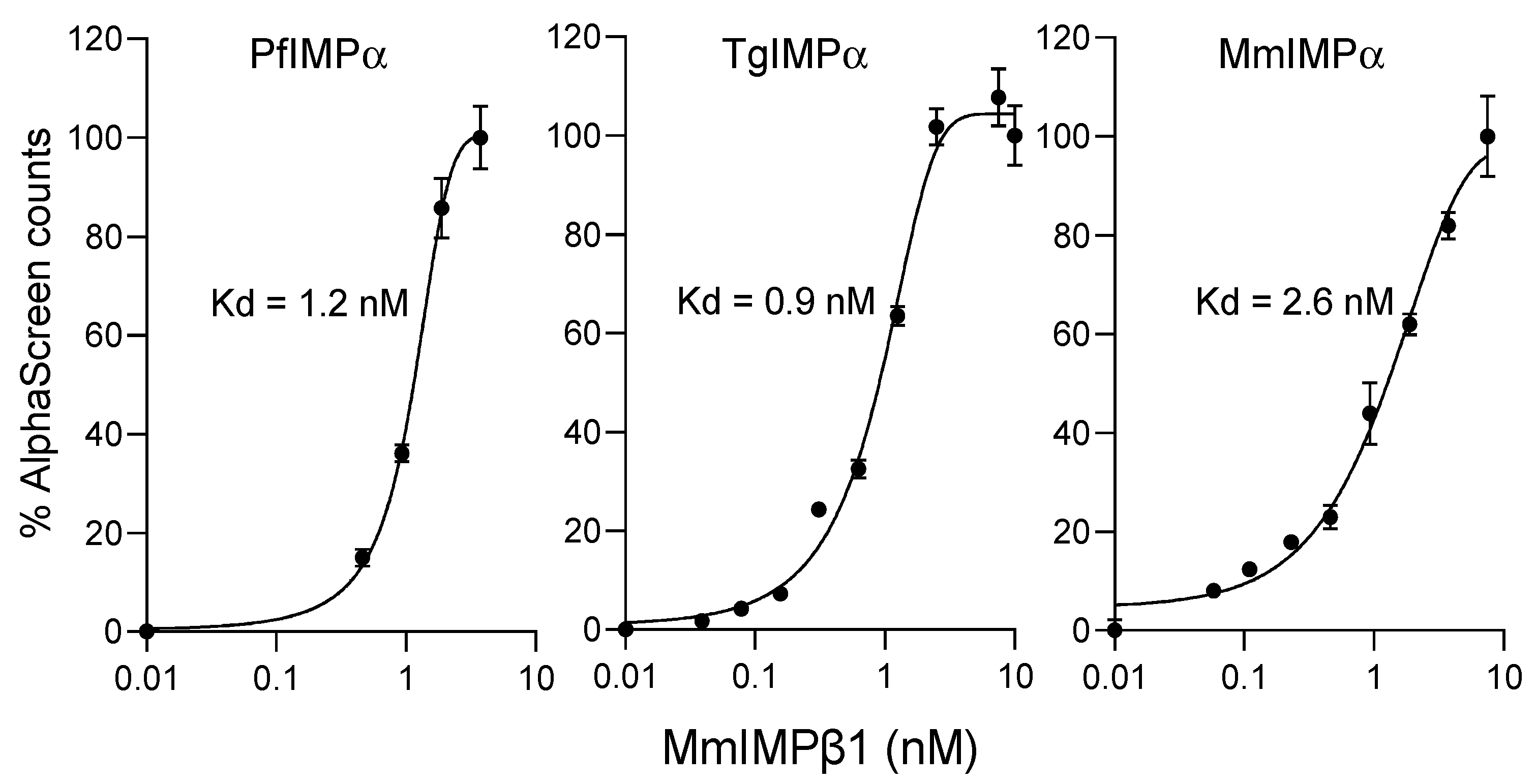
2.2. Apicomplexan IMPα Can Bind to MmIMPβ1 with High Affinity
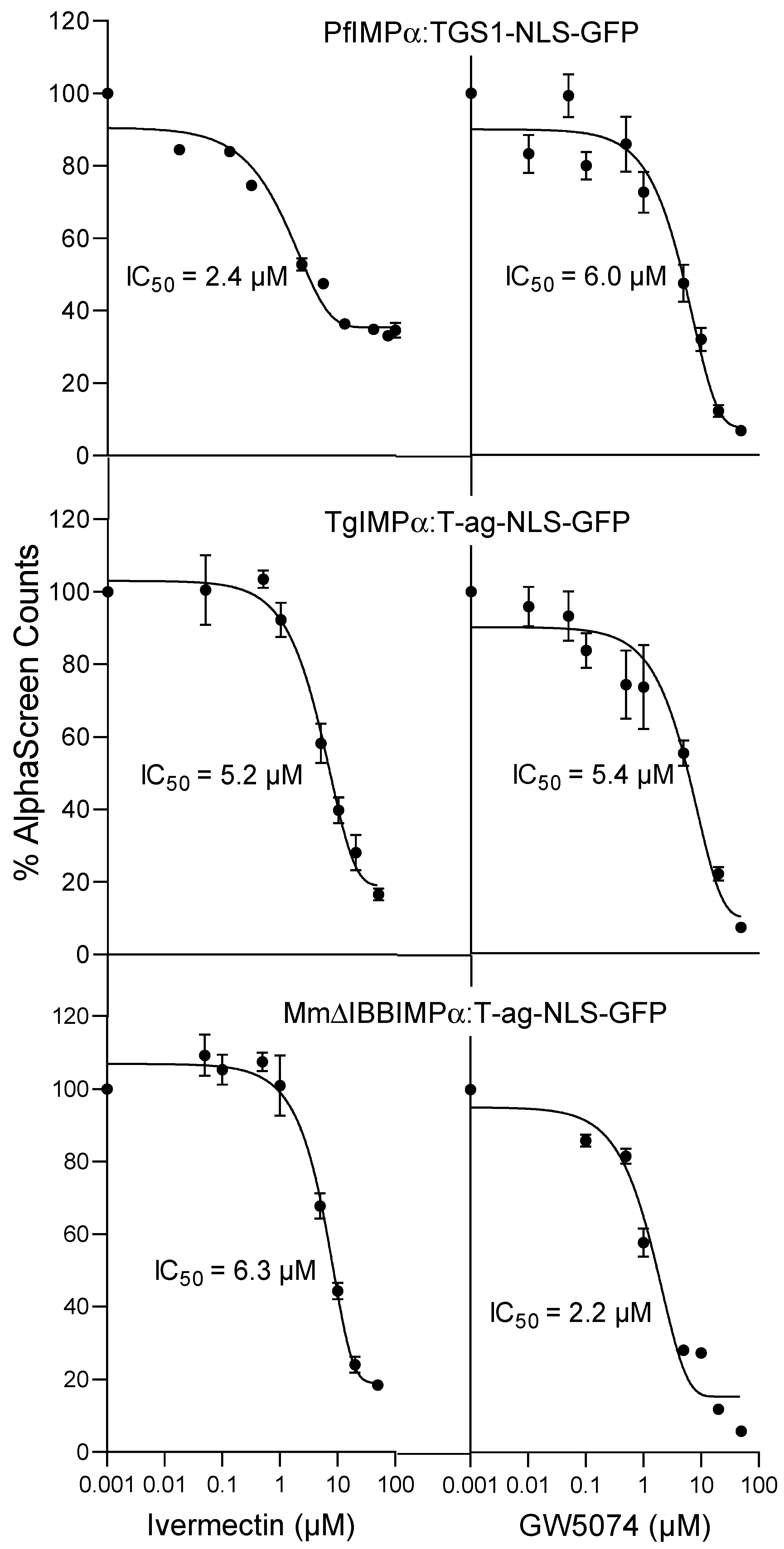
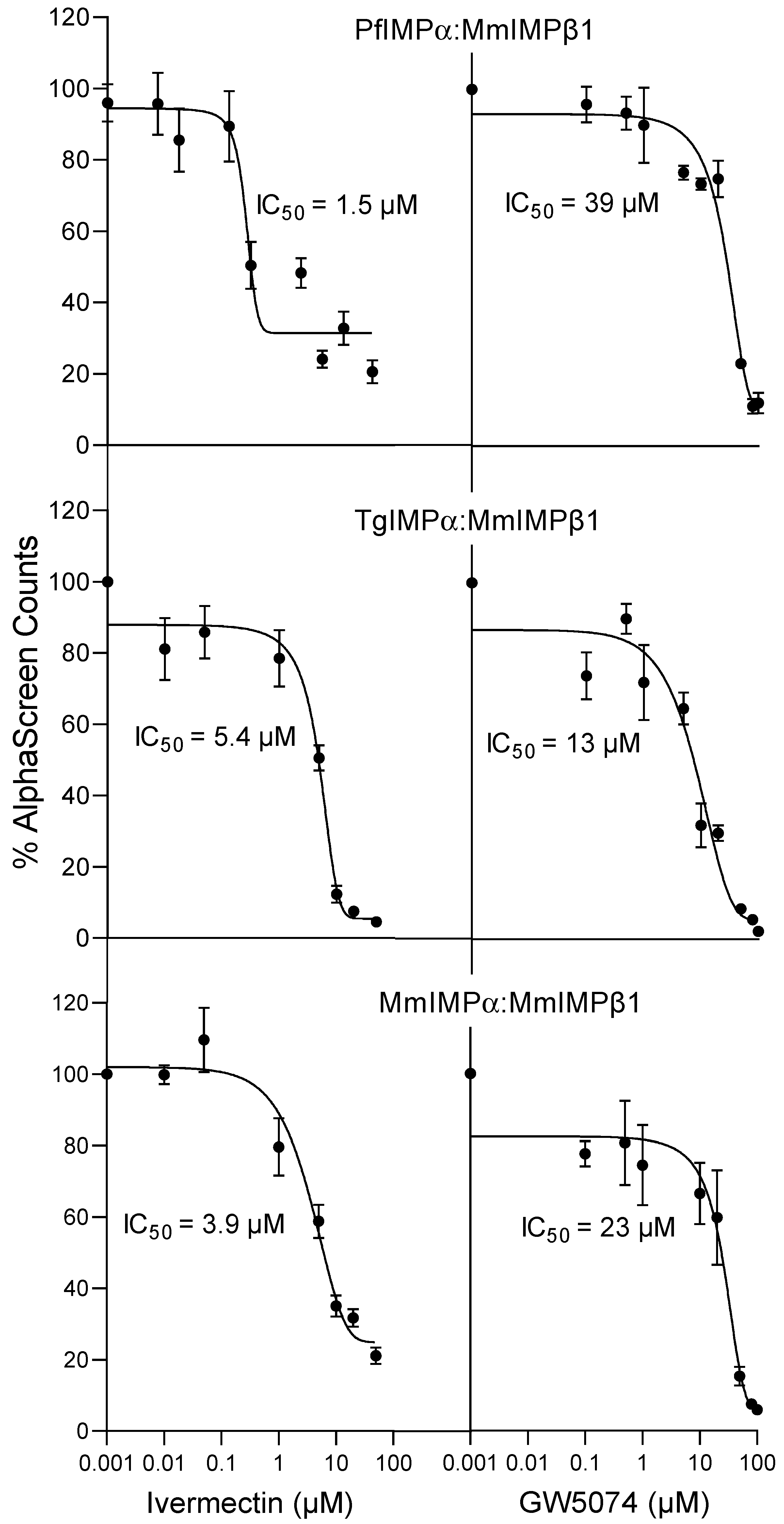
2.3. Inhibitors of Mammalian IMPα Can Block Apicomplexan IMPα Interaction with NLSs and MmIMPβ1
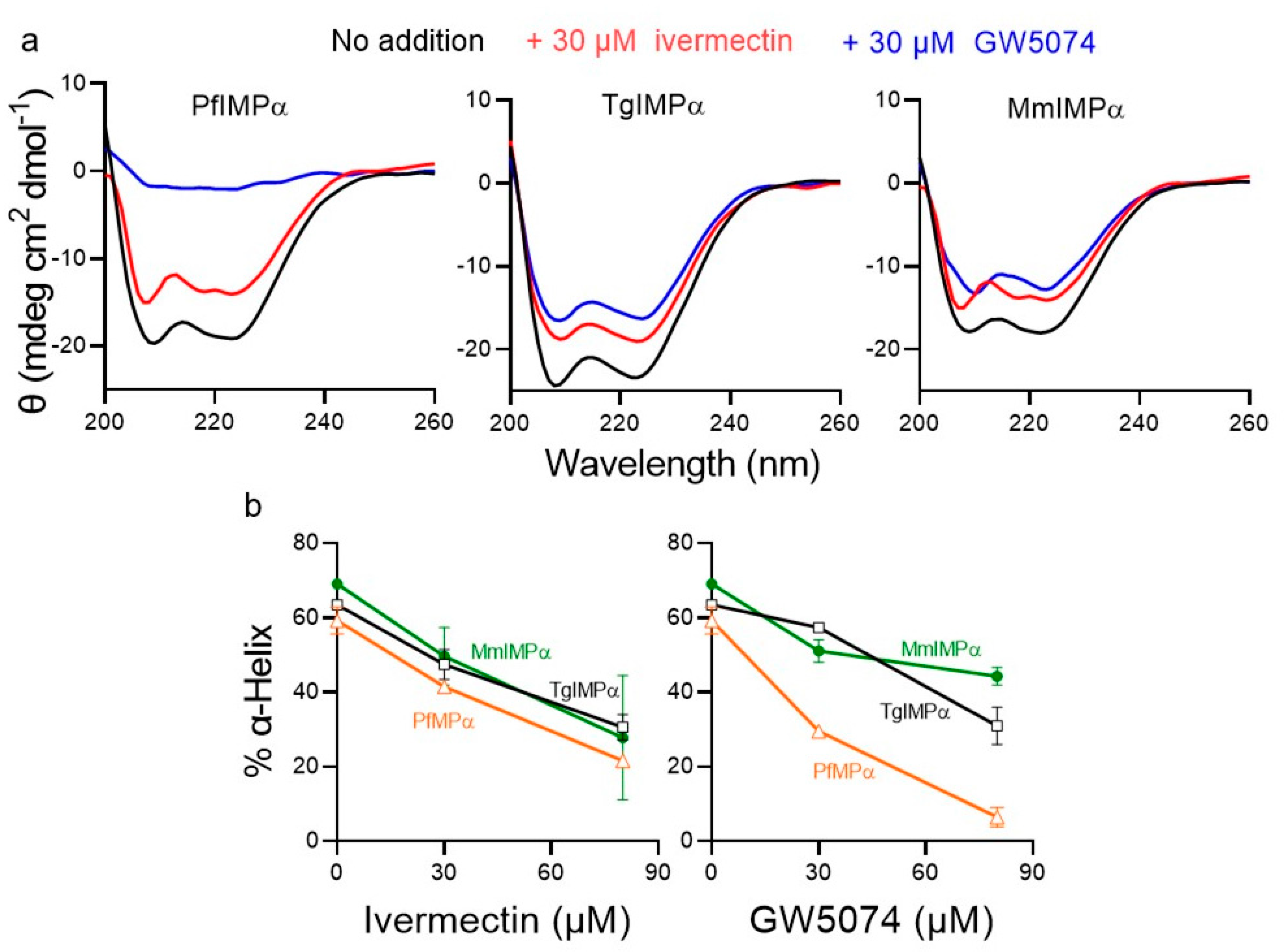
2.4. Ivermectin and GW5074 Appear to Bind Directly to Apicomplexan IMPαs to Have an Impact on Conformation

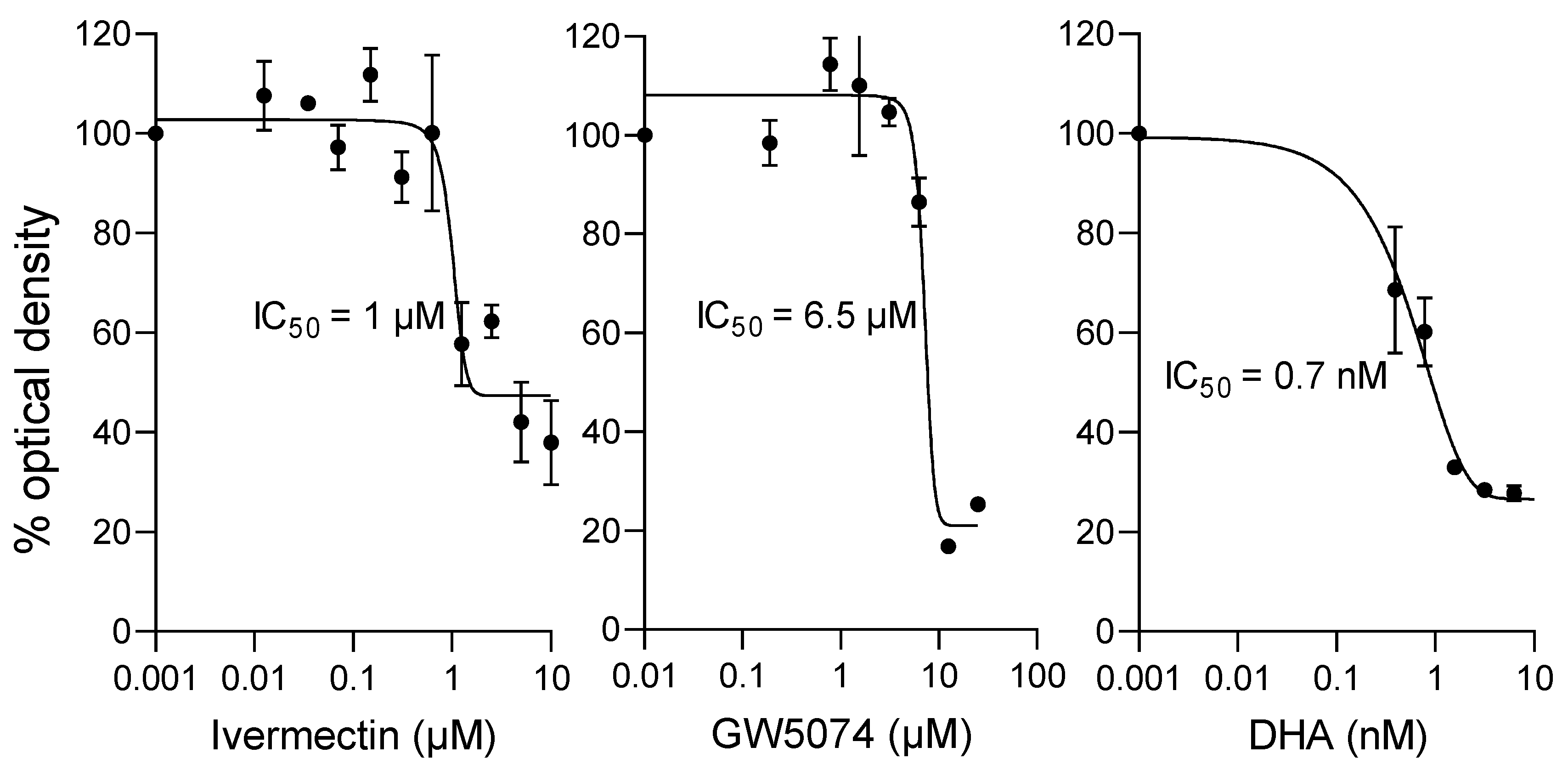
2.5. Ivermectin and GW5074 Can Limit the Proliferation of P. falciparum In Vitro
3. Discussion
4. Materials and Methods
4.1. Protein Expression, Purification, and Use in AlphaScreen Assay
4.2. Inhibitors
4.3. CD Spectroscopy
4.4. P. falciparum Culture and Growth Inhibition Assay
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Seeber, F.; Steinfelder, S. Recent Advances in Understanding Apicomplexan Parasites. F1000Research 2016, 5, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowman, A.F.; Healer, J.; Marapana, D.; Marsh, K. Malaria: Biology and Disease. Cell 2016, 167, 610–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Malaria Report 2021; World Health Organization: Geneva, Switzerland, 2021; Licence: CC BY-NC-SA 3.0 IGO.
- Dunay, I.R.; Gajurel, K.; Dhakal, R.; Liesenfeld, O.; Montoya, J.G. Treatment of toxoplasmosis: Historical perspective, animal models, and current clinical practice. Clin. Microbiol. Infect. 2018, 31, 1–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konstantinovic, N.; Guegan, H.; Stäjner, T.; Belaz, S.; Robert-Gangneux, F. Treatment of Toxoplasmosis: Current Options and Future Perspectives. Food Waterborne Parasitol. 2019, 15, e00036. [Google Scholar] [CrossRef]
- Zavala, F. RTS, S: The first malaria vaccine. J. Clin. Investig. 2022, 132, e156588. [Google Scholar] [CrossRef]
- Blasco, B.; Leroy, D.; Fidock, D.A. Antimalarial Drug Resistance: Linking Plasmodium falciparum Parasite Biology to the Clinic. Nat. Med. 2017, 23, 917–928. [Google Scholar] [CrossRef]
- Cui, L.; Mharakurwa, S.; Ndiaye, D.; Rathod, P.K.; Rosenthal, P.J. Antimalarial Drug Resistance: Literature Review and Activities and Findings of the ICEMR Network. Am. J. Trop. Med. Hyg. 2015, 93, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Montazeri, M.; Mehrzadi, S.; Sharif, M.; Sarvi, S.; Tanzifi, A.; Aghayan, S.A.; Daryani, A. Drug Resistance in Toxoplasma gondii. Front. Microbiol. 2018, 9, 2587. [Google Scholar] [CrossRef] [Green Version]
- Kosyna, F.K.; Depping, R. Controlling the Gatekeeper: Therapeutic Targeting of Nuclear Transport. Cells 2018, 7, 221. [Google Scholar] [CrossRef] [Green Version]
- Martin, A.J.; Jans, D.A. Antivirals That Target the Host IMPα/Β1-Virus Interface. Biochem. Soc. Trans. 2021, 49, 281–295. [Google Scholar] [CrossRef]
- Fulcher, A.J.; Jans, D.A. Regulation of Nucleocytoplasmic Trafficking of Viral Proteins: An Integral Role in Pathogenesis? Biochim. Biophys. Acta -Mol. Cell Res. 2011, 1813, 2176–2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jans, D.A.; Martin, A.J.; Wagstaff, K.M. Inhibitors of Nuclear Transport. Curr. Opin. Cell Biol. 2019, 58, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Goldfarb, D.S.; Corbett, A.H.; Mason, D.A.; Harreman, M.T.; Adam, S.A. Importin α: A Multipurpose Nuclear-Transport Receptor. Trends Cell Biol. 2004, 14, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Wente, S.R.; Rout, M.P. The Nuclear Pore Complex and Nuclear Transport. Cold Spring Harb. Perspect. Biol. 2010, 2, a000562. [Google Scholar] [CrossRef] [PubMed]
- Lott, K.; Bhardwaj, A.; Mitrousis, G.; Pante, N.; Cingolani, G. The Nuclear Pore Complex and Nuclear Transport. J. Biol. Chem. 2010, 285, 13769–13780. [Google Scholar] [CrossRef] [Green Version]
- Stewart, M. Molecular Mechanism of the Nuclear Protein Import Cycle. Nat. Rev. Mol. Cell Biol. 2007, 8, 195–208. [Google Scholar] [CrossRef]
- Loveland, K.L.; Major, A.T.; Butler, R.; Young, J.C.; Jans, D.A.; Miyamoto, Y. Putting Things in Place for Fertilization: Discovering Roles for Importin Proteins in Cell Fate and Spermatogenesis. Asian J. Androl. 2015, 17, 537. [Google Scholar] [CrossRef]
- Okada, N.; Ishigami, Y.; Suzuki, T.; Kaneko, A.; Yasui, K.; Fukutomi, R.; Isemura, M. Importins and Exportins in Cellular Differentiation. J. Cell. Mol. Med. 2008, 12, 1863–1871. [Google Scholar] [CrossRef] [Green Version]
- Wagstaff, K.M.; Rawlinson, S.M.; Hearps, A.C.; Jans, D.A. An AlphaScreen®-Based Assay for High-Throughput Screening for Specific Inhibitors of Nuclear Import. J. Biomol. Screen. 2011, 16, 192–200. [Google Scholar] [CrossRef] [Green Version]
- Shechter, S.; Thomas, D.R.; Lundberg, L.; Pinkham, C.; Lin, S.; Wagstaff, K.M.; Debono, A.; Kehn-hall, K.; Jans, D.A. Novel Inhibitors Targeting Venezuelan Equine Encephalitis Virus Capsid Protein Identified Using In Silico Structure-Based-Drug- Design. Sci. Rep. 2017, 7, 17705. [Google Scholar] [CrossRef]
- Thomas, D.R.; Lundberg, L.; Pinkham, C.; Shechter, S.; Debono, A.; Baell, J.; Wagstaff, K.M.; Hick, C.A.; Kehn-hall, K.; Jans, D.A. Identification of Novel Antivirals Inhibiting Recognition of Venezuelan Equine Encephalitis Virus Capsid Protein by the Importin α/β1 Heterodimer through High-Throughput Screening. Antivir. Res. 2018, 151, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Fraser, J.E.; Watanabe, S.; Wang, C.; Chan, W.K.K.; Maher, B.; Lopez-Denman, A.; Hick, C.; Wagstaff, K.M.; Mackenzie, J.M.; Sexton, P.M.; et al. A Nuclear Transport Inhibitor That Modulates the Unfolded Protein Response and Provides in Vivo Protection against Lethal Dengue Virus Infection. J. Infect. Dis. 2014, 210, 1780–1791. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Atkinson, S.; Fraser, J.; Wang, C.; Maher, B.; Roman, N.; Forwood, J.; Wagstaff, K.; Borg, N.; Jans, D. Novel Flavivirus Antiviral That Targets the Host Nuclear Transport Importin α/Β1 Heterodimer. Cells 2019, 8, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caly, L.; Druce, J.D.; Catton, M.G.; Jans, D.A.; Wagstaff, K.M. The FDA-Approved Drug Ivermectin Inhibits the Replication of SARS-CoV-2 in Vitro. Antivir. Res. 2020, 178, 104787. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, C.; Otto, T.D.; Oberstaller, J.; Liao, X.; Adapa, S.R.; Udenze, K.; Bronner, I.F.; Casandra, D.; Mayho, M.; et al. Uncovering the Essential Genes of the Human Malaria Parasite Plasmodium falciparum by Saturation Mutagenesis. Science 2018, 360, 521–526. [Google Scholar] [CrossRef] [Green Version]
- Sidik, S.M.; Huet, D.; Ganesan, S.M.; Huynh, M.H.; Wang, T.; Nasamu, A.S.; Thiru, P.; Saeij, J.P.J.; Carruthers, V.B.; Niles, J.C.; et al. A Genome-Wide CRISPR Screen in Toxoplasma Identifies Essential Apicomplexan Genes. Cell 2016, 166, 1423–1435.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, V.; Patankar, S. Molecular Basis for the Lack of Auto-Inhibition of Plasmodium falciparum Importin α. Biochem. Biophys. Res. Commun. 2018, 503, 1792–1797. [Google Scholar] [CrossRef]
- Babar, P.H.; Dey, V.; Jaiswar, P.; Patankar, S. An Insertion in the Methyltransferase Domain of P. falciparum Trimethylguanosine Synthase Harbors a Classical Nuclear Localization Signal. Mol. Biochem. Parasitol. 2016, 210, 58–70. [Google Scholar] [CrossRef]
- Bhatti, M.M.; Sullivan, W.J. Histone Acetylase GCN5 Enters the Nucleus via Importin-α in Protozoan Parasite Toxoplasma gondii. J. Biol. Chem. 2005, 280, 5902–5908. [Google Scholar] [CrossRef] [Green Version]
- Frankel, M.B.; Knoll, L.J. The Ins and Outs of Nuclear Trafficking: Unusual Aspects in Apicomplexan Parasites. DNA Cell Biol. 2009, 28, 277–284. [Google Scholar] [CrossRef]
- Yang, S.N.Y.; Atkinson, S.C.; Wang, C.; Lee, A.; Bogoyevitch, M.A.; Borg, N.A.; Jans, D.A. The Broad Spectrum Antiviral Ivermectin Targets the Host Nuclear Transport Importin α/Β1 Heterodimer. Antivir. Res. 2020, 177, 104760. [Google Scholar] [CrossRef] [PubMed]
- Wagstaff, K.M.; Sivakumaran, H.; Heaton, S.M.; Harrich, D.; Jans, D.A. Ivermectin Is a Specific Inhibitor of Importin α/β-Mediated Nuclear Import Able to Inhibit Replication of HIV-1 and Dengue Virus. Biochem. J. 2012, 443, 851–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panchal, M.; Rawat, K.; Kumar, G.; Kibria, K.M.; Singh, S.; Kalamuddin, M.; Mohmmed, A.; Malhotra, P.; Tuteja, R. Plasmodium falciparum Signal Recognition Particle Components and Anti-Parasitic Effect of Ivermectin in Blocking Nucleo-Cytoplasmic Shuttling of SRP. Cell Death Dis. 2014, 5, e994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobe, B. Autoinhibition by an Internal Nuclear Localization Signal Revealed by the Crystal Structure of Mammalian Importin α. Nat. Struct. Biol. 1999, 6, 388–397. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, L.P.; Sandri, T.L.; Tenório de Melo, E.J.; Fendel, R.; Kremsner, P.G.; Mordmüller, B.; Held, J. Ivermectin Impairs the Development of Sexual and Asexual Stages of Plasmodium falciparum in Vitro. Antimicrob. Agents Chemother. 2019, 63, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Slater, H.C.; Foy, B.D.; Kobylinski, K.; Chaccour, C.; Watson, O.J.; Hellewell, J.; Aljayyoussi, G.; Bousema, T.; Burrows, J.; D’Alessandro, U.; et al. Ivermectin as a Novel Complementary Malaria Control Tool to Reduce Incidence and Prevalence: A Modelling Study. Lancet Infect. Dis. 2020, 20, 498–508. [Google Scholar] [CrossRef]
- Kurth, F.; Pongratz, P.; Bélard, S.; Mordmüller, B.; Kremsner, P.G.; Ramharter, M. In Vitro Activity of Pyronaridine against Plasmodium falciparum and Comparative Evaluation of Anti-Malarial Drug Susceptibility Assays. Malar. J. 2009, 8, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Noedl, H.; Wernsdorfer, W.H.; Miller, R.S.; Wongsrichanalai, C. Histidine-Rich Protein II: A Novel Approach to Malaria Drug Sensitivity Testing. Antimicrob. Agents Chemother. 2002, 46, 1658–1664. [Google Scholar] [CrossRef] [Green Version]
- Noedl, H.; Bronnert, J.; Yingyuen, K.; Attlmayr, B.; Kollaritsch, H.; Fukuda, M. Simple Histidine-Rich Protein 2 Double-Site Sandwich Enzyme-Linked Immunosorbent Assay for Use in Malaria Drug Sensitivity Testing. Antimicrob. Agents Chemother. 2005, 49, 3575–3577. [Google Scholar] [CrossRef] [Green Version]
- Cautain, B.; Hill, R.; de Pedro, N.; Link, W. Components and Regulation of Nuclear Transport Processes. FEBS J. 2015, 282, 445–462. [Google Scholar] [CrossRef]
- Devos, D.P.; Gräf, R.; Field, M.C. Evolution of the Nucleus. Curr. Opin. Cell Biol. 2014, 28, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krissinel, E. On the Relationship between Sequence and Structure Similarities in Proteomics. Bioinformatics 2007, 23, 717–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.; Bogoyevitch, M.A.; Jans, D.A. Bimolecular Fluorescence Complementation: Quantitative Analysis of In Cell Interaction of Nuclear Transporter Importin α with Cargo Proteins. Methods Mol. Biol. 2022, 250, 215–233. [Google Scholar] [CrossRef]
- Walunj, S.B.; Dias, M.M.; Kaur, C.; Wagstaff, K.M.; Dey, V.; Hick, C.; Patankar, S.; Jans, D.A. High-Throughput Screening to Identify Inhibitors of Plasmodium falciparum Importin alpha. Cells 2022, 11, 1201. [Google Scholar] [CrossRef] [PubMed]
- Crump, A.; Omura, S. Ivermectin, wonder drug from Japan: The human use perspective. Proc. Jpn. Acad. 2011, 87, 13–28. [Google Scholar] [CrossRef] [Green Version]
- González Canga, A.; Sahagún Prieto, A.M.; Diez Liébana, M.J.; Fernández Martínez, N.; Sierra Vega, M.; García Vieitez, J.J. The pharmacokinetics and interactions of ivermectin in humans—A mini-review. AAPSJ 2008, 10, 42–46. [Google Scholar] [CrossRef] [Green Version]
- Ooi, E.E. Repurposing Ivermectin as an Anti-Dengue Drug. Clin. Infect. Dis. 2021, 72, e594–e595. [Google Scholar] [CrossRef]
- Mastrangelo, E.; Pezzullo, M.; De Burghgraeve, T.; Kaptein, S.; Pastorino, B.; Dallmeier, K.; de Lamballerie, X.; Neyts, J.; Hanson, A.M.; Frick, D.N.; et al. Ivermectin is a potent inhibitor of flavivirus replication specifically targeting NS3 helicase activity: New prospects for an old drug. J. Antimicrob. Chemother. 2012, 67, 1884–1894. [Google Scholar] [CrossRef] [Green Version]
- Jans, D.A.; Wagstaff, K.M. Ivermectin as a Broad-Spectrum Host-Directed Antiviral: The Real Deal? Cells 2020, 9, 2100. [Google Scholar] [CrossRef]
- Bryant, A.; Lawrie, T.A.; Dowswell, T.; Fordham, E.J.; Mitchell, S.; Hill, S.R.; Tham, T.C. Ivermectin for Prevention and Treatment of COVID-19 Infection: A Systematic Review, Meta-Analysis, and Trial Sequential Analysis to Inform Clinical Guidelines. Am. J. Ther. 2021, 28, e434–e460. [Google Scholar] [CrossRef]
- Foy, B.D.; Alout, H.; Seaman, J.A.; Rao, S.; Magalhaes, T.; Wade, M.; Parikh, S.; Soma, D.; Sagna, B.; Fournet, F.; et al. Efficacy and Risk of Harms of Repeat Ivermectin Mass Drug Administrations for Control of Malaria (RIMDAMAL): A Cluster-Randomised Trial. Lancet 2019, 393, 1517–1526. [Google Scholar] [CrossRef] [Green Version]
- Ouédraogo, A.L.; Bastiaens, G.J.H.; Tiono, A.B.; Guelbéogo, W.M.; Kobylinski, K.C.; Ouédraogo, A.; Barry, A.; Bougouma, E.C.; Nebie, I.; Ouattara, M.S.; et al. Efficacy and Safety of the Mosquitocidal Drug Ivermectin to Prevent Malaria Transmission after Treatment: A Double-Blind, Randomized, Clinical Trial. Clin. Infect. Dis. 2015, 60, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Oshokoya, O.O.; Roach, C.A.; Jiji, R.D. Quantification of Protein Secondary Structure Content by Multivariate Analysis of Deep-Ultraviolet Resonance Raman and Circular Dichroism Spectroscopies. Anal. Methods 2014, 6, 1691–1699. [Google Scholar] [CrossRef]
- Trager, W.; Jensen, J.B. Human Malaria Parasites in Continuous Culture. Science 1976, 193, 673–675. [Google Scholar] [CrossRef]
- Radfar, A.; Méndez, D.; Moneriz, C.; Linares, M.; Marín-García, P.; Puyet, A.; Diez, A.; Bautista, J.M. Synchronous Culture of Plasmodium falciparum at High Parasitemia Levels. Nat. Protoc. 2009, 4, 1899–1915. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Binding Interaction | Kd (nM) * |
|---|---|
| PfIMPα:T-ag-NLS-GFP | 2.9 ± 1.6 |
| PfIMPα:TGS1-NLS-GFP | 6.6 ± 0.3 |
| PfIMPα:GCN5-NLS-GFP | 2.5 ± 1.7 |
| TgIMPα:T-ag-NLS-GFP | 3.9 ± 1.1 |
| TgIMPα:GCN5-NLS-GFP | 5.0 ± 1.5 |
| TgIMPα:TGS1-NLS-GFP | 1.9 ± 0.7 |
| MmIMPα/β1:T-ag-NLS-GFP | 2.8 ± 0.2 |
| MmIMPα/β1:TGS1-NLS-GFP | 2.5 ± 0.1 |
| MmIMPα/β1:GCN5-NLS-GFP | 3.2 ± 0.2 |
| Binding Interaction | Kd (nM) * |
|---|---|
| PfIMPα:MmIMPβ1 | 1.3 ± 0.3 |
| TgIMPα:MmIMPβ1 | 1.2 ± 0.4 |
| MmIMPα:MmIMPβ1 | 2.5 ± 0.2 |
| IC50 (μM) * | ||
|---|---|---|
| Binding Interaction | Ivermectin | GW5074 |
| PfIMPα:TGS1 | 5.0 ± 1.3 | 6.5 ± 0.4 |
| TgIMPα:SV40 T-ag | 4.9 ± 0.2 | 7.7 ± 1.9 |
| ΔIBBmIMPα:SV40 T-ag | 6.2 ± 0.2 | 4.9 ± 1.4 |
| IC50 (µM) * | ||
|---|---|---|
| Binding Interaction | Ivermectin | GW5074 |
| PfIMPα:MmIMPβ1 | 1.9 ± 0.2 | 41 ± 1.5 |
| TgIMPα:MmIMPβ1 | 6.9 ± 1.2 | 11 ± 1.5 |
| MmIMPα:MmIMPβ1 | 3.1 ± 0.5 | 22 ± 0.6 |
| Compound | IC50 (µM) * |
|---|---|
| Ivermectin | 0.7 ± 0.1 |
| GW5074 | 5.5 ± 0.9 |
| DHA | 0.001 ± 0.0005 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walunj, S.B.; Wang, C.; Wagstaff, K.M.; Patankar, S.; Jans, D.A. Conservation of Importin α Function in Apicomplexans: Ivermectin and GW5074 Target Plasmodium falciparum Importin α and Inhibit Parasite Growth in Culture. Int. J. Mol. Sci. 2022, 23, 13899. https://doi.org/10.3390/ijms232213899
Walunj SB, Wang C, Wagstaff KM, Patankar S, Jans DA. Conservation of Importin α Function in Apicomplexans: Ivermectin and GW5074 Target Plasmodium falciparum Importin α and Inhibit Parasite Growth in Culture. International Journal of Molecular Sciences. 2022; 23(22):13899. https://doi.org/10.3390/ijms232213899
Chicago/Turabian StyleWalunj, Sujata B., Chunxiao Wang, Kylie M. Wagstaff, Swati Patankar, and David A. Jans. 2022. "Conservation of Importin α Function in Apicomplexans: Ivermectin and GW5074 Target Plasmodium falciparum Importin α and Inhibit Parasite Growth in Culture" International Journal of Molecular Sciences 23, no. 22: 13899. https://doi.org/10.3390/ijms232213899