
Anticarcinogenic Effects of Isothiocyanates on Hepatocellular Carcinoma


Abstract
:1. Introduction
2. Aetiology and Pathophysiology of HCC
2.1. Risk Factors for HCC
2.2. Pathophysiology
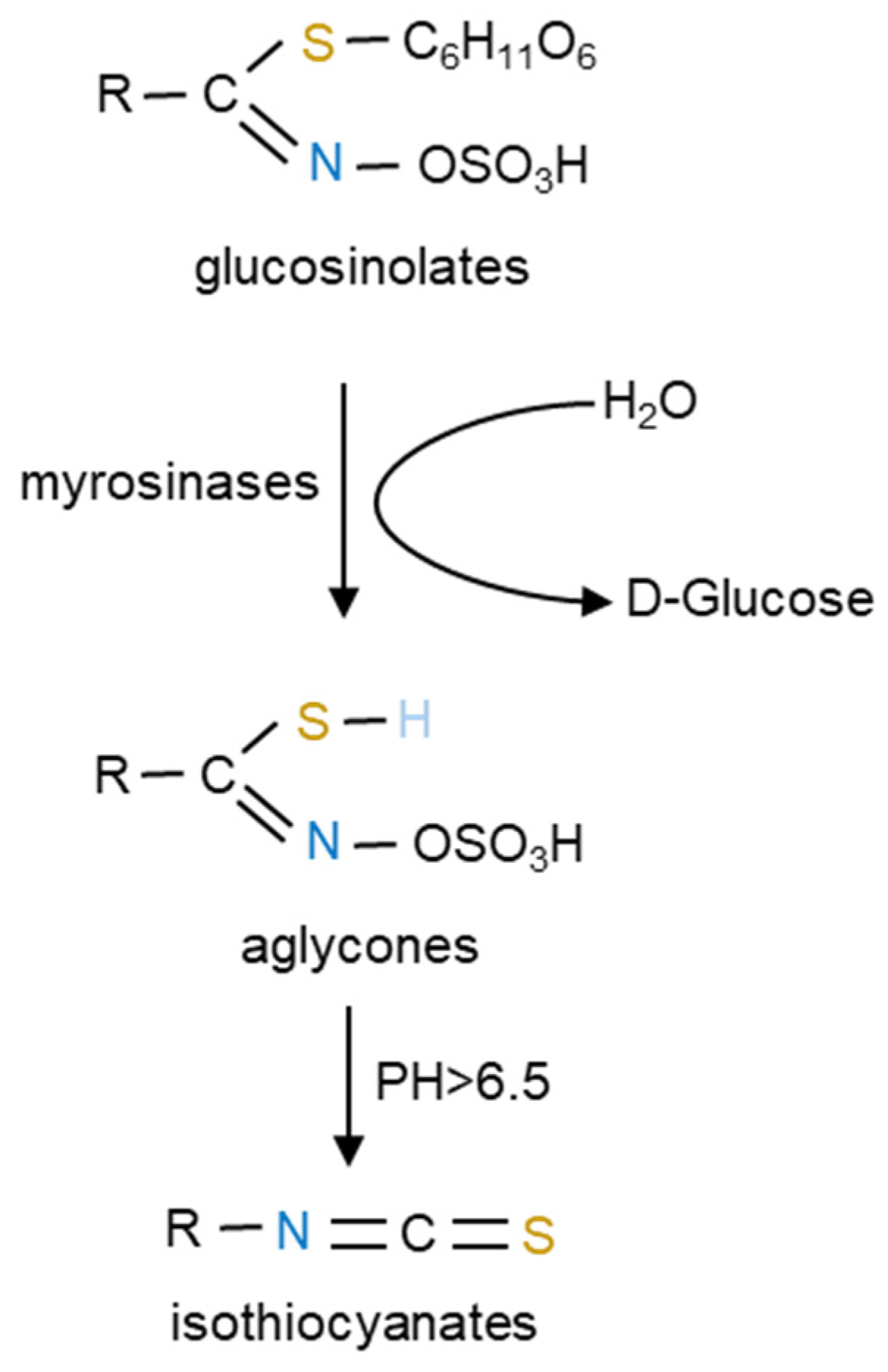
3. GLSs and Their Derived ITCs
4. The Role of ITCs as Chemopreventive Agents on HCC
4.1. ITCs Inhibit the Activation and Enhance the Detoxification of Carcinogens by Modulation of Phase I and Phase II Enzymes
4.2. Chemopreventive Activity of ITCs on HCC through Epigenetic Regulation
4.2.1. The Effect of ITCs on Post-Translational Histone Modification and DNA Methylation in HCC
4.2.2. The Ability of ITCs to Alter miRNA Expression in HCC
5. Anticancer Effects and Molecular Mechanisms of ITCs on HCC
5.1. Inhibition of the Proliferation of HCC Cells
5.2. Arrest of HCC Cell Cycle
5.3. Inducing Apoptosis of HCC Cells
5.3.1. Apoptosis Process Mediated by Mitochondria
5.3.2. Apoptotic Pathway Induced by Endoplasmic Reticulum Stress
5.3.3. Apoptosis Process Mediated by Death Receptors
5.4. Inhibition of Tumor Cell Migration
5.5. Inhibition of Tumor Cell Angiogenesis
5.6. Decrease in Telomerase Activity of Tumor Cells
6. Sensitization to Chemotherapeutic Agents or Radiation Therapy by ITC Pre- or Co-Treatment
7. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Llovet, J.M.; Zucman Rossi, J.; Pikarsky, E.; Sangro, B.; Schwartz, M.; Sherman, M.; Gores, G. Hepatocellular carcinoma. Nat. Rev. Dis. Prim. 2016, 2, 16018. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.S.; Qin, S.; Ikeda, M.; Galle, P.R.; Ducreux, M.; Kim, T.Y.; Kudo, M.; Breder, V.; Merle, P.; Kaseb, A.O.; et al. Atezolizumab plus bevacizumab in unresectable hepatocellular carcinoma. N. Engl. J. Med. 2020, 382, 1894–1905. [Google Scholar] [CrossRef]
- Llovet, J.M.; Villanueva, A.; Marrero, J.A.; Schwartz, M.; Meyer, T.; Galle, P.R.; Lencioni, R.; Greten, T.F.; Kudo, M.; Mandrekar, S.J.; et al. Trial design and endpoints in hepatocellular carcinoma: AASLD consensus conference. Hepatology 2021, 73 (Suppl. 1), 158–191. [Google Scholar] [CrossRef]
- Llovet, J.M.; Ricci, S.; Mazzaferro, V.; Hilgard, P.; Gane, E.; Blanc, J.F.; de Oliveira, A.C.; Santoro, A.; Raoul, J.L.; Forner, A.; et al. Sorafenib in advanced hepatocellular carcinoma. N. Engl. J. Med. 2008, 359, 378–390. [Google Scholar] [CrossRef] [Green Version]
- Sato, J.; Satouchi, M.; Itoh, S.; Okuma, Y.; Niho, S.; Mizugaki, H.; Murakami, H.; Fujisaka, Y.; Kozuki, T.; Nakamura, K.; et al. Lenvatinib in patients with advanced or metastatic thymic carcinoma (REMORA): A multicentre, phase 2 trial. Lancet Oncol. 2020, 21, 843–850. [Google Scholar] [CrossRef]
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Zucman-Rossi, J.; Finn, R.S. Hepatocellular carcinoma. Nat. Rev. Dis. Prim. 2021, 7, 6. [Google Scholar] [CrossRef]
- Fontana, F.; Raimondi, M.; Marzagalli, M.; Di Domizio, A.; Limonta, P. The emerging role of paraptosis in tumor cell biology: Perspectives for cancer prevention and therapy with natural compounds. Biochim. Biophys. Acta Rev. Cancer 2020, 1873, 188338. [Google Scholar] [CrossRef]
- Kubczak, M.; Szustka, A.; Rogalińska, M. Molecular targets of natural compounds with anti-cancer properties. Int. J. Mol. Sci. 2021, 22, 13659. [Google Scholar] [CrossRef]
- Tewari, D.; Patni, P.; Bishayee, A.; Sah, A.N.; Bishayee, A. Natural products targeting the PI3K-AKT-mTOR signaling pathway in cancer: A novel therapeutic strategy. Semin. Cancer Biol. 2022, 80, 1–17. [Google Scholar] [CrossRef]
- Thomford, N.; Senthebane, D.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural products for drug discovery in the 21st century: Innovations for novel drug discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef] [Green Version]
- Nambiar, D.M.; Kumari, J.; Arya, G.C.; Singh, A.K.; Bisht, N.C. A cell suspension based uptake method to study high affinity glucosinolate transporters. Plant. Methods 2020, 16, 75. [Google Scholar] [CrossRef]
- Bones, A.M.; Rossiter, J.T. The enzymic and chemically induced decomposition of glucosinolates. Phytochemistry 2006, 67, 1053–1067. [Google Scholar] [CrossRef]
- Gu, H.F.; Mao, X.Y.; Du, M. Metabolism, absorption, and anti-cancer effects of sulforaphane: An update. Crit. Rev. Food Sci. Nutr. 2021, 62, 3437–3452. [Google Scholar] [CrossRef]
- Mastuo, T.; Miyata, Y.; Yuno, T.; Mukae, Y.; Otsubo, A.; Mitsunari, K.; Ohba, K.; Sakai, H. Molecular mechanisms of the anti-cancer effects of isothiocyanates from cruciferous Vegetables in bladder cancer. Molecules 2020, 25, E575. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Arora, R.; Bhatia, A.; Singh, H.; Singh, B.; Arora, S. Molecular targets in cancer prevention by 4-(methylthio)butyl isothiocyanate—A comprehensive review. Life Sci. 2020, 241, 117061. [Google Scholar] [CrossRef]
- Boldry, E.J.; Yuan, J.M.; Carmella, S.G.; Wang, R.; Tessier, K.; Hatsukami, D.K.; Hecht, S.S.; Tretyakova, N.Y. Effects of 2-phenethyl isothiocyanate on metabolism of 1,3-butadiene in smokers. Cancer Prev. Res. 2020, 13, 91–100. [Google Scholar] [CrossRef]
- Rekha, K.; Venkidasamy, B.; Govindasamy, R.; Neralla, M.; Thiruvengadam, M. Isothiocyanates (AITC & BITC) bioactive molecules: Therapeutic potential for oral cancer. Oral. Oncol. 2022, 133, 106060. [Google Scholar] [CrossRef]
- Rakariyatham, K.; Yang, X.; Gao, Z.; Song, M.; Han, Y.; Chen, X.; Xiao, H. Synergistic chemopreventive effect of allyl isothiocyanate and sulforaphane on non-small cell lung carcinoma cells. Food Funct. 2019, 10, 893–902. [Google Scholar] [CrossRef]
- Galle, P.R.; Forner, A.; Llovet, J.M. EASL clinical practice guidelines: Management of hepatocellular carcinoma. J. Hepatol. 2018, 69, 182–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrero, J.A.; Kulik, L.M.; Sirlin, C.B.; Zhu, A.X.; Finn, R.S.; Abecassis, M.M.; Roberts, L.R.; Heimbach, J.K. Diagnosis, staging, and management of hepatocellular carcinoma: 2018 practice guidance by the american association for the study of liver diseases. Hepatology 2018, 68, 723–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akinyemiju, T.; Abera, S.; Ahmed, M.; Alam, N.; Alemayohu, M.A.; Allen, C.; Al Raddadi, R.; Alvis Guzman, N.; Amoako, Y.; Artaman, A.; et al. The burden of primary liver cancer and underlying etiologies from 1990 to 2015 at the global, regional, and national level: Results from the global burden of disease study 2015. JAMA Oncol. 2017, 3, 1683–1691. [Google Scholar] [CrossRef] [PubMed]
- Estes, C.; Razavi, H.; Loomba, R.; Younossi, Z.; Sanyal, A.J. Modeling the epidemic of nonalcoholic fatty liver disease demonstrates an exponential increase in burden of disease. Hepatology 2018, 67, 123–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.W.; Lin, C.C.; Mo, L.R.; Chang, C.Y.; Perng, D.S.; Hsu, C.C.; Lo, G.H.; Chen, Y.S.; Yen, Y.C.; Hu, J.T.; et al. Heavy alcohol consumption increases the incidence of hepatocellular carcinoma in hepatitis B virus-related cirrhosis. J. Hepatol. 2013, 58, 730–735. [Google Scholar] [CrossRef]
- Trinchet, J.C.; Bourcier, V.; Chaffaut, C.; Ait Ahmed, M.; Allam, S.; Marcellin, P.; Guyader, D.; Pol, S.; Larrey, D.; De Lédinghen, V.; et al. Complications and competing risks of death in compensated viral cirrhosis (ANRS CO12 CirVir prospective cohort). Hepatology 2015, 62, 737–750. [Google Scholar] [CrossRef]
- Yang, J.D.; Roberts, L.R. Hepatocellular carcinoma: A global view. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 448–458. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Chenivesse, X.; Henglein, B.; Bréchot, C. Hepatitis B virus integration in a cyclin A gene in a hepatocellular carcinoma. Nature 1990, 343, 555–557. [Google Scholar] [CrossRef]
- Kew, M.C. Synergistic interaction between aflatoxin B1 and hepatitis B virus in hepatocarcinogenesis. Liver Int. 2003, 23, 405–409. [Google Scholar] [CrossRef]
- Yang, J.D.; Gyedu, A.; Afihene, M.Y.; Duduyemi, B.M.; Micah, E.; Kingham, T.P.; Nyirenda, M.; Nkansah, A.A.; Bandoh, S.; Duguru, M.J.; et al. Hepatocellular carcinoma occurs at an earlier age in africans, particularly in association with chronic hepatitis B. Am. J. Gastroenterol. 2015, 110, 1629–1631. [Google Scholar] [CrossRef]
- De Villiers, M.J.; Nayagam, S.; Hallett, T.B. The impact of the timely birth dose vaccine on the global elimination of hepatitis B. Nat. Commun. 2021, 12, 6223. [Google Scholar] [CrossRef] [PubMed]
- Trucchi, C.; Orsi, A.; Alicino, C.; Sticchi, L.; Icardi, G.; Ansaldi, F. State of the art, unresolved issues, and future research directions in the fight against hepatitis C virus: Perspectives for screening, diagnostics of resistances, and immunization. J. Immunol. Res. 2016, 2016, 1412840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rich, N.E.; Hester, C.; Odewole, M.; Murphy, C.C.; Parikh, N.D.; Marrero, J.A.; Yopp, A.C.; Singal, A.G. Racial and ethnic differences in presentation and outcomes of hepatocellular carcinoma. Clin. Gastroenterol. Hepatol. 2019, 17, 551–559. [Google Scholar] [CrossRef] [Green Version]
- Rich, N.E.; Yopp, A.C.; Singal, A.G.; Murphy, C.C. Hepatocellular carcinoma incidence is decreasing among younger adults in the United States. Clin. Gastroenterol. Hepatol. 2020, 18, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Forner, A.; Reig, M.; Bruix, J. Hepatocellular carcinoma. Lancet 2018, 391, 1301–1314. [Google Scholar] [CrossRef]
- Farazi, P.A.; DePinho, R.A. Hepatocellular carcinoma pathogenesis: From genes to environment. Nat. Rev. Cancer 2006, 6, 674–687. [Google Scholar] [CrossRef]
- Zucman Rossi, J.; Villanueva, A.; Nault, J.C.; Llovet, J.M. Genetic landscape and biomarkers of hepatocellular carcinoma. Gastroenterology 2015, 149, 1226–1239.e4. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.D.; Hainaut, P.; Gores, G.J.; Amadou, A.; Plymoth, A.; Roberts, L.R. A global view of hepatocellular carcinoma: Trends, risk, prevention and management. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 589–604. [Google Scholar] [CrossRef]
- Liao, C.L.; Peng, S.F.; Chen, J.C.; Chen, P.Y.; Huang, A.C.; Lien, J.C.; Chueh, F.S.; Chiang, T.A.; Wu, P.P.; Lin, K.I. Allyl isothiocyanate induces DNA damage and impairs DNA repair in human breast cancer MCF-7 cells. Anticancer Res. 2021, 41, 4343–4351. [Google Scholar] [CrossRef]
- Mitsiogianni, M.; Anestopoulos, I.; Kyriakou, S.; Trafalis, D.T.; Franco, R.; Pappa, A.; Panayiotidis, M.I. Benzyl and phenethyl isothiocyanates as promising epigenetic drug compounds by modulating histone acetylation and methylation marks in malignant melanoma. Investig. New Drugs 2021, 39, 1460–1468. [Google Scholar] [CrossRef] [PubMed]
- Ngo, S.N.T.; Williams, D.B. Protective effect of isothiocyanates from cruciferous vegetables on breast cancer: Epidemiological and preclinical perspectives. Anticancer Agents Med. Chem. 2021, 21, 1413–1430. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.F.; Hao, Y.Y.; Gao, S.; Li, X.Q.; Liu, F.-H.; Wen, Z.Y.; Wang, H.Y.; Zhang, S.; Yan, S.; Luan, M.; et al. Pre-diagnosis cruciferous vegetables and isothiocyanates intake and ovarian cancer survival: A prospective cohort study. Front. Nutr. 2021, 8, 778031. [Google Scholar] [CrossRef]
- Novío, S.; Cartea, M.; Soengas, P.; Freire Garabal, M.; Núñez Iglesias, M. Effects of brassicaceae isothiocyanates on prostate cancer. Molecules 2016, 21, 626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wittstock, U.; Halkier, B.A. Glucosinolate research in the Arabidopsis era. Trends Plant. Sci. 2002, 7, 263–270. [Google Scholar] [CrossRef]
- Agerbirk, N.; Olsen, C.E. Glucosinolate structures in evolution. Phytochemistry 2012, 77, 16–45. [Google Scholar] [CrossRef]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Karcher, A.; El Rassi, Z. Capillary electrophoresis of glucosinolates and their degradation products. Electrophoresis 1999, 20, 3181–3189. [Google Scholar] [CrossRef]
- Vig, A.P.; Rampal, G.; Thind, T.S.; Arora, S. Bio-protective effects of glucosinolates—A review. LWT-Food Sci. Technol. 2009, 42, 1561–1572. [Google Scholar] [CrossRef]
- Thangstad, O.P.; Gilde, B.; Chadchawan, S.; Seem, M.; Husebye, H.; Bradley, D.; Bones, A.M. Cell specific, cross-species expression of myrosinases in brassica napus, arabidopsis thaliana and nicotiana tabacum. Plant Mol. Biol. 2004, 54, 597–611. [Google Scholar] [CrossRef]
- Dinkova Kostova, A.T. Chemoprotection against cancer by isothiocyanates: A focus on the animal models and the protective mechanisms. Top. Curr. Chem. 2012, 329, 179–201. [Google Scholar] [CrossRef]
- Murata, M.; Yamashita, N.; Inoue, S.; Kawanishi, S. Mechanism of oxidative DNA damage induced by carcinogenic allyl isothiocyanate. Free Radic. Biol. Med. 2000, 28, 797–805. [Google Scholar] [CrossRef]
- Zhang, Y.; Yao, S.; Li, J. Vegetable-derived isothiocyanates: Anti-proliferative activity and mechanism of action. Proc. Nutr. Soc. 2006, 65, 68–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, W.T.; Yen, C.C.; Huang, C.S.; Chen, H.W.; Lii, C.K. Benzyl isothiocyanate ameliorates high-fat diet-induced hyperglycemia by enhancing Nrf2-dependent antioxidant defense-mediated IRS-1/AKT/TBC1D1 signaling and GLUT4 expression in skeletal muscle. J. Agric. Food Chem. 2020, 68, 15228–15238. [Google Scholar] [CrossRef]
- Dixit, S.; Widemann, E.; Bensoussan, N.; Salehipourshirazi, G.; Bruinsma, K.; Milojevic, M.; Shukla, A.; Romero, L.C.; Zhurov, V.; Bernards, M.A.; et al. β-Cyanoalanine synthase protects mites against arabidopsis defenses. Plant. Physiol. 2022, 189, 1961–1975. [Google Scholar] [CrossRef]
- Jain, M.K.; Rich, N.E.; Ahn, C.; Turner, B.J.; Sanders, J.M.; Adamson, B.; Quirk, L.; Perryman, P.; Santini, N.O.; Singal, A.G. Evaluation of a multifaceted intervention to reduce health disparities in hepatitis C screening: A pre-post analysis. Hepatology 2019, 70, 40–50. [Google Scholar] [CrossRef]
- Li, P.; Zhao, Y.; Wang, C.; Zhu, H. Antibacterial activity and main action pathway of benzyl isothiocyanate extracted from papaya seeds. J. Food Sci. 2021, 86, 169–176. [Google Scholar] [CrossRef]
- Lohning, A.; Kidachi, Y.; Kamiie, K.; Sasaki, K.; Ryoyama, K.; Yamaguchi, H. 6-(methylsulfinyl)hexyl isothiocyanate (6-MITC) from Wasabia japonica alleviates inflammatory bowel disease (IBD) by potential inhibition of glycogen synthase kinase 3 beta (GSK-3β). Eur. J. Med. Chem. 2021, 216, 113250. [Google Scholar] [CrossRef]
- Justin, S.; Rutz, J.; Maxeiner, S.; Chun, F.K.H.; Juengel, E.; Blaheta, R.A. Bladder cancer metastasis induced by chronic everolimus application can be counteracted by sulforaphane in vitro. Int. J. Mol. Sci. 2020, 21, E5582. [Google Scholar] [CrossRef]
- Nguyen, Y.T.K.; Moon, J.Y.; Ediriweera, M.K.; Cho, S.K. Phenethyl isothiocyanate suppresses stemness in the chemo- and radio-resistant triple-negative breast cancer cell line MDA-MB-231/IR via downregulation of metadherin. Cancers 2020, 12, 268. [Google Scholar] [CrossRef]
- Psurski, M.; Filip Psurska, B.; Cuprych, M.; Wietrzyk, J.; Oleksyszyn, J. 3,4-dimethoxybenzyl isothiocyanate enhances doxorubicin efficacy in LoVoDX doxorubicin-resistant colon cancer and attenuates its toxicity in vivo. Life Sci. 2019, 231, 116530. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Lin, K.; Hu, Y.; Zhou, Y.; Ding, X.; Wang, Y.; Wu, W. Sulforaphane metabolites inhibit migration and invasion via microtubule-mediated Claudins dysfunction or inhibition of autolysosome formation in human non-small cell lung cancer cells. Cell Death Dis. 2019, 10, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajra, S.; Patra, A.R.; Basu, A.; Saha, P.; Bhattacharya, S. Indole-3-carbinol (I3C) enhances the sensitivity of murine breast adenocarcinoma cells to doxorubicin (DOX) through inhibition of NF-κβ, blocking angiogenesis and regulation of mitochondrial apoptotic pathway. Chem.-Biol. Interact. 2018, 290, 19–36. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.Y.; Lee, S.C.; Liu, H.C.; Peng, S.F.; Chueh, F.S.; Lu, T.J.; Lee, H.T.; Chou, Y.C. Phenethyl isothiocyanate suppresses the proinflammatory cytokines in human glioblastoma cells through the PI3K/AKT/NF-κB signaling pathway in vitro. Oxidative Med. Cell. Longev. 2022, 2022, 2108289. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Salmen, S.H.; Alahmadi, T.A.; Veeraraghavan, V.P.; Surapaneni, K.M.; Natarajan, N.; Subramanian, S. Anticancer effect of selenium/chitosan/polyethylene glycol/allyl isothiocyanate nanocomposites against diethylnitrosamine-induced liver cancer in rats. Saudi J. Biol. Sci. 2022, 29, 3354–3365. [Google Scholar] [CrossRef]
- Lv, X.; Meng, G.; Li, W.; Fan, D.; Wang, X.; Espinoza Pinochet, C.A.; Cespedes Acuña, C.L. Sulforaphane and its antioxidative effects in broccoli seeds and sprouts of different cultivars. Food Chem. 2020, 316, 126216. [Google Scholar] [CrossRef]
- Po, W.W.; Choi, W.S.; Khing, T.M.; Lee, J.Y.; Lee, J.H.; Bang, J.S.; Min, Y.S.; Jeong, J.H.; Sohn, U.D. Benzyl isothiocyanate-induced cytotoxicity via the inhibition of autophagy and lysosomal function in AGS cells. Biomol. Ther. 2022, 30, 348–359. [Google Scholar] [CrossRef]
- Lee, Y.; Chen, M.; Lee, J.D.; Zhang, J.; Lin, S.Y.; Fu, T.M.; Chen, H.; Ishikawa, T.; Chiang, S.Y.; Katon, J.; et al. Reactivation of PTEN tumor suppressor for cancer treatment through inhibition of a MYC-WWP1 inhibitory pathway. Science 2019, 364, eaau0159. [Google Scholar] [CrossRef]
- Russo, M.; Spagnuolo, C.; Russo, G.L.; Skalicka Woźniak, K.; Daglia, M.; Sobarzo Sánchez, E.; Nabavi, S.F.; Nabavi, S.M. Nrf2 targeting by sulforaphane: A potential therapy for cancer treatment. Crit. Rev. Food Sci. Nutr. 2018, 58, 1391–1405. [Google Scholar] [CrossRef]
- Mazumder, A.; Dwivedi, A.; du Plessis, J. Sinigrin and its therapeutic benefits. Molecules 2016, 21, 416. [Google Scholar] [CrossRef]
- Vanduchova, A.; Anzenbacher, P.; Anzenbacherova, E. Isothiocyanate from broccoli, sulforaphane, and its properties. J. Med. Food 2019, 22, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Dinh, T.N.; Parat, M.O.; Ong, Y.S.; Khaw, K.Y. Anticancer activities of dietary benzyl isothiocyanate: A comprehensive review. Pharmacol. Res. 2021, 169, 105666. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, L.; Cao, L.; Zhang, Q.; Song, Q.; Meng, Z.; Wu, X.; Xu, K. Inhibition of autophagy potentiates the anti-metastasis effect of phenethyl isothiocyanate through JAK2/STAT3 pathway in lung cancer cells. Mol. Carcinog. 2018, 57, 522–535. [Google Scholar] [CrossRef] [PubMed]
- Gmelin, R.; Schlüter, M. Isolierung von 4-methylthiobutylglucosinolat (glucoerucin) aus samen von eruca sativa mill. Arch. der Pharm. 1970, 303, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Shertzer, H.G. Indole-3-carbinol protects against covalent binding of benzo[a]pyrene and N-nitrosodimethylamine metabolites to mouse liver macromolecules. Chem. Biol. Interact. 1984, 48, 81–90. [Google Scholar] [CrossRef]
- Fimognari, C.; Turrini, E.; Ferruzzi, L.; Lenzi, M.; Hrelia, P. Natural isothiocyanates: Genotoxic potential versus chemoprevention. Mutat. Res./Rev. Mutat. Res. 2012, 750, 107–131. [Google Scholar] [CrossRef]
- Bianchini, F.; Vainio, H. Isothiocyanates in cancer prevention. Drug Metab. Rev. 2004, 36, 655–667. [Google Scholar] [CrossRef]
- Lahtisham, U.H.; Khan, S.; Awan, K.A.; Iqbal, M.J. Sulforaphane as a potential remedy against cancer: Comprehensive mechanistic review. J. Food Biochem. 2022, 46, e13886. [Google Scholar] [CrossRef]
- Wattenberg, L.W. Inhibition of carcinogenic effects of polycyclic hydrocarbons by benzyl isothiocyanate and related compounds. J. Natl. Cancer Inst. 1977, 58, 395–398. [Google Scholar] [CrossRef]
- Tsai, J.Y.; Tsai, S.H.; Wu, C.C. The chemopreventive isothiocyanate sulforaphane reduces anoikis resistance and anchorage-independent growth in non-small cell human lung cancer cells. Toxicol. Appl. Pharmacol. 2019, 362, 116–124. [Google Scholar] [CrossRef]
- Wang, Z.; Tu, C.; Pratt, R.; Khoury, T.; Qu, J.; Fahey, J.W.; McCann, S.E.; Zhang, Y.; Wu, Y.; Hutson, A.D.; et al. A presurgical-window intervention trial of isothiocyanate-rich broccoli sprout extract in patients with breast cancer. Mol. Nutr. Food Res. 2022, 66, e2101094. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Li, S.; Sargsyan, D.; Yin, R.; Kuo, H.C.; Peter, R.; Wang, L.; Hudlikar, R.; Liu, X.; Kong, A.N. DNA methylome, transcriptome, and prostate cancer prevention by phenethyl isothiocyanate in TRAMP mice. Mol. Carcinog. 2021, 60, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, S.; Helmy, M.W.; Salahuddin, A.; Omran, G. Chemopreventive and antitumor effects of benzyl isothiocynate on HCC models: A possible role of HGF /PAKT/ STAT3 axis and VEGF. Biomed. Pharmacother. 2018, 108, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Gründemann, C.; Huber, R. Chemoprevention with isothiocyanates—From bench to bedside. Cancer Lett. 2018, 414, 26–33. [Google Scholar] [CrossRef]
- Alumkal, J. The Effects of Sulforaphane in Patients with Biochemical Recurrence of Prostate Cancer; U.S. National Library of Medicine: Bethesda, MD, USA, 2017.
- Lee, H. Randomized, Phase II Clinical Trial of Sulforaphane in Bladder Cancer Chemoprevention; U.S. National Library of Medicine: Bethesda, MD, USA, 2020.
- Lin, D. In Vivo Effects of Sulforaphane Supplementation on Normal Human Prostate; U.S. National Library of Medicine: Bethesda, MD, USA, 2016.
- Yuan, J.M. Randomized Clinical Trial of Lung Cancer Chemoprevention with Sulforaphane in Former Smokers; U.S. National Library of Medicine: Bethesda, MD, USA, 2022.
- Shannon, J. Chemoprevention of Prostate Cancer, HDAC Inhibition and DNA Methylation; U.S. National Library of Medicine: Bethesda, MD, USA, 2019.
- Yoxall, V.; Kentish, P.; Coldham, N.; Kuhnert, N.; Sauer, M.J.; Ioannides, C. Modulation of hepatic cytochromes P450 and phase II enzymes by dietary doses of sulforaphane in rats: Implications for its chemopreventive activity. Int. J. Cancer 2005, 117, 356–362. [Google Scholar] [CrossRef]
- Konsue, N.; Ioannides, C. Tissue differences in the modulation of rat cytochromes P450 and phase II conjugation systems by dietary doses of phenethyl isothiocyanate. Food Chem. Toxicol. 2008, 46, 3677–3683. [Google Scholar] [CrossRef]
- Keum, Y.S.; Jeong, W.S.; Tony Kong, A.N. Chemoprevention by isothiocyanates and their underlying molecular signaling mechanisms. Mutat. Res./Fundam. Mol. Mech. Mutagen. 2004, 555, 191–202. [Google Scholar] [CrossRef]
- Cartus, A.T.; Schrenk, D. Metabolism of carcinogenic alpha-asarone by human cytochrome P450 enzymes. Naunyn Schmiedebergs Arch. Pharmacol. 2020, 393, 213–223. [Google Scholar] [CrossRef]
- Guengerich, F.P.; Johnson, W.W.; Shimada, T.; Ueng, Y.F.; Yamazaki, H.; Langouët, S. Activation and detoxication of aflatoxin B1. Mutat. Res. 1998, 402, 121–128. [Google Scholar] [CrossRef]
- Li, Y.; Cui, J.; Jia, J. The activation of procarcinogens by CYP1A1/1B1 and related chemo-preventive agents: A review. Curr. Cancer Drug Targets 2021, 21, 21–54. [Google Scholar] [CrossRef]
- Moorthy, B.; Chu, C.; Carlin, D.J. Polycyclic aromatic hydrocarbons: From metabolism to lung cancer. Toxicol. Sci. 2015, 145, 5–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.S.; Tu, Y.Y.; Koop, D.R.; Coon, M.J. Metabolism of nitrosamines by purified rabbit liver cytochrome P-450 isozymes. Cancer Res. 1985, 45, 1140–1145. [Google Scholar]
- Mitsiogianni, M.; Koutsidis, G.; Mavroudis, N.; Trafalis, D.T.; Botaitis, S.; Franco, R.; Zoumpourlis, V.; Amery, T.; Galanis, A.; Pappa, A.; et al. The role of isothiocyanates as cancer chemo-preventive, chemo-therapeutic and anti-melanoma agents. Antioxidants 2019, 8, E106. [Google Scholar] [CrossRef] [Green Version]
- Abdull Razis, A.F.; Konsue, N.; Ioannides, C. Isothiocyanates and xenobiotic detoxification. Mol. Nutr. Food Res. 2018, 62, e1700916. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, M.; Yoshida, R.; Shimada, N.; Yamazaki, H.; Yokoi, T. Inhibition and inactivation of human cytochrome P450 isoforms by phenethyl isothiocyanate. Drug Metab. Dispos. 2001, 29, 1110–1113. [Google Scholar] [PubMed]
- Skupinska, K.; Misiewicz Krzeminska, I.; Stypulkowski, R.; Lubelska, K.; Kasprzycka Guttman, T. Sulforaphane and its analogues inhibit CYP1A1 and CYP1A2 activity induced by benzo[a]pyrene. J. Biochem. Mol. Toxicol. 2009, 23, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Lněničková, K.; Dymáková, A.; Szotáková, B.; Boušová, I. Sulforaphane alters β-naphthoflavone-induced changes in activity and expression of drug-metabolizing enzymes in rat hepatocytes. Molecules 2017, 22, E1983. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.S.; Chen, X.Y.; Zhu, R.Z.; Choi, B.M.; Kim, B.R. Sulforaphane induces glutathione S-transferase isozymes which detoxify aflatoxin B (1)-8,9-epoxide in AML 12 cells. Biofactors 2010, 36, 289–296. [Google Scholar] [CrossRef]
- La Marca, M.; Beffy, P.; Della Croce, C.; Gervasi, P.G.; Iori, R.; Puccinelli, E.; Longo, V. Structural influence of isothiocyanates on expression of cytochrome P450, phase II enzymes, and activation of Nrf2 in primary rat hepatocytes. Food Chem. Toxicol. 2012, 50, 2822–2830. [Google Scholar] [CrossRef] [Green Version]
- Mallard, A.R.; Spathis, J.G.; Coombes, J.S. Nuclear factor (erythroid-derived 2)-like 2 (Nrf2) and exercise. Free Radic. Biol. Med. 2020, 160, 471–479. [Google Scholar] [CrossRef]
- Michaličková, D.; Hrnčíř, T.; Canová, N.K.; Slanař, O. Targeting Keap1/Nrf2/ARE signaling pathway in multiple sclerosis. Eur. J. Pharmacol. 2020, 873, 172973. [Google Scholar] [CrossRef]
- Tu, W.; Wang, H.; Li, S.; Liu, Q.; Sha, H. The anti-inflammatory and anti-oxidant mechanisms of the Keap1/Nrf2/ARE signaling pathway in chronic diseases. Aging Dis. 2019, 10, 637–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.D.; Lo, S.-C.; Cross, J.V.; Templeton, D.J.; Hannink, M. Keap1 is a redox-regulated substrate adaptor protein for a Cul3-dependent ubiquitin ligase complex. Mol. Cell. Biol. 2004, 24, 10941–10953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Yamamoto, M. Molecular basis of the Keap1-Nrf2 system. Free Radic. Biol. Med. 2015, 88, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.J.; Jeon, J.H. Recent advances in understanding Nrf2 agonism and its potential clinical application to metabolic and inflammatory diseases. Int. J. Mol. Sci. 2022, 23, 2846. [Google Scholar] [CrossRef] [PubMed]
- Papierska, K.; Krajka Kuźniak, V.; Kleszcz, R.; Stefański, T.; Kurczab, R.; Kubicki, M. The synthesis of novel thioderivative chalcones and their influence on NF-κB, STAT3 and Nrf2 signaling pathways in colorectal cancer cells. Sci. Rep. 2022, 12, 14915. [Google Scholar] [CrossRef]
- Song, M.Y.; Lee, D.Y.; Chun, K.S.; Kim, E.H. The role of Nrf2/Keap1 signaling pathway in cancer metabolism. Int. J. Mol. Sci. 2021, 22, 4376. [Google Scholar] [CrossRef]
- Zhou, C.; Huang, Z.; Lin, H.; Ma, Z.; Wang, J.; Wang, Y.; Yu, W. Rhizoma curcumae Longae ameliorates high dietary carbohydrate-induced hepatic oxidative stress, inflammation in golden pompano Trachinotus ovatus. Fish Shellfish Immunol. 2022, 130, 31–42. [Google Scholar] [CrossRef]
- Cykowiak, M.; Krajka Kuźniak, V.; Kleszcz, R.; Kucińska, M.; Szaefer, H.; Piotrowska Kempisty, H.; Plewiński, A.; Murias, M.; Baer Dubowska, W. Comparison of the impact of xanthohumol and phenethyl isothiocyanate and their combination on Nrf2 and NF-κB pathways in HepG2 cells in vitro and tumor burden in vivo. Nutrients 2021, 13, 3000. [Google Scholar] [CrossRef]
- Ernst, I.M.A.; Wagner, A.E.; Schuemann, C.; Storm, N.; Höppner, W.; Döring, F.; Stocker, A.; Rimbach, G. Allyl-, butyl- and phenylethyl-isothiocyanate activate Nrf2 in cultured fibroblasts. Pharmacol. Res. 2011, 63, 233–240. [Google Scholar] [CrossRef]
- Li, J.; Baker, J.; Higham, A.; Shah, R.; Montero Fernandez, A.; Murray, C.; Cooper, N.; Lucas, C.; Fox, C.; Singh, D.; et al. COPD lung studies of Nrf2 expression and the effects of Nrf2 activators. Inflammopharmacology 2022, 30, 1431–1443. [Google Scholar] [CrossRef] [PubMed]
- Dinkova-Kostova, A.T.; Holtzclaw, W.D.; Cole, R.N.; Itoh, K.; Wakabayashi, N.; Katoh, Y.; Yamamoto, M.; Talalay, P. Direct evidence that sulfhydryl groups of Keap1 are the sensors regulating induction of phase 2 enzymes that protect against carcinogens and oxidants. Proc. Natl. Acad. Sci. USA 2002, 99, 11908–11913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yates, M.S.; Kensler, T.W. Keap1 eye on the target: Chemoprevention of liver cancer. Acta Pharmacol. Sin. 2007, 28, 1331–1342. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.S.; Keum, Y.S.; Chen, C.; Jain, M.R.; Shen, G.; Kim, J.H.; Li, W.; Kong, A.N.T. Differential expression and stability of endogenous nuclear factor E2-related factor 2 (Nrf2) by natural chemopreventive compounds in HepG2 human hepatoma cells. J. Biochem. Mol. Biol. 2005, 38, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Saw, C.L.L.; Cintrón, M.; Wu, T.Y.; Guo, Y.; Huang, Y.; Jeong, W.S.; Kong, A.N.T. Pharmacodynamics of dietary phytochemical indoles I3C and DIM: Induction of Nrf2-mediated phase II drug metabolizing and antioxidant genes and synergism with isothiocyanates: Induction Nrf2-mediated genes by indoles and isothiocyanates. Biopharm. Drug Dispos. 2011, 32, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Wu, M.; Lai, Q.; Zhou, W.; Wen, X.; Yin, X. Epigenetic regulation of synaptic disorder in Alzheimer’s disease. Front. Neurosci. 2022, 16, 888014. [Google Scholar] [CrossRef]
- Sundaram, M.K.; R, P.; Haque, S.; Akhter, N.; Khan, S.; Ahmad, S.; Hussain, A. Dietary isothiocyanates inhibit cancer progression by modulation of epigenome. Semin. Cancer Biol. 2022, 83, 353–376. [Google Scholar] [CrossRef]
- Yu, H.; Wang, Y.; Wang, D.; Yi, Y.; Liu, Z.; Wu, M.; Wu, Y.; Zhang, Q. Landscape of the epigenetic regulation in wound healing. Front. Physiol. 2022, 13, 949498. [Google Scholar] [CrossRef]
- Mitsiogianni, M.; Amery, T.; Franco, R.; Zoumpourlis, V.; Pappa, V.; Panayiotidis, M. From chemo-prevention to epigenetic regulation: The role of isothiocyanates in skin cancer prevention. Pharmacol. Ther. 2018, 190, 187–201. [Google Scholar] [CrossRef]
- Chi, P.; Allis, C.D.; Wang, G.G. Covalent histone modifications--miswritten, misinterpreted and mis-erased in human cancers. Nat. Rev. Cancer 2010, 10, 457–469. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.A.; Reddy, D.; Gupta, S. Global histone post-translational modifications and cancer: Biomarkers for diagnosis, prognosis and treatment? World J. Biol. Chem. 2015, 6, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Mannello, F.; Gazzanelli, G. Tissue inhibitors of metalloproteinases and programmed cell death: Conundrums, controversies and potential implications. Apoptosis 2001, 6, 479–482. [Google Scholar] [CrossRef] [PubMed]
- Tischoff, I.; Tannapfel, A. DNA methylation in hepatocellular carcinoma. World J. Gastroenterol. 2008, 14, 1741–1748. [Google Scholar] [CrossRef] [Green Version]
- Umer, M.; Qureshi, S.A.; Hashmi, Z.Y.; Raza, A.; Ahmad, J.; Rahman, M.; Iqbal, M. Promoter hypermethylation of wnt pathway inhibitors in hepatitis C virus-induced multistep hepatocarcinogenesis. Virol. J. 2014, 11, 117. [Google Scholar] [CrossRef] [Green Version]
- Agnieszka, K.S.; Grzegorz, M.; Katarzyna, L.P.; Krystyna, F.M. The role of sulforaphane in epigenetic mechanisms, including interdependence between histone modification and DNA methylation. Int. J. Mol. Sci. 2015, 16, 29732–29743. [Google Scholar] [CrossRef] [Green Version]
- dos Santos, P.W.d.S.; Machado, A.R.T.; De Grandis, R.A.; Ribeiro, D.L.; Tuttis, K.; Morselli, M.; Aissa, A.F.; Pellegrini, M.; Antunes, L.M.G. Transcriptome and DNA methylation changes modulated by sulforaphane induce cell cycle arrest, apoptosis, DNA damage, and suppression of proliferation in human liver cancer cells. Food Chem. Toxicol. 2020, 136, 111047. [Google Scholar] [CrossRef]
- Lu, T.X.; Rothenberg, M.E. MicroRNA. J. Allergy Clin. Immunol. 2018, 141, 1202–1207. [Google Scholar] [CrossRef] [Green Version]
- Basu, A.; Alder, H.; Khiyami, A.; Leahy, P.; Croce, C.M.; Haldar, S. MicroRNA-375 and microRNA-221: Potential noncoding RNAs associated with antiproliferative activity of benzyl isothiocyanate in pancreatic cancer. Genes Cancer 2011, 2, 108–119. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Shu, L.; Kim, H.; Khor, T.O.; Wu, R.; Li, W.; Kong, A.N.T. Phenethyl isothiocyanate (PEITC) suppresses prostate cancer cell invasion epigenetically through regulating microRNA-194. Mol. Nutr. Food Res. 2016, 60, 1427–1436. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, S.; Chen, Y.; Li, X.; Jiang, Y.; Yang, X.; Li, Y.; Wang, Y.; Meng, Y.; Zhu, M.; et al. MiR-19 targeting of GSK3β mediates sulforaphane suppression of lung cancer stem cells. J. Nutr. Biochem. 2017, 44, 80–91. [Google Scholar] [CrossRef]
- Anwar, S.L.; Lehmann, U. DNA methylation, microRNAs, and their crosstalk as potential biomarkers in hepatocellular carcinoma. World J. Gastroenterol. 2014, 20, 7894–7913. [Google Scholar] [CrossRef] [PubMed]
- Ladeiro, Y.; Couchy, G.; Balabaud, C.; Bioulac Sage, P.; Pelletier, L.; Rebouissou, S.; Zucman Rossi, J. MicroRNA profiling in hepatocellular tumors is associated with clinical features and oncogene/tumor suppressor gene mutations. Hepatology 2008, 47, 1955–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, F.; Henson, R.; Wehbe Janek, H.; Ghoshal, K.; Jacob, S.T.; Patel, T. MicroRNA-21 regulates expression of the PTEN tumor suppressor gene in human hepatocellular cancer. Gastroenterology 2007, 133, 647–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; He, H.; Lu, Y.; Ren, W.; Teng, K.; Chiang, C.; Yang, Z.; Yu, B.; Hsu, S.; Jacob, S.T.; et al. Indole-3-carbinol inhibits tumorigenicity of hepatocellular carcinoma cells via suppression of microRNA-21 and upregulation of phosphatase and tensin homolog. Biochim. Biophys. Acta (BBA)—Mol. Cell Res. 2015, 1853, 244–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, M.; Li, W.; Guo, J.; Lu, Y.; Dong, X.; Lin, B.; Chen, Y.; Zhang, X.; Li, M. Alpha fetoprotein antagonises benzyl isothiocyanate inhibition of the malignant behaviors of hepatocellular carcinoma cells. Oncotarget 2016, 7, 75749–75762. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, W.W.; Hou, S.K. The role and mechanism of allyl isothiocyanate on suppressing tumor cells proliferation and invasion of human hepatocellular cancer HepG2 cell line. Int. J. Clin. Exp. Pathol. 2016, 9, 3389–3395. [Google Scholar]
- Lamy, E.; Hertrampf, A.; Herz, C.; Schüler, J.; Erlacher, M.; Bertele, D.; Bakare, A.; Wagner, M.; Weiland, T.; Lauer, U.; et al. Preclinical evaluation of 4-methylthiobutyl isothiocyanate on liver cancer and cancer stem cells with different p53 status. PLoS ONE 2013, 8, e70846. [Google Scholar] [CrossRef]
- Kntayya, S.; Ibrahim, M.; Mohd Ain, N.; Iori, R.; Ioannides, C.; Abdull Razis, A. Induction of apoptosis and cytotoxicity by isothiocyanate sulforaphene in human hepatocarcinoma HepG2 cells. Nutrients 2018, 10, 718. [Google Scholar] [CrossRef] [Green Version]
- Pocasap, P.; Weerapreeyakul, N.; Thumanu, K. Alyssin and iberin in cruciferous vegetables exert anticancer activity in HepG2 by increasing intracellular reactive oxygen species and tubulin depolymerization. Biomol. Ther. 2019, 27, 540–552. [Google Scholar] [CrossRef]
- Wu, S.J.; Ng, L.T.; Lin, C.C. Effects of antioxidants and caspase-3 inhibitor on the phenylethyl isothiocyanate-induced apoptotic signaling pathways in human PLC/PRF/5 cells. Eur. J. Pharmacol. 2005, 518, 96–106. [Google Scholar] [CrossRef]
- Rose, P.; Whiteman, M.; Huang, S.H.; Halliwell, B.; Ong, C.N. Beta-Phenylethyl isothiocyanate-mediated apoptosis in hepatoma HepG2 cells. Cell Mol. Life Sci. 2003, 60, 1489–1503. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Kim, G.Y.; Bae, S.J.; Yoo, Y.H.; Choi, Y.H. Induction of apoptosis by isothiocyanate sulforaphane in human cervical carcinoma HeLa and hepatocarcinoma HepG2 cells through activation of caspase-3. Oncol. Rep. 2007, 18, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.H.; Chyau, C.C.; Chen, J.H.; Tu, S.F.; Lin, H.H.; Chou, F.P. Anti-cancerous effects of Wasabia japonica extract in Hep3B liver cancer cells via ROS accumulation, DNA damage and p73-mediated apoptosis. J. Funct. Foods 2015, 14, 445–455. [Google Scholar] [CrossRef]
- Zou, X.; Qu, Z.; Fang, Y.; Shi, X.; Ji, Y. Endoplasmic reticulum stress mediates sulforaphane-induced apoptosis of HepG2 human hepatocellular carcinoma cells. Mol. Med. Rep. 2017, 15, 331–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pocasap, P.; Weerapreeyakul, N.; Thumanu, K. Structures of isothiocyanates attributed to reactive oxygen species generation and microtubule depolymerization in HepG2 cells. Biomed. Pharmacother. 2018, 101, 698–709. [Google Scholar] [CrossRef]
- Lee, C.; Park, S.H.; Nam, M. Anticarcinogenic effect of indole-3-carbinol (I3C) on human hepatocellular carcinoma SNU449 cells. Hum. Exp. Toxicol. 2019, 38, 136–147. [Google Scholar] [CrossRef]
- Hwang, E.S.; Lee, H.J. Allyl isothiocyanate and its N-acetylcysteine conjugate suppress metastasis via inhibition of invasion, migration, and matrix metalloproteinase-2/-9 activities in SK-Hep1 human hepatoma cells. Exp. Biol. Med. 2006, 231, 421–430. [Google Scholar] [CrossRef]
- Hwang, E.S.; Lee, H.J. Phenylethyl isothiocyanate and its N-acetylcysteine conjugate suppress the metastasis of SK-Hep1 human hepatoma cells. Nutr. Rep. Int. 2006, 17, 837–846. [Google Scholar] [CrossRef]
- Hwang, E.S.; Kim, G.H. Allyl isothiocyanate influences cell adhesion, migration and metalloproteinase gene expression in SK-Hep1 cells. Exp. Biol. Med. 2009, 234, 105–111. [Google Scholar] [CrossRef]
- Hwang, E.S.; Lee, H.J. Benzyl isothiocyanate inhibits metalloproteinase-2/-9 expression by suppressing the mitogen-activated protein kinase in SK-Hep1 human hepatoma cells. Food Chem. Toxicol. 2008, 46, 2358–2364. [Google Scholar] [CrossRef]
- Gupta, B.; Chiang, L.; Chae, K.M.; Lee, D.H. Phenethyl isothiocyanate inhibits hypoxia-induced accumulation of HIF-1α and VEGF expression in human glioma cells. Food Chem. 2013, 141, 1841–1846. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Atkinson, S.J.; Akbareian, S.E.; Zhou, Z.; Munsterberg, A.; Robinson, S.D.; Bao, Y. Sulforaphane exerts anti-angiogenesis effects against hepatocellular carcinoma through inhibition of STAT3/HIF-1α/VEGF signalling. Sci. Rep. 2017, 7, 12651. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.O.; Kang, S.H.; Kim, K.C.; Kim, M.O.; Choi, Y.H.; Kim, G.Y. Sulforaphane decreases viability and telomerase activity in hepatocellular carcinoma Hep3B cells through the reactive oxygen species-dependent pathway. Cancer Lett. 2010, 295, 260–266. [Google Scholar] [CrossRef]
- Lamy, E.; Herz, C.; Lutz Bonengel, S.; Hertrampf, A.; Márton, M.-R.; Mersch Sundermann, V. The MAPK pathway signals telomerase modulation in response to isothiocyanate-induced DNA damage of human liver cancer cells. PLoS ONE 2013, 8, e53240. [Google Scholar] [CrossRef] [Green Version]
- Herz, C.; Hertrampf, A.; Zimmermann, S.; Stetter, N.; Wagner, M.; Kleinhans, C.; Erlacher, M.; Schüler, J.; Platz, S.; Rohn, S.; et al. The isothiocyanate erucin abrogates telomerase in hepatocellular carcinoma cells in vitro and in an orthotopic xenograft tumour model of HCC. J. Cell Mol. Med. 2014, 18, 2393–2403. [Google Scholar] [CrossRef] [PubMed]
- Jie, M.; Cheung, W.M.; Yu, V.; Zhou, Y.; Tong, P.H.; Ho, J.W.S. Anti-proliferative activities of sinigrin on carcinogen-induced hepatotoxicity in rats. PLoS ONE 2014, 9, e110145. [Google Scholar] [CrossRef]
- Wang, S.; Zhou, D.; Xu, Z.; Song, J.; Qian, X.; Lv, X.; Luan, J. Anti-tumor drug targets analysis: Current insight and future prospect. Curr. Drug Targets 2019, 20, 1180–1202. [Google Scholar] [CrossRef]
- Han, K.W.W.; Po, W.W.; Sohn, U.D.; Kim, H.J. Benzyl isothiocyanate induces apoptosis via reactive oxygen species-initiated mitochondrial dysfunction and DR4 and DR5 death receptor activation in gastric adenocarcinoma cells. Biomolecules 2019, 9, 839. [Google Scholar] [CrossRef] [Green Version]
- Núñez Iglesias, M.J.; Novio, S.; García Santiago, C.; Cartea, M.E.; Soengas, P.; Velasco, P.; Freire Garabal, M. Effects of 3-butenyl isothiocyanate on phenotypically different prostate cancer cells. Int. J. Oncol. 2018, 53, 2213–2223. [Google Scholar] [CrossRef] [Green Version]
- Ye, Z.; Zheng, M.; Zeng, Y.; Wei, S.; Wang, Y.; Lin, Z.; Shu, C.; Xie, Y.; Zheng, Q.; Chen, L. Bioinformatics analysis reveals an association between cancer cell stemness, gene mutations, and the immune microenvironment in stomach adenocarcinoma. Front. Genet. 2020, 11, 595477. [Google Scholar] [CrossRef]
- Besson, A.; Dowdy, S.F.; Roberts, J.M. CDK inhibitors: Cell cycle regulators and beyond. Dev. Cell 2008, 14, 159–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, T.; Sicinski, P. Cell cycle proteins as promising targets in cancer therapy. Nat. Rev. Cancer 2017, 17, 93–115. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.C.; Huang, W.W.; Huang, W.C.; Lu, C.C.; Chiang, J.H.; Peng, S.F.; Chung, J.G.; Lin, Y.H.; Hsu, Y.M.; Amagaya, S.; et al. ERK-modulated intrinsic signaling and G2/M phase arrest contribute to the induction of apoptotic death by allyl isothiocyanate in MDA-MB-468 human breast adenocarcinoma cells. Int. J. Oncol. 2012, 41, 2065–2072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldar, S.; Khaniani, M.S.; Derakhshan, S.M.; Baradaran, B. Molecular mechanisms of apoptosis and roles in cancer development and treatment. Asian Pac. J. Cancer Prev. 2015, 16, 2129–2144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, H.; Tian, M.; Ding, C.; Yu, S. The C/EBP homologous protein (CHOP) transcription factor functions in endoplasmic reticulum stress-induced apoptosis and mcrobial infection. Front. Immunol. 2018, 9, 3083. [Google Scholar] [CrossRef] [Green Version]
- Rousalova, I.; Krepela, E. Granzyme B-induced apoptosis in cancer cells and its regulation (review). Int. J. Oncol. 2010, 37, 1361–1378. [Google Scholar] [CrossRef] [Green Version]
- Chiang, J.H.; Tsai, F.J.; Hsu, Y.M.; Yin, M.C.; Chiu, H.Y.; Yang, J.S. Sensitivity of allyl isothiocyanate to induce apoptosis via ER stress and the mitochondrial pathway upon ROS production in colorectal adenocarcinoma cells. Oncol. Rep. 2020, 44, 1415–1424. [Google Scholar] [CrossRef]
- Henklewska, M.; Pawlak, A.; Li, R.F.; Yi, J.; Zbyryt, I.; Obminska Mrukowicz, B. Benzyl isothiocyanate, a vegetable-derived compound, induces apoptosis via ROS accumulation and DNA damage in canine lymphoma and leukemia cells. Int. J. Mol. Sci. 2021, 22, 11772. [Google Scholar] [CrossRef]
- Sita, G.; Graziosi, A.; Hrelia, P.; Morroni, F. Sulforaphane causes cell cycle arrest and apoptosis in human glioblastoma U87MG and U373MG cell lines under hypoxic conditions. Int. J. Mol. Sci. 2021, 22, 11201. [Google Scholar] [CrossRef]
- Estaquier, J.; Vallette, F.; Vayssiere, J.L.; Mignotte, B. The mitochondrial pathways of apoptosis. Adv. Exp. Med. Biol. 2012, 942, 157–183. [Google Scholar] [CrossRef]
- Noutsopoulos, D.; Markopoulos, G.; Vartholomatos, G.; Kolettas, E.; Kolaitis, N.; Tzavaras, T. VL30 retrotransposition signals activation of a caspase-independent and p53-dependent death pathway associated with mitochondrial and lysosomal damage. Cell Res. 2010, 20, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Marciniak, S.J.; Chambers, J.E.; Ron, D. Pharmacological targeting of endoplasmic reticulum stress in disease. Nat. Rev. Drug Discov. 2022, 21, 115–140. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.W.; Wang, Z.M.; Sun, S.M.; Su, Y.; Li, Z.H.; Shao, J.J.; Tan, S.Z.; Chen, A.P.; Wang, S.J.; Zhang, Z.L.; et al. Endoplasmic reticulum stress and protein degradation in chronic liver disease. Pharmacol. Res. 2020, 161, 105218. [Google Scholar] [CrossRef]
- Evyapan, G.; Ay, G.; Comertpay, G.; Luleyap, H.U. Role of endoplasmic reticulum stress response in tumorogenesis. Cukurova Med. J. 2019, 44, 241–248. [Google Scholar] [CrossRef] [Green Version]
- He, Q.; Wang, T.; Ni, H.; Liu, Q.; An, K.; Tao, J.; Chen, Y.; Xu, L.; Zhu, C.; Yao, M. Endoplasmic reticulum stress promoting caspase signaling pathway-dependent apoptosis contributes to bone cancer pain in the spinal dorsal horn. Mol. Pain 2019, 15, 1744806919876150. [Google Scholar] [CrossRef] [PubMed]
- Long, D.; Chen, K.; Yang, Y.; Tian, X. Unfolded protein response activated by endoplasmic reticulum stress in pancreatic cancer: Potential therapeutical target. Front. Biosci. 2021, 26, 1689–1696. [Google Scholar] [CrossRef]
- Tai, H.C.; Chung, S.D.; Chien, C.T.; Yu, H.J. Sulforaphane improves ischemia-induced detrusor overactivity by downregulating the enhancement of associated endoplasmic reticulum stress, autophagy, and apoptosis in rat bladder. Sci. Rep. 2016, 6, 36110. [Google Scholar] [CrossRef]
- Zhang, Q.; Pan, Z.; Liu, B.; Meng, Z.; Wu, X.; Zhou, Q.; Xu, K. Benzyl isothiocyanate induces protective autophagy in human lung cancer cells through an endoplasmic reticulum stress-mediated mechanism. Acta Pharmacol. Sin. 2017, 38, 539–550. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, M.; Cao, L.; Ren, Y.; Guo, X.; Wu, X.; Xu, K. Phenethyl isothiocyanate synergistically induces apoptosis with Gefitinib in non-small cell lung cancer cells via endoplasmic reticulum stress-mediated degradation of Mcl-1. Mol. Carcinog. 2020, 59, 590–603. [Google Scholar] [CrossRef]
- Bertolotti, A.; Zhang, Y.; Hendershot, L.M.; Harding, H.P.; Ron, D. Dynamic interaction of BiP and ER stress transducers in the unfolded-protein response. Nat. Cell Biol. 2000, 2, 326–332. [Google Scholar] [CrossRef]
- Zhou, Y.; Lee, J.; Reno, C.M.; Sun, C.; Park, S.W.; Chung, J.; Lee, J.; Fisher, S.J.; White, M.F.; Biddinger, S.B.; et al. Regulation of glucose homeostasis through a XBP-1-FoxO1 interaction. Nat. Med. 2011, 17, 356–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wulf, S.B.; Ulrike, H.; Martin, E. Membrane trafficking of death receptors: Implications on signalling. Int. J. Mol. Sci. 2013, 14, 14475–14503. [Google Scholar] [CrossRef]
- Yoon, J.H.; Gores, G.J. Death receptor-mediated ap.poptosis and the liver. J. Hepatol. 2002, 37, 400–410. [Google Scholar] [CrossRef]
- Cha, Z.; Cheng, J.; Xiang, H.; Qin, J.; He, Y.; Peng, Z.; Jia, J.; Yu, H. Celastrol enhances TRAIL-induced apoptosis in human glioblastoma via the death receptor pathway. Cancer Chemother. Pharm. 2019, 84, 719–728. [Google Scholar] [CrossRef]
- Huang, C.C.; Cheng, Y.C.; Lin, Y.C.; Chou, C.H.; Ho, C.T.; Wang, H.K.; Way, T.D. CSC-3436 sensitizes triple negative breast cancer cells to TRAIL-induced apoptosis through ROS-mediated p38/CHOP/death receptor 5 signaling pathways. Environ. Toxicol. 2021, 36, 2578–2588. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Ji, H.; Dong, X.; Liu, A.; Yu, J. FAS/FAS-L-mediated apoptosis and autophagy of SPC-A-1 cells induced by water-soluble polysaccharide from Polygala tenuifolia. Int. J. Biol. Macromol. 2020, 150, 449–458. [Google Scholar] [CrossRef]
- Yang, M.; Ren, M.; Qu, Y. Sulforaphene inhibits hepatocellular carcinoma through repressing keratin 8 and activating anoikis. RSC Adv. 2016, 6, 70326–70334. [Google Scholar] [CrossRef]
- Abou El Naga, R.N.; Azab, S.S.; El Demerdash, E.; Shaarawy, S.; El Merzabani, M.; Ammar, E.S.M. Sensitization of TRAIL-induced apoptosis in human hepatocellular carcinoma HepG2 cells by phytochemicals. Life Sci. 2013, 92, 555–561. [Google Scholar] [CrossRef]
- Scheau, C.; Badarau, I.A.; Costache, R.; Caruntu, C.; Mihai, G.L.; Didilescu, A.C.; Constantin, C.; Neagu, M. The role of matrix metalloproteinases in the epithelial-mesenchymal transition of hepatocellular carcinoma. Anal. Cell Pathol. 2019, 2019, 9423907. [Google Scholar] [CrossRef] [Green Version]
- Minard, M.E.; Ellis, L.M.; Gallick, G.E. Tiam1 regulates cell adhesion, migration and apoptosis in colon tumor cells. Clin. Exp. Metastasis 2006, 23, 301. [Google Scholar] [CrossRef]
- Zhu, M.; Li, W.; Dong, X.; Chen, Y.; Lu, Y.; Lin, B.; Guo, J.; Li, M. Benzyl-isothiocyanate induces apoptosis and inhibits migration and invasion of hepatocellular carcinoma cells in vitro. J. Cancer 2017, 8, 240–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morse, M.A.; Sun, W.; Kim, R.; He, A.R.; Abada, P.B.; Mynderse, M.; Finn, R.S. The role of angiogenesis in hepatocellular carcinoma. Clin. Cancer Res. 2019, 25, 912–920. [Google Scholar] [CrossRef] [PubMed]
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in signaling and disease: Beyond discovery and development. Cell 2019, 176, 1248–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vempati, P.; Popel, A.S.; Gabhann, F.M. Extracellular regulation of VEGF: Isoforms, proteolysis, and vascular patterning. Cytokine Growth Factor Rev. 2014, 25, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.; Zhang, Y.; Liu, D.; Songyang, Z.; Wan, M. Telomeres—Structure, function, and regulation. Exp. Cell Res. 2013, 319, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Pickett, H.A. Targeting telomeres: Advances in telomere maintenance mechanism-specific cancer therapies. Nat. Rev. Cancer 2022, 22, 515–532. [Google Scholar] [CrossRef]
- Zhou, J.; Sun, H.; Wang, Z.; Cong, W.; Wang, J.; Zeng, M.; Zhou, W.; Bie, P.; Liu, L.; Wen, T.; et al. Guidelines for the diagnosis and treatment of hepatocellular carcinoma (2019 Edition). Liver Cancer 2020, 9, 682–720. [Google Scholar] [CrossRef]
- Antonini, E.; Lori, R.; Ninfali, P.; Scarpa, E.S. A combination of moringin and avenanthramide 2f inhibits the proliferation of Hep3B liver cancer cells inducing intrinsic and extrinsic apoptosis. Nutr. Cancer 2018, 70, 1159–1165. [Google Scholar] [CrossRef]
- Violetta, K.K. Combination of xanthohumol and phenethyl isothiocyanate inhibits NF-κB and activates Nrf2 in pancreatic cancer cells. Toxicol. Vitr. 2020, 65, 104799. [Google Scholar]
- Xu, Y.; Han, X.; Li, Y.; Min, H.; Zhao, X.; Zhang, Y.; Qi, Y.; Shi, J.; Qi, S.; Bao, Y.; et al. Sulforaphane mediates glutathione depletion via polymeric nanoparticles to restore cisplatin chemosensitivity. ACS Nano 2019, 13, 13445–13455. [Google Scholar] [CrossRef]
- Ren, K.; Li, Z.; Li, Y.; Zhang, W.; Han, X. Sulforaphene enhances radiosensitivity of hepatocellular carcinoma through suppression of the NF-κB pathway. J. Biochem. Mol. Toxicol. 2017, 31, e21917. [Google Scholar] [CrossRef]
- Yang, Y.T.; Shi, Y.; Jay, M.; Di Pasqua, A.J. Enhanced toxicity of cisplatin with chemosensitizer phenethyl isothiocyanate toward non-small cell lung cancer cells when delivered in liposomal nanoparticles. Chem. Res. Toxicol. 2014, 27, 946–948. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Isothiocyanates | Structures | Glucosinolate Precursors | Dietary Sources | References |
|---|---|---|---|---|
| Allyl isothiocyanate (AITC) |  | Sinigrin | Broccoli, brussels sprouts, and brassicanigra | [70] |
| Sulforaphane (SFN) |  | Glucoraphanin | Broccoli | [71] |
| Benzyl isothiocyanate (BITC) |  | Glucotropaeolin | Watercress, cabbage, and broccoli | [72] |
| Phenethyl isothiocyanate (PEITC) |  | Gluconasturtiin | Broccoli, brussels sprouts, and watercress | [73] |
| 4-(methylthio) butyl isothiocyanate (4-MTB-ITC) |  | Glucoerrucin | Rocket salad | [74] |
| Indole-3-carbinol (I3C, a derivative of indolic ITCs) |  | Glucobrassicin | Cabbage, broccoli, brussels sprouts, and cauliflower | [75] |
| Isothiocyanates | Types of Cancers | ClinicalTrial.gov Identifiers a | Doses | References |
|---|---|---|---|---|
| SFN | Lung cancer | NCT03232138 | Oral-120 μM/day | [88] |
| SFN | Bladder cancer | NCT03517995 | Oral-200 μM/day | [86] |
| SFN | Prostate cancer | NCT01265953 | Oral-200 μM/day | [89] |
| SFN | Prostate cancer | NCT01228084 | Oral-200 μM/day | [85] |
| SFN | Prostate cancer | NCT00946309 | Oral-100 μM/day | [87] |
| SFN | Liver cancer | / | 12 mg/kg | [90] |
| PEITC | Live cancer | / | 0.6–6.0 μM/g | [91] |
| Isothiocyanates | Inhibitory Approaches | Molecular Targets a | Experimental Models | Concentrations | References |
|---|---|---|---|---|---|
| BITC | Inhibit cell proliferation | Survivin↓ | Bel7402 and HLE | 20 μM | [140] |
| AITC | Inhibit cell proliferation | Survivin↓ | HepG2 | 2 and 5 μM | [141] |
| MTBITC | Arrest cell cycle | G2/M phase arrest | HepG2 | 25 μM | [142] |
| AITC | Arrest cell cycle | G2/M phase arrest Cyclin B1↓, p53↑, and p21↑ | HepG2 | 2 and 5 μM | [141] |
| SFN | Arrest cell cycle | Sub G0/G1 phase arrest | HepG2 | 33.8 μM | [143] |
| SFN | Arrest cell cycle | G2/M phase arrest | HepG2 | 8 μM | [131] |
| SFN | Arrest cell cycle | S; G2/M phase arrest | HepG2 | 40 μM | [144] |
| Iberin | Arrest cell cycle | S; G2/M phase arrest | HepG2 | 40 μM | [144] |
| Alyssin | Arrest cell cycle | S; G2/M phase arrest | HepG2 | 40 μM | [144] |
| PEITC | Induce cell apoptosis | caspase-9/-3/-8↑, Bax↑, p53↑, Bcl-2↓, BclXL↓, and cytochrome C↓ | PLC/PRF/5 cells | 5 μM | [145] |
| β-PEITC | Induce cell apoptosis | caspase-9/-3↑, Bax↑, mitochondrial membrane potential↓, cytochrome C↓ | HepG2 | 20μM | [146] |
| SFN | Induce cell apoptosis | caspase-3↑, Bcl-2, BclXL↓, and Bax↑ | HepG2 | 20 μM | [147] |
| MTBITC | Induce cell apoptosis | Caspase-3/-7↑ | HepG2 | 25 μM | [142] |
| Wasabia japonica extract contained 5-(methylsulfinyl) pentyl ITC, 6-(methylsulfinyl) hexyl ITC, and 7-(methylsulfinyl) heptyl ITC) | Induce cell apoptosis | ROS↑ and p73↑ | Hep3B | 0.25 to 1 mg/mL | [148] |
| Wasabia japonica extract contained 5-(methylsulfinyl) pentyl ITC, 6-(methylsulfinyl) hexyl ITC, and 7-(methylsulfinyl) heptyl ITC) | Induce cell apoptosis | ROS↑ and p73↑ | Xenograft tumors | 5 mg/kg | [148] |
| BITC | Induce cell apoptosis | caspase-3↑ and PARP-1↑ | Bel7402 | 20 μM | [140] |
| AITC | Induce cell apoptosis | caspase-3/-8↑ and Bcl-2↓ | HepG2 | 2 and 5 μM | [141] |
| SFN | Induce cell apoptosis | Bip/RP78↓, XBP-1↓, caspase-12↓, CHOP/GADD153↓, and Bid↓ | HepG2 | 40 μM | [149] |
| SFN | Induce cell apoptosis | caspases-3/7/-9↑ caspases-8↓ | HepG2 | 33.8 μM | [143] |
| MTBITC | Induce cell apoptosis | ROS↑ | HepG2 | 10, 20, and 40 μM | [150] |
| Erysolin | Induce cell apoptosis | ROS↑ | HepG2 | 10, 20, and 40 μM | [150] |
| PEITC | Induce cell apoptosis | ROS↑ | HepG2 | 10, 20, and 40 μM | [150] |
| SFN | Induce cell apoptosis | ROS↑ | HepG2 | 10, 20, and 40 μM | [150] |
| Sulforaphene | Induce cell apoptosis | ROS↑ | HepG2 | 10, 20, and 40 μM | [150] |
| I3C | Induce cell apoptosis | p53↑, PARP↑, and caspase-3/-7↑ | SNU449 | 300 μM | [151] |
| AITC | Inhibit cell migration | MMP-2/-9↓ | SK-Hep-1 | 5 μM | [152] |
| AITC | Inhibit cell migration | MMP-2/-9↓, integrinα5β1↓ | HepG2 | 2, and 5 μM | [141] |
| PEITC | Inhibit cell migration | MMP-2/-9↓ and TIMP1/2↑ | SK-Hep-1 | 5 μM | [153] |
| AITC | Inhibit cell migration | COL8A1↓, COL4A3↓, and MMP-2/-9↓ | SK-Hep-1 | 10 μM | [154] |
| I3C | Inhibit cell migration | miR-21↓ and PTEN↑ | SK-Hep-1 and SUN449 | 200 μM | [139] |
| BITC | Inhibit cell migration | MMP-2/-9↓ and CXCR4↓ | Bel7402 | 20 μM | [140] |
| BITC | Inhibit cell migration | MMP-2/-9↓ | SK-Hep-1 | 0.1, 1, and 5μM | [155] |
| AITC | Inhibit cell migration | MMP-2/-9↓ and AKT/NF-κB pathway | HepG2 | 2, and 5 μM | [141] |
| PEITC | Inhibit cell angiogenesis | HIF-1α↓ and VEGF↓ | HepG2 | 10 μM | [156] |
| MTBITC | Inhibit cell angiogenesis | microtubule depolymerization | HepG2 | 10, 20, and 40 μM | [150] |
| Erysolin | Inhibit cell angiogenesis | microtubule depolymerization | HepG2 | 10, 20, and 40 μM | [150] |
| PEITC | Inhibit cell angiogenesis | microtubule depolymerization | HepG2 | 10, 20, and 40 μM | [150] |
| SFN | Inhibit cell angiogenesis | microtubule depolymerization | HepG2 | 10, 20, and 40 μM | [150] |
| Sulforaphene | Inhibit cell angiogenesis | microtubule depolymerization | HepG2 | 10, 20, and 40 μM | [150] |
| I3C | Inhibit cell angiogenesis | p53↑, PARP↑, and caspase-3/-7↑ | SNU449 | 300 μM | [151] |
| SFN | Inhibit cell angiogenesis | STAT3↓, HIF-1α↓, and VEGF↓ | HepG2 | 20 μM | [157] |
| SFN | Decrease telomerase activity | hTERT↑ and ROS↑ | Hep3B | 20 μM | [158] |
| MTBITC | Decrease telomerase activity | MAPK and hTERT↑ | HepG2, Hep3B, and Huh7 | 25 μM | [159] |
| MTBITC | Decrease telomerase activity | telomerase activity↓ | Xenograft tumors | 50 mg/kg | [160] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Huang, H.; Jin, L.; Lin, S. Anticarcinogenic Effects of Isothiocyanates on Hepatocellular Carcinoma. Int. J. Mol. Sci. 2022, 23, 13834. https://doi.org/10.3390/ijms232213834
Zhang Y, Huang H, Jin L, Lin S. Anticarcinogenic Effects of Isothiocyanates on Hepatocellular Carcinoma. International Journal of Molecular Sciences. 2022; 23(22):13834. https://doi.org/10.3390/ijms232213834
Chicago/Turabian StyleZhang, Yuting, Huiting Huang, Libo Jin, and Sue Lin. 2022. "Anticarcinogenic Effects of Isothiocyanates on Hepatocellular Carcinoma" International Journal of Molecular Sciences 23, no. 22: 13834. https://doi.org/10.3390/ijms232213834