
Cell-Specific Transport and Thyroid Hormone Receptor Isoform Selectivity Account for Hepatocyte-Targeted Thyromimetic Action of MGL-3196
 , ,
, ,
Abstract
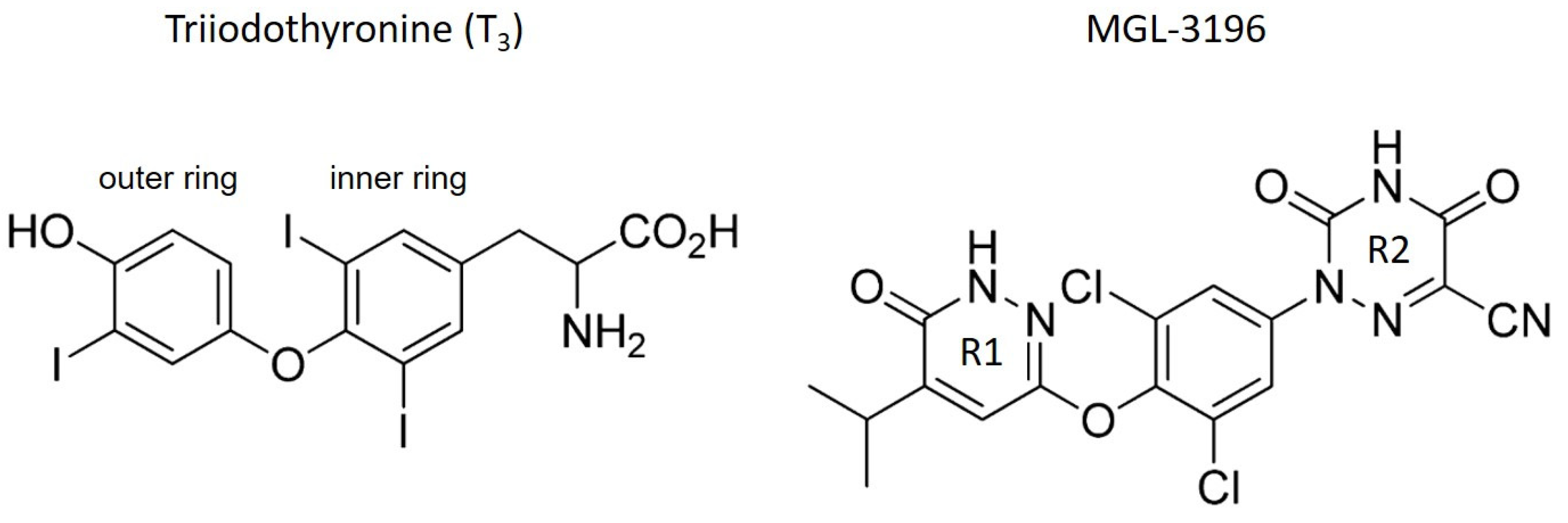
:1. Introduction
2. Results and Discussion
2.1. Expression of TH Transporters and TRs in HEK293 Cells
2.2. MGL-3196 Transport Is Mediated by OATP1B1
2.3. MGL-3196 Predominantly Acts via TRβ
2.4. Equivalent Induction of TH Target Genes by MGL-3196 and T3
2.5. MGL-3196 Improves Mitochondrial Function
3. Conclusions
4. Material and Methods
4.1. Chemicals
4.2. Cell Culture and Transfection
4.3. Luciferase Assay
4.4. Isolation and Cultivation of Primary Murine Hepatocytes
4.5. Gene Expression Analysis
4.6. Seahorse XF-24 Mito Stress Test
4.7. Statistics
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sinha, R.A.; Singh, B.K.; Yen, P.M. Direct effects of thyroid hormones on hepatic lipid metabolism. Nat. Reviews. Endocrinol. 2018, 14, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Araki, O.; Ying, H.; Zhu, X.G.; Willingham, M.C.; Cheng, S.Y. Distinct dysregulation of lipid metabolism by unliganded thyroid hormone receptor isoforms. Mol. Endocrinol. 2009, 23, 308–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groeneweg, S.; van Geest, F.S.; Peeters, R.P.; Heuer, H.; Visser, W.E. Thyroid Hormone Transporters. Endocr. Rev. 2019, 41, 146–201. [Google Scholar] [CrossRef] [PubMed]
- Bookout, A.L.; Jeong, Y.; Downes, M.; Yu, R.T.; Evans, R.M.; Mangelsdorf, D.J. Anatomical profiling of nuclear receptor expression reveals a hierarchical transcriptional network. Cell 2006, 126, 789–799. [Google Scholar] [CrossRef] [Green Version]
- Minakhina, S.; Bansal, S.; Zhang, A.; Brotherton, M.; Janodia, R.; De Oliveira, V.; Tadepalli, S.; Wondisford, F.E. A Direct Comparison of Thyroid Hormone Receptor Protein Levels in Mice Provides Unexpected Insights into Thyroid Hormone Action. Thyroid 2020, 30, 1193–1204. [Google Scholar] [CrossRef]
- Harvey, C.B.; O’Shea, P.J.; Scott, A.J.; Robson, H.; Siebler, T.; Shalet, S.M.; Samarut, J.; Chassande, O.; Williams, G.R. Molecular mechanisms of thyroid hormone effects on bone growth and function. Mol. Genet. Metab. 2002, 75, 17–30. [Google Scholar] [CrossRef]
- Elbers, L.P.; Kastelein, J.J.; Sjouke, B. Thyroid Hormone Mimetics: The Past, Current Status and Future Challenges. Curr. Atheroscler. Rep. 2016, 18, 14. [Google Scholar] [CrossRef] [Green Version]
- Scanlan, T.S. Sobetirome: A case history of bench-to-clinic drug discovery and development. Heart Fail. Rev. 2010, 15, 177–182. [Google Scholar] [CrossRef]
- Vatner, D.F.; Weismann, D.; Beddow, S.A.; Kumashiro, N.; Erion, D.M.; Liao, X.H.; Grover, G.J.; Webb, P.; Phillips, K.J.; Weiss, R.E.; et al. Thyroid hormone receptor-beta agonists prevent hepatic steatosis in fat-fed rats but impair insulin sensitivity via discrete pathways. Am. J. Physiology. Endocrinol. Metab. 2013, 305, E89–E100. [Google Scholar] [CrossRef]
- Kowalik, M.A.; Columbano, A.; Perra, A. Thyroid Hormones, Thyromimetics and Their Metabolites in the Treatment of Liver Disease. Front. Endocrinol. 2018, 9, 382. [Google Scholar] [CrossRef]
- Kelly, M.J.; Pietranico-Cole, S.; Larigan, J.D.; Haynes, N.E.; Reynolds, C.H.; Scott, N.; Vermeulen, J.; Dvorozniak, M.; Conde-Knape, K.; Huang, K.S.; et al. Discovery of 2-[3,5-dichloro-4-(5-isopropyl-6-oxo-1,6-dihydropyridazin-3-yloxy)phenyl]-3,5-dio xo-2,3,4,5-tetrahydro[1,2,4]triazine-6-carbonitrile (MGL-3196), a Highly Selective Thyroid Hormone Receptor beta agonist in clinical trials for the treatment of dyslipidemia. J. Med. Chem. 2014, 57, 3912–3923. [Google Scholar] [CrossRef] [PubMed]
- Harrison, S.A.; Bashir, M.R.; Guy, C.D.; Zhou, R.; Moylan, C.A.; Frias, J.P.; Alkhouri, N.; Bansal, M.B.; Baum, S.; Neuschwander-Tetri, B.A.; et al. Resmetirom (MGL-3196) for the treatment of non-alcoholic steatohepatitis: A multicentre, randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2019, 394, 2012–2024. [Google Scholar] [CrossRef]
- Harrison, S.A.; Bashir, M.; Moussa, S.E.; McCarty, K.; Pablo Frias, J.; Taub, R.; Alkhouri, N. Effects of Resmetirom on Noninvasive Endpoints in a 36-Week Phase 2 Active Treatment Extension Study in Patients With NASH. Hepatol. Commun. 2021, 5, 573–588. [Google Scholar] [CrossRef] [PubMed]
- Luong, X.G.; Stevens, S.K.; Jekle, A.; Lin, T.I.; Gupta, K.; Misner, D.; Chanda, S.; Mukherjee, S.; Williams, C.; Stoycheva, A.; et al. Regulation of gene transcription by thyroid hormone receptor β agonists in clinical development for the treatment of non-alcoholic steatohepatitis (NASH). PLoS ONE 2020, 15, e0240338. [Google Scholar] [CrossRef]
- Obaidat, A.; Roth, M.; Hagenbuch, B. The expression and function of organic anion transporting polypeptides in normal tissues and in cancer. Annu. Rev. Pharmacol. Toxicol. 2012, 52, 135–151. [Google Scholar] [CrossRef] [Green Version]
- Abe, T.; Kakyo, M.; Tokui, T.; Nakagomi, R.; Nishio, T.; Nakai, D.; Nomura, H.; Unno, M.; Suzuki, M.; Naitoh, T.; et al. Identification of a novel gene family encoding human liver-specific organic anion transporter LST-1. J. Biol. Chem. 1999, 274, 17159–17163. [Google Scholar] [CrossRef] [Green Version]
- Kinne, A.; Wittner, M.; Wirth, E.K.; Hinz, K.M.; Schulein, R.; Kohrle, J.; Krause, G. Involvement of the L-Type Amino Acid Transporter Lat2 in the Transport of 3,3’-Diiodothyronine across the Plasma Membrane. Eur. Thyroid J. 2015, 4, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Trajkovic, M.; Visser, T.J.; Mittag, J.; Horn, S.; Lukas, J.; Darras, V.M.; Raivich, G.; Bauer, K.; Heuer, H. Abnormal thyroid hormone metabolism in mice lacking the monocarboxylate transporter 8. J. Clin. Invest. 2007, 117, 627–635. [Google Scholar] [CrossRef] [Green Version]
- Visser, W.E.; Friesema, E.C.; Visser, T.J. Minireview: Thyroid hormone transporters: The knowns and the unknowns. Mol. Endocrinol. 2011, 25, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Hones, G.S.; Rakov, H.; Logan, J.; Liao, X.H.; Werbenko, E.; Pollard, A.S.; Praestholm, S.M.; Siersbaek, M.S.; Rijntjes, E.; Gassen, J.; et al. Noncanonical thyroid hormone signaling mediates cardiometabolic effects in vivo. Proc. Natl. Acad. Sci. USA 2017, 114, E11323–E11332. [Google Scholar] [CrossRef]
- Dong, J.; Olaleye, O.E.; Jiang, R.; Li, J.; Lu, C.; Du, F.; Xu, F.; Yang, J.; Wang, F.; Jia, W.; et al. Glycyrrhizin has a high likelihood to be a victim of drug-drug interactions mediated by hepatic organic anion-transporting polypeptide 1B1/1B3. Br. J. Pharmacol. 2018, 175, 3486–3503. [Google Scholar] [CrossRef] [PubMed]
- Ishida, K.; Ullah, M.; Tóth, B.; Juhasz, V.; Unadkat, J.D. Transport Kinetics, Selective Inhibition, and Successful Prediction of In Vivo Inhibition of Rat Hepatic Organic Anion Transporting Polypeptides. Drug Metab. Dispos. Biol. Fate Chem. 2018, 46, 1251–1258. [Google Scholar] [CrossRef]
- van der Deure, W.M.; Friesema, E.C.; de Jong, F.J.; de Rijke, Y.B.; de Jong, F.H.; Uitterlinden, A.G.; Breteler, M.M.; Peeters, R.P.; Visser, T.J. Organic anion transporter 1B1: An important factor in hepatic thyroid hormone and estrogen transport and metabolism. Endocrinology 2008, 149, 4695–4701. [Google Scholar] [CrossRef] [PubMed]
- Roth, M.; Obaidat, A.; Hagenbuch, B. OATPs, OATs and OCTs: The organic anion and cation transporters of the SLCO and SLC22A gene superfamilies. Br. J. Pharmacol. 2012, 165, 1260–1287. [Google Scholar] [CrossRef] [Green Version]
- Taub, R.; Chiang, E.; Chabot-Blanchet, M.; Kelly, M.J.; Reeves, R.A.; Guertin, M.C.; Tardif, J.C. Lipid lowering in healthy volunteers treated with multiple doses of MGL-3196, a liver-targeted thyroid hormone receptor-beta agonist. Atherosclerosis 2013, 230, 373–380. [Google Scholar] [CrossRef]
- Caddeo, A.; Kowalik, M.A.; Serra, M.; Runfola, M.; Bacci, A.; Rapposelli, S.; Columbano, A.; Perra, A. TG68, a Novel Thyroid Hormone Receptor-β Agonist for the Treatment of NAFLD. Int. J. Mol. Sci. 2021, 22, 13105. [Google Scholar] [CrossRef]
- Black, J.W.; Leff, P. Operational models of pharmacological agonism. Proc. R. Soc. Lond. B Biol. Sci. 1983, 220, 141–162. [Google Scholar] [CrossRef]
- Louisa, M.; Suyatna, F.D.; Wanandi, S.I.; Asih, P.B.; Syafruddin, D. Differential expression of several drug transporter genes in HepG2 and Huh-7 cell lines. Adv. Biomed. Res. 2016, 5, 104. [Google Scholar] [CrossRef]
- Sinha, R.A.; Singh, B.K.; Zhou, J.; Wu, Y.; Farah, B.L.; Ohba, K.; Lesmana, R.; Gooding, J.; Bay, B.-H.; Yen, P.M. Thyroid hormone induction of mitochondrial activity is coupled to mitophagy via ROS-AMPK-ULK1 signaling. Autophagy 2015, 11, 1341–1357. [Google Scholar] [CrossRef] [Green Version]
- Klein, I.; Ojamaa, K. Thyroid hormone and the cardiovascular system. N. Engl. J. Med. 2001, 344, 501–509. [Google Scholar] [CrossRef]
- Bassett, J.H.; Nordstrom, K.; Boyde, A.; Howell, P.G.; Kelly, S.; Vennstrom, B.; Williams, G.R. Thyroid status during skeletal development determines adult bone structure and mineralization. Mol. Endocrinol. 2007, 21, 1893–1904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seithel, A.; Eberl, S.; Singer, K.; Auge, D.; Heinkele, G.; Wolf, N.B.; Dörje, F.; Fromm, M.F.; König, J. The influence of macrolide antibiotics on the uptake of organic anions and drugs mediated by OATP1B1 and OATP1B3. Drug Metab. Dispos. Biol. Fate Chem. 2007, 35, 779–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konig, J.; Klatt, S.; Dilger, K.; Fromm, M.F. Characterization of ursodeoxycholic and norursodeoxycholic acid as substrates of the hepatic uptake transporters OATP1B1, OATP1B3, OATP2B1 and NTCP. Basic Clin. Pharmacol. Toxicol. 2012, 111, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Moeller, L.C.; Wardrip, C.; Niekrasz, M.; Refetoff, S.; Weiss, R.E. Comparison of thyroidectomized calf serum and stripped serum for the study of thyroid hormone action in human skin fibroblasts in vitro. Thyroid 2009, 19, 639–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuels, H.H.; Stanley, F.; Casanova, J. Depletion of L-3,5,3′-triiodothyronine and L-thyroxine in euthyroid calf serum for use in cell culture studies of the action of thyroid hormone. Endocrinology 1979, 105, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Echtermeyer, F.; Eberhardt, M.; Risser, L.; Herzog, C.; Gueler, F.; Khalil, M.; Engel, M.; Vondran, F.; Leffler, A. Acetaminophen-induced liver injury is mediated by the ion channel TRPV4. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 10257–10268. [Google Scholar] [CrossRef] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TR Isoform | Transporter | log(EC50) ± SD | EC50 [µM] | |
|---|---|---|---|---|
| T3 | TRα | OATP1B1 | −8.091 ± 0.043 | 0.007 |
| OATP1B3 | −8.441 ± 0.030 | 0.004 | ||
| TRβ | OATP1B1 | −8.059 ± 0.031 | 0.009 | |
| OATP1B3 | −8.356 ± 0.022 | 0.004 | ||
| MGL-3196 | TRα | OATP1B1 | - | - |
| OATP1B3 | - | - | ||
| TRβ | OATP1B1 | −6.221 ± 0.046 | 0.601 | |
| OATP1B3 | −4.753 ± 0.013 | 17.660 |
| Gene | Forward Primer 5′-->3′ | Reverse Primer 5′-->3′ | Accession No. | |
|---|---|---|---|---|
| human | 18S | CGG CTA CCA CAT CCA AGG AA | GCT GGA ATT ACC GCG GCT | NR_145820.1 |
| ACTB | AGA GCT ACG AGC TGC CTG AC | AGC ACT GTG TTG GCG TAC AG | NM_001101.5 | |
| PPIA | AAC GTG GTA TAA AAG GGG CGG | CTG CAA ACA GCT CAA AGG AGA C | NM_021130.5 | |
| SLCO1B1 | CAC TTG CAC TGG GTT TCC AC | AAG CCC AAG TAG ACC CTT GAA A | NM_006446.5 | |
| SLCO1B3 | TTT TTG GAA GGG TCT ACT TGG G | TCA TTG TCC GAT GCC TTG GTA | NM_019844.4 | |
| PCK-1 | GGA GAA GGA GGT GGAAGA C | GAA CAC TTG CCC TCT CTT GC | NM_002591.4 | |
| KLF9 | CTC CCA TCT CAA AGC CCA TTA C | TGA GCG GGA GAA CTT TTT AAG G | NM_001206.2 | |
| SLC16A2 | AGC TCC TTC ACC AGC TCC CTA AGC | TCC TCC ACA TAC TTC ATC AGG TG | NM_006517.5 | |
| SLC7A5 | GGG AAG GGT GAT GTG TCC AAT CTA | CAA GTA ATT CCA TCC TCC | NM_003486.7 | |
| SLC7A8 | GTG CTA TCA TCG TAG GGA AGA TC | CAC AGC CTC AGG AAC CCA G | NM_012244.4 | |
| murine | 18s | CGG CTA CCA CAT CCA AGG AA | GCT GGA ATT ACC GCG GCT | NR_003278.3 |
| Ppia | CTT GGG CCG CGT CTC CTT CG | GCG TGT AAA GTC ACC ACC CTG GC | NM_008907.2 | |
| Actb | GGC CCA GAG CAA GAG AGG TA | CTG GAT GGC TAC GTA CAT GGC | NM_007393.5 | |
| Rpl13a | GGG CAG GTT CTG GTA TTG GA | GGG GTT GGT ATT CAT CCG CT | NM_009438.5 | |
| Slco1b2 | ATC GGA CCA ATC CTT GGC TTT | TTA TGC GGA CAC TTC TCA GGT | NM_020495.2 | |
| Pck-1 | ATC TTT GGT GGC CGT AGA CC | ATC TTG CCC TTG TGT TCT GC | NM_011044.3 | |
| Dio1 | GGG CAG GAT CTG CTA CAA GG | CGT GTC TAG GTG GAG TGC AA | NM_007860.4 | |
| Serpina7 | TGG GCA TGT GCT ATC ATC TTC A | GAG TGG CAT TTT GTT GGG GC | NM_177920.5 | |
| Thrb | GGA CAA GCA CCC ATC GTG AA | ACA TGG CAG CTC ACA AAA CAT | NM_001113417.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hönes, G.S.; Sivakumar, R.G.; Hoppe, C.; König, J.; Führer, D.; Moeller, L.C. Cell-Specific Transport and Thyroid Hormone Receptor Isoform Selectivity Account for Hepatocyte-Targeted Thyromimetic Action of MGL-3196. Int. J. Mol. Sci. 2022, 23, 13714. https://doi.org/10.3390/ijms232213714
Hönes GS, Sivakumar RG, Hoppe C, König J, Führer D, Moeller LC. Cell-Specific Transport and Thyroid Hormone Receptor Isoform Selectivity Account for Hepatocyte-Targeted Thyromimetic Action of MGL-3196. International Journal of Molecular Sciences. 2022; 23(22):13714. https://doi.org/10.3390/ijms232213714
Chicago/Turabian StyleHönes, Georg Sebastian, Ramona Gowry Sivakumar, Christoph Hoppe, Jörg König, Dagmar Führer, and Lars Christian Moeller. 2022. "Cell-Specific Transport and Thyroid Hormone Receptor Isoform Selectivity Account for Hepatocyte-Targeted Thyromimetic Action of MGL-3196" International Journal of Molecular Sciences 23, no. 22: 13714. https://doi.org/10.3390/ijms232213714