
Prolyl Carboxypeptidase Activity Is Present in Human Adipose Tissue and Is Elevated in Serum of Obese Men with Type 2 Diabetes
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. PRCP Activity Is Higher in Adipose Tissue Compared to Skeletal Muscle Tissue
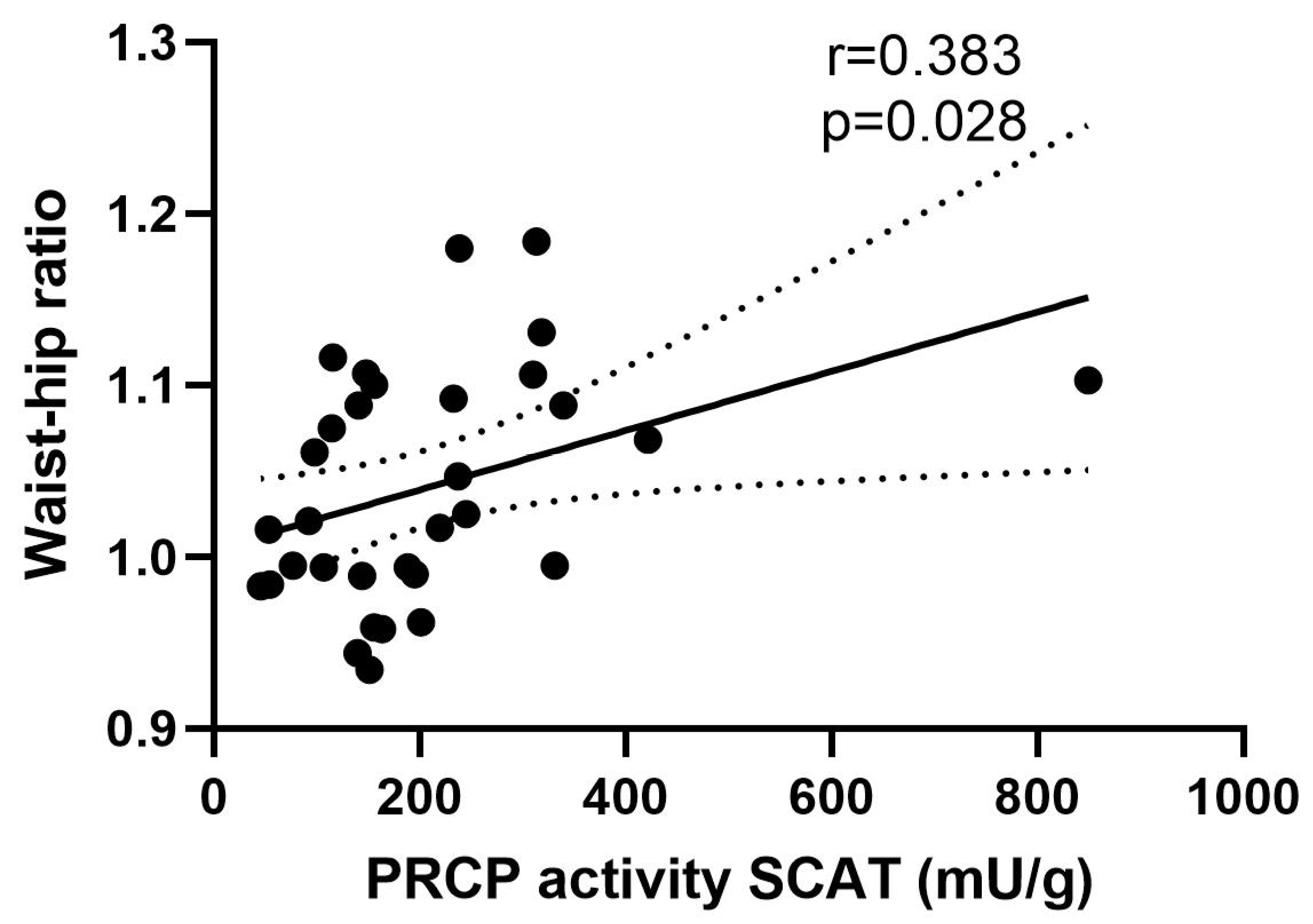
2.2. Serum PRCP Activity Is Elevated in Obese Men with T2D
2.3. PRCP Activity Is Positively Correlated with Immune Cell Populations
2.4. PRCP Activity Could Not Be Detected in Conditioned Media of Human Adipocytes
3. Discussion
4. Materials and Methods
4.1. Study Design
4.2. Anthropometric Measurements and Blood Sampling
4.3. Abdominal Subcutaneous and Visceral Adipose Tissue Biopsies
4.4. Tissue Lysate Preparation
4.5. PRCP Activity Measurement
4.6. Western Blot
4.7. Flow Cytometry
4.8. Adipocyte Size
4.9. Conditioned Media of Human Adipocytes
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wallingford, N.; Perroud, B.; Gao, Q.; Coppola, A.; Gyengesi, E.; Liu, Z.W.; Gao, X.B.; Diament, A.; Haus, K.A.; Shariat-Madar, Z.; et al. Prolylcarboxypeptidase regulates food intake by inactivating alpha-MSH in rodents. J. Clin. Investig. 2009, 119, 2291–2303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kehoe, K.; Van Elzen, R.; Verkerk, R.; Sim, Y.; Van der Veken, P.; Lambeir, A.-M.; De Meester, I. Prolyl carboxypeptidase purified from human placenta: Its characterization and identification as an apelin-cleaving enzyme. Biochim. Biophys. Acta Proteins Proteom. 2016, 1864, 1481–1488. [Google Scholar] [CrossRef] [PubMed]
- De Hert, E.; Bracke, A.; Pintelon, I.; Janssens, E.; Lambeir, A.M.; Van Der Veken, P.; De Meester, I. Prolyl carboxypeptidase mediates the C-terminal cleavage of (Pyr)-apelin-13 in human umbilical vein and aortic endothelial cells. Int. J. Mol. Sci. 2021, 22, 6698. [Google Scholar] [CrossRef] [PubMed]
- Coll, A.P.; Farooqi, I.S.; Challis, B.G.; Yeo, G.S.H.; O’Rahilly, S. Proopiomelanocortin and energy balance: Insights from human and murine genetics. J. Clin. Endocrinol. Metab. 2004, 89, 2557–2562. [Google Scholar] [CrossRef] [Green Version]
- Tatemoto, K.; Hosoya, M.; Habata, Y.; Fujii, R.; Kakegawa, T.; Zou, M.X.; Kawamata, Y.; Fukusumi, S.; Hinuma, S.; Kitada, C.; et al. Isolation and characterization of a novel endogenous peptide ligand for the human APJ receptor. Biochem. Biophys. Res. Commun. 1998, 251, 471–476. [Google Scholar] [CrossRef]
- Shin, K.; Kenward, C.; Rainey, J.K. Apelinergic system structure and function. Compr. Physiol. 2018, 8, 407–450. [Google Scholar] [CrossRef]
- Wysocka, M.B.; Pietraszek-Gremplewicz, K.; Nowak, D. The role of apelin in cardiovascular diseases, obesity and cancer. Front. Physiol. 2018, 9, 557. [Google Scholar] [CrossRef]
- Chaves-Almagro, C.; Castan-Laurell, I.; Dray, C.; Knauf, C.; Valet, P.; Masri, B. Apelin receptors: From signaling to antidiabetic strategy. Eur. J. Pharmacol. 2015, 763, 149–159. [Google Scholar] [CrossRef]
- Marsault, E.; Llorens-Cortes, C.; Iturrioz, X.; Chun, H.J.; Lesur, O.; Oudit, G.Y.; Auger-Messier, M. The apelinergic system: A perspective on challenges and opportunities in cardiovascular and metabolic disorders. Ann. N. Y. Acad. Sci. 2019, 1455, 12–33. [Google Scholar] [CrossRef]
- Bruschetta, G.; Jin, S.; Kim, J.D.; Diano, S. Prolyl carboxypeptidase in Agouti-related Peptide neurons modulates food intake and body weight. Mol. Metab. 2018, 10, 28–38. [Google Scholar] [CrossRef]
- Bruschetta, G.; Jin, S.; Liu, Z.W.; Kim, J.D.; Diano, S. MC4R Signaling in Dorsal Raphe Nucleus Controls Feeding, Anxiety, and Depression. Cell Rep. 2020, 33, 108267. [Google Scholar] [CrossRef] [PubMed]
- Graham, T.H. Prolylcarboxypeptidase (PrCP) inhibitors and the therapeutic uses thereof: A patent review. Expert Opin. Ther. Pat. 2017, 27, 1077–1088. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.K.; Diano, S. Prolyl carboxypeptidase and its inhibitors in metabolism. Trends Endocrinol. Metab. 2013, 24, 61–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Lind, L.; Zhao, L.; Lindahl, B.; Venge, P. Plasma prolylcarboxypeptidase (Angiotensinase C) is increased in obesity and diabetes mellitus and related to cardiovascular dysfunction. Clin. Chem. 2012, 58, 1110–1115. [Google Scholar] [CrossRef] [Green Version]
- Kehoe, K.; Noels, H.; Theelen, W.; De Hert, E.; Xu, S.; Verrijken, A.; Arnould, T.; Fransen, E.; Hermans, N.; Lambeir, A.; et al. Prolyl carboxypeptidase activity in the circulation and its correlation with body weight and adipose tissue in lean and obese subjects. PLoS ONE 2018, 13, e0197603. [Google Scholar]
- Tabrizian, T.; Hataway, F.; Murray, D.; Shariat-Madar, Z. Prolylcarboxypeptidase Gene Expression in the Heart and Kidney: Effects of Obesity and Diabetes. Cardiovasc. Hematol. Agents Med. Chem. 2015, 13, 113–123. [Google Scholar] [CrossRef]
- Kumamoto, K.; Stewart, T.A.; Johnson, A.R.; Erdos, E.G. Prolylcarboxypeptidase (angiotensinase C) in human lung and cultured cells. J. Clin. Investig. 1981, 67, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Kehoe, K.; Brouns, R.; Verkerk, R.; Engelborghs, S.; De Deyn, P.P.; Hendriks, D.; De Meester, I. Prolyl carboxypeptidase activity decline correlates with severity and short-term outcome in acute ischemic stroke. Neurochem. Res. 2015, 40, 81–88. [Google Scholar] [CrossRef]
- Vliegen, G.; Kehoe, K.; Bracke, A.; de Hert, E.; Verkerk, R.; Fransen, E.; Jongers, B.; Peters, E.; Lambeir, A.M.; Kumar-Singh, S.; et al. Dysregulated activities of proline-specific enzymes in septic shock patients (sepsis-2). PLoS ONE 2020, 15, e0231555. [Google Scholar] [CrossRef] [Green Version]
- Ngo, M.-L.; Mahdi, F.; Kolte, D.; Shariat-Madar, Z. Upregulation of prolylcarboxypeptidase (PRCP) in lipopolysaccharide (LPS) treated endothelium promotes inflammation. J. Inflamm. 2009, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Goossens, G.H.; Jocken, J.W.E.; Blaak, E.E. Sexual dimorphism in cardiometabolic health: The role of adipose tissue, muscle and liver. Nat. Rev. Endocrinol. 2021, 17, 47–66. [Google Scholar] [CrossRef]
- Verboven, K.; Hansen, D.; Moro, C.; Eijnde, B.O.; Hoebers, N.; Knol, J.; Bouckaert, W.; Dams, A.; Blaak, E.E.; Jocken, J.W.E. Attenuated atrial natriuretic peptide-mediated lipolysis in subcutaneous adipocytes of obese type 2 diabetic men. Clin. Sci. 2016, 130, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Verboven, K.; Wouters, K.; Gaens, K.; Hansen, D.; Bijnen, M.; Wetzels, S.; Stehouwer, C.D.; Goossens, G.H.; Schalkwijk, C.G.; Blaak, E.E.; et al. Abdominal subcutaneous and visceral adipocyte size, lipolysis and inflammation relate to insulin resistance in male obese humans. Sci. Rep. 2018, 8, 4677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kehoe, K.; Verkerk, R.; Sim, Y.; Waumans, Y.; Van Der Veken, P.; Lambeir, A.M.; De Meester, I. Validation of a specific prolylcarboxypeptidase activity assay and its suitability for plasma and serum measurements. Anal. Biochem. 2013, 443, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Wouters, K.; Gaens, K.; Bijnen, M.; Verboven, K.; Jocken, J.; Wetzels, S.; Wijnands, E.; Hansen, D.; Van Greevenbroek, M.; Duijvestijn, A.; et al. Circulating classical monocytes are associated with CD11c+ macrophages in human visceral adipose tissue. Sci. Rep. 2017, 7, 42665. [Google Scholar] [CrossRef] [Green Version]
- Goossens, G.H.; Bizzarri, A.; Venteclef, N.; Essers, Y.; Cleutjens, J.P.; Konings, E.; Jocken, J.W.E.; Merima, C.; Blaak, E.E. Increased Adipose Tissue Oxygen Tension in Obese Compared with Lean Men Is Accompanied by Insulin Resistance, Impaired Adipose Tissue Capillarization, and Inflammation. Circulation 2011, 124, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Jocken, J.W.E.; Goossens, G.H.; Popeijus, H.; Essers, Y.; Hoebers, N.; Blaak, E.E. Contribution of lipase deficiency to mitochondrial dysfunction and insulin resistance in hMADS adipocytes. Int. J. Obes. 2016, 40, 507–513. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Lean Individuals | Obese, Non-Diabetic Individuals | Obese, Diabetic Individuals |
|---|---|---|---|
| n | 19 | 25 | 11 |
| Age (years) | 52 ± 6 | 50 ± 7 | 52 ± 7 |
| Length (m) | 1.80 ± 0.07 | 1.80 ± 0.07 | 1.76 ± 0.04 |
| Weight (kg) | 78 ± 7 | 124 ± 15 | 117 ± 9 |
| BMI (kg/m2) | 24 ± 1 | 38 ± 4 | 38 ± 4 |
| Waist circumference (cm) | 91 ± 5 | 129 ± 10 | 126 ± 7 |
| Hip circumference (cm) | 94 ± 4 | 117 ± 9 | 116 ± 7 |
| Waist-to-hip ratio | 0.98 ± 0.03 | 1.09 ± 0.04 | 1.09 ± 0.06 |
| Systolic blood pressure (mmHg) | 125 ± 15 | 143 ± 14 | 147 ± 15 |
| Diastolic blood pressure (mmHg) | 80 ± 9 | 84 ± 8 | 85 ± 10 |
| Fasting glucose (mmol/L) | 5.7 ± 0.8 | 6 ± 1 | 8 ± 2 |
| Serum insulin (mU/L) | 7 ± 4 | 21 ± 10 | 30 ± 31 |
| HOMA-IR | 2 ± 1 | 6 ± 3 | 12 ± 13 |
| HbA1c (%) | 5.2 ± 0.2 | 5.8 ± 0.7 | 7.0 ± 1.0 |
| HbA1c (mmol/mol) | 33 ± 3 | 40 ± 8 | 56 ± 11 |
| Fat percentage (%) | 23 ± 4 | 37 ± 5 | 37 ± 3 |
| Fat mass (kg) | 18 ± 4 | 46 ± 11 | 43 ± 7 |
| Fat free mass (kg) | 58 ± 6 | 76 ± 7 | 74 ± 4 |
| NEFA (µmol/L) | 244 ± 169 | 759 ± 319 | 872 ± 301 |
| Triacylglycerols (mmol/L) | 907 ± 587 | 1274 ± 407 | 1271 ± 524 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Hert, E.; Verboven, K.; Wouters, K.; Jocken, J.W.E.; De Meester, I. Prolyl Carboxypeptidase Activity Is Present in Human Adipose Tissue and Is Elevated in Serum of Obese Men with Type 2 Diabetes. Int. J. Mol. Sci. 2022, 23, 13529. https://doi.org/10.3390/ijms232113529
De Hert E, Verboven K, Wouters K, Jocken JWE, De Meester I. Prolyl Carboxypeptidase Activity Is Present in Human Adipose Tissue and Is Elevated in Serum of Obese Men with Type 2 Diabetes. International Journal of Molecular Sciences. 2022; 23(21):13529. https://doi.org/10.3390/ijms232113529
Chicago/Turabian StyleDe Hert, Emilie, Kenneth Verboven, Kristiaan Wouters, Johan W. E. Jocken, and Ingrid De Meester. 2022. "Prolyl Carboxypeptidase Activity Is Present in Human Adipose Tissue and Is Elevated in Serum of Obese Men with Type 2 Diabetes" International Journal of Molecular Sciences 23, no. 21: 13529. https://doi.org/10.3390/ijms232113529