
Computer-Aided Screening and Revealing Action Mechanism of Food-Derived Tripeptides Intervention in Acute Colitis
Abstract
:1. Introduction
2. Results
2.1. Network Interaction Analysis
2.2. Hub Targets GO Biological Functions Enrichments
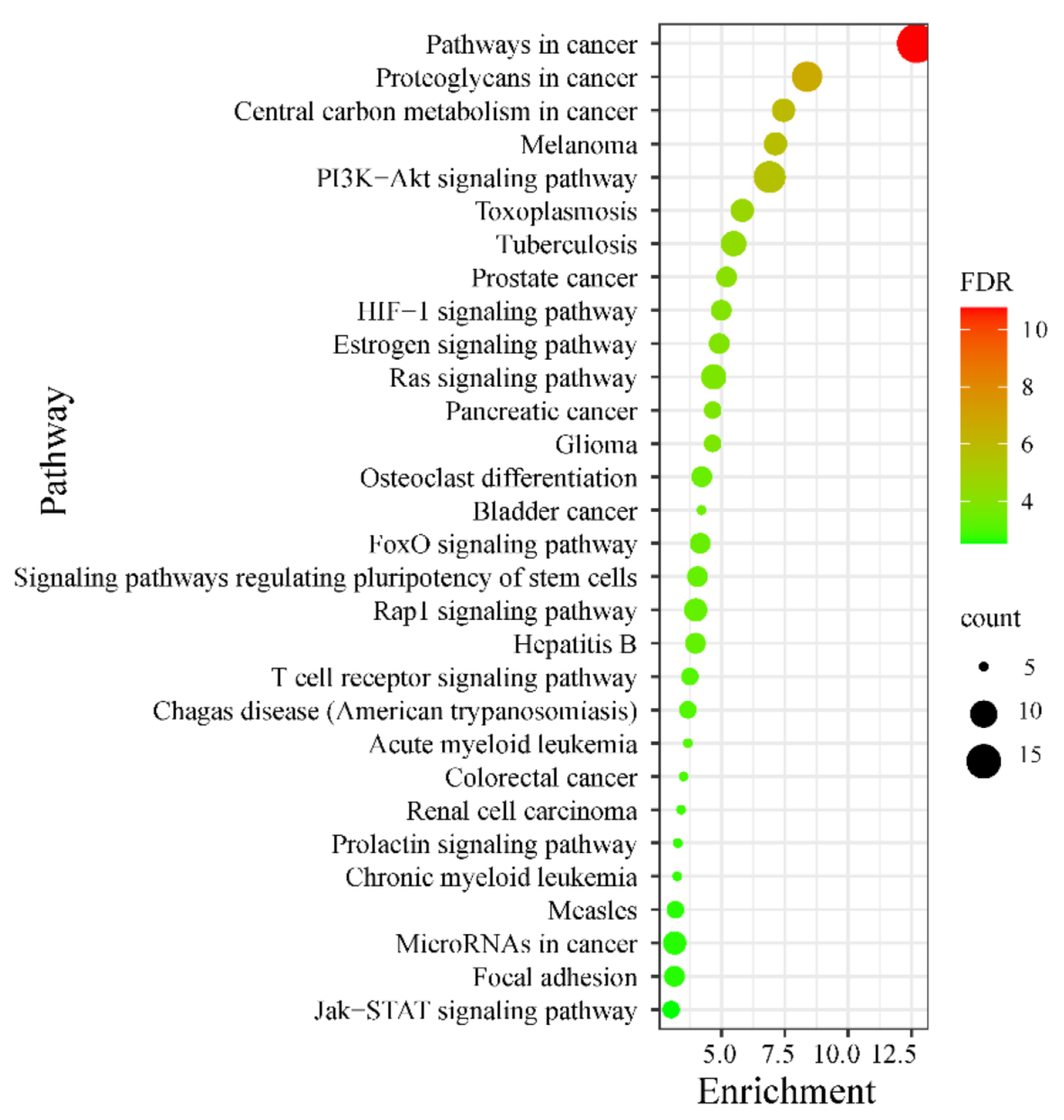
2.3. Hub Targets KEGG Pathway Enrichments
2.4. Expression of Hub Targets in Organs Site
2.5. Molecular Docking Results of Tripeptide and Hub Target
3. Discussion
4. Material and Methods
4.1. Construction of Tripeptide Set and Prediction of Colitis-Related Targets
4.2. Construction of Protein-Protein Interaction (PPI) Network
4.3. Analysis of GO and KEGG Pathway Enrichment
4.4. Hub Target’s Expression Organs Site
4.5. Molecular Docking
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hu, B.; Yu, S.; Shi, C.; Gu, J.; Shao, Y.; Chen, Q.; Li, Y.; Mezzenga, R. Amyloid–polyphenol hybrid nanofilaments mitigate colitis and regulate gut microbial dysbiosis. ACS Nano 2020, 14, 2760–2776. [Google Scholar] [CrossRef] [PubMed]
- Ge, H.F.; Zhang, B.Y.; Li, T.; Yu, Y.; Men, F.B.; Zhao, S.N.; Liu, J.B.; Zhang, T. Potential targets and the action mechanism of food-derived dipeptides on colitis: Network pharmacology and bioinformatics analysis. Food Funct. 2021, 12, 5989–6000. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, G.G. The global burden of IBD: From 2015 to 2025. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Gao, Y.; Wang, L.; Yang, D.; Bu, W.; Gou, L.; Huang, J.; Duan, X.; Pan, Y.; Cao, S.; et al. Troxerutin improves dextran sulfate sodium-induced ulcerative colitis in mice. J. Agric. Food Chem. 2021, 69, 2729–2744. [Google Scholar] [CrossRef] [PubMed]
- Ge, H.; Zhang, B.; Li, T.; Yang, Q.; Tang, Y.; Liu, J.; Zhang, T. In vivo and in silico studies on the mechanisms of egg white peptides in relieving acute colitis symptoms. Food Funct. 2021, 12, 12774–12787. [Google Scholar] [CrossRef] [PubMed]
- Ungaro, R.; Mehandru, S.; Allen, P.B.; Peyrin-Biroulet, L.; Colombel, J.-F. Ulcerative colitis. Lancet 2017, 389, 1756–1770. [Google Scholar] [CrossRef]
- Zhang, J.; Fu, X.; Li, W.; Li, H.; Ying, Z.; Liu, X.; Yin, L. Enhancement of nutritional soy protein and peptide supplementation on skin repair in rats. J. Funct. Foods 2020, 75, 104231. [Google Scholar] [CrossRef]
- Ge, H.; Cai, Z.; Chai, J.; Liu, J.; Liu, B.; Yu, Y.; Liu, J.; Zhang, T. Egg white peptides ameliorate dextran sulfate sodium-induced acute colitis symptoms by inhibiting the production of pro-inflammatory cytokines and modulation of gut microbiota composition. Food Chem. 2021, 360, 129981. [Google Scholar] [CrossRef]
- Shi, W.; Hou, T.; Guo, D.; He, H. Evaluation of hypolipidemic peptide (Val-Phe-Val-Arg-Asn) virtual screened from chickpea peptides by pharmacophore model in high-fat diet-induced obese rat. J. Funct. Foods 2019, 54, 136–145. [Google Scholar] [CrossRef]
- Liu, C.; Liu, J.; Wang, M.; Zhang, B.; Wang, E.; Liu, B.; Zhang, T. Construction and application of membrane-bound angiotensin-i converting enzyme system: A new approach for the evaluation of angiotensin-i converting enzyme inhibitory peptides. J. Agric. Food Chem. 2020, 68, 5723–5731. [Google Scholar] [CrossRef]
- Wen, H.; Li, Z.; Li, Y.; Hao, Y.; Du, Z.; Liu, X.; Shang, X.; Liu, J.; Zhang, T. Aggregation of egg white peptides (EWP) induced by proanthocyanidins: A promising fabrication strategy for EWP emulsion. Food Chem. 2022, 400, 134019. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.Y.; Zhao, Y.; Wu, N.; Yao, Y.; Xu, M.S.; Du, H.Y.; Tu, Y.G. The anti-inflammatory activity of peptides from simulated gastrointestinal digestion of preserved egg white in DSS-induced mouse colitis. Food Funct. 2018, 9, 6445–6455. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Ge, Q.; Du, H.; Jiang, P.; Bao, Z.; Chen, D.; Lin, S. Potential mechanisms mediating the protective effects of tricholoma matsutake-derived peptides in mitigating dss-induced colitis. J. Agric. Food Chem. 2021, 69, 5536–5546. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.; Shao, A.; Li, H.; Tang, Y.; Li, Q.; Guo, Z.; Wu, C.; Cheng, Y.; Tian, H.; Zhu, W.-H. Peptide receptor-targeted fluorescent probe: Visualization and discrimination between chronic and acute ulcerative colitis. ACS Appl. Mater. 2017, 9, 13029–13036. [Google Scholar] [CrossRef] [PubMed]
- Xiao, B.; Xu, Z.; Viennois, E.; Zhang, Y.; Zhang, Z.; Zhang, M.; Han, M.K.; Kang, Y.; Merlin, D. Orally targeted delivery of tripeptide kpv via hyaluronic acid-functionalized nanoparticles efficiently alleviates ulcerative colitis. Mol. Ther. 2017, 25, 1628–1640. [Google Scholar] [CrossRef] [Green Version]
- Deng, Z.; Ni, J.; Wu, X.; Wei, H.; Peng, J. GPA peptide inhibits NLRP3 inflammasome activation to ameliorate colitis through AMPK pathway. Aging 2020, 12, 18522–18544. [Google Scholar] [CrossRef]
- Wang, J.S.; Gong, X.F.; Feng, J.L.; Li, H.S.; Li, X.; Deng, S.; Ren, P.Z.; Wang, J.M.; Lv, M.S.; Jin, R.F.; et al. Study on the mechanism of Jiawei Shengjiang powder in improving male asthma-induced asthenospermia based on network pharmacology and bioinformatics. Drug. Des. Dev. Ther. 2021, 15, 1245–1259. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Fumery, M.; Dulai, P.S.; Prokop, L.J.; Sandborn, W.J.; Murad, M.H.; Singh, S. Comparative efficacy and tolerability of pharmacological agents for management of mild to moderate ulcerative colitis: A systematic review and network meta-analyses. Lancet Gastroenterol. 2018, 3, 742–753. [Google Scholar] [CrossRef]
- Duan, H.; Khan, G.J.; Shang, L.-j.; Peng, H.; Hu, W.-C.; Zhang, J.-Y.; Hua, J.; Cassandra, A.; Rashed, M.M.A.; Zhai, K.-F. Computational pharmacology and bioinformatics to explore the potential mechanism of Schisandra against atherosclerosis. Food Chem. Toxicol. 2021, 150, 112058. [Google Scholar] [CrossRef]
- Whaley, K.G.; Rosen, M.J. Contemporary medical management of acute severe ulcerative colitis. Inflamm. Bowel. Dis. 2019, 25, 56–66. [Google Scholar] [CrossRef]
- Pineton de Chambrun, G.; Peyrin-Biroulet, L.; Lémann, M.; Colombel, J.-F. Clinical implications of mucosal healing for the management of IBD. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.L.; Xu, J.; Zhang, X.H.; Qiu, B.Y.; Peng, L.; Zhang, M.; Gan, H.T. PI3K/Akt signaling pathway is involved in the pathogenesis of ulcerative colitis. Inflamm. Res. 2011, 60, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Son, Y.S.; Lee, M.O.; Ryu, J.W.; Park, K.; Kwon, O.; Jung, K.B.; Kim, K.; Ryu, T.Y.; Baek, A.; et al. Low-dose interleukin-2 alleviates dextran sodium sulfate-induced colitis in mice by recovering intestinal integrity and inhibiting AKT-dependent pathways. Theranostics 2020, 10, 5048–5063. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; Kim, H.G.; Kim, E.; Han, S.Y.; Aziz, N.; Yi, Y.S.; Kim, S.; Lee, Y.; Yoo, B.C.; Han, J.W.; et al. Isoprenylcysteine carboxyl methyltransferase and its substrate ras are critical players regulating tlr-mediated inflammatory responses. Cells 2020, 9, 1216. [Google Scholar] [CrossRef]
- Lu, N.; Wang, L.; Cao, H.; Liu, L.; Van Kaer, L.; Washington, M.K.; Rosen, M.J.; Dubé, P.E.; Wilson, K.T.; Ren, X.; et al. Activation of the epidermal growth factor receptor in macrophages regulates cytokine production and experimental colitis. J. Immunol. 2014, 192, 1013. [Google Scholar] [CrossRef] [Green Version]
- Belle, N.M.; Ji, Y.; Herbine, K.; Wei, Y.; Park, J.; Zullo, K.; Hung, L.-Y.; Srivatsa, S.; Young, T.; Oniskey, T.; et al. TFF3 interacts with LINGO2 to regulate EGFR activation for protection against colitis and gastrointestinal helminths. Nat. Commun. 2019, 10, 4408. [Google Scholar] [CrossRef] [Green Version]
- Yarden, Y.; Sliwkowski, M.X. Untangling the ErbB signalling network. Nat. Rev. Mol. Cell Biol. 2001, 2, 127–137. [Google Scholar] [CrossRef]
- Dubé, P.E.; Yan, F.; Punit, S.; Girish, N.; McElroy, S.J.; Washington, M.K.; Polk, D.B. Epidermal growth factor receptor inhibits colitis-associated cancer in mice. J. Clin. Investig. 2012, 122, 2780–2792. [Google Scholar] [CrossRef]
- Sternlicht, M.D.; Werb, Z. How matrix metalloproteinases regulate cell behavior. Annu. Rev. Cell Dev. Biol. 2001, 17, 463–516. [Google Scholar] [CrossRef] [Green Version]
- Walter, L.; Canup, B.; Pujada, A.; Bui, T.A.; Arbasi, B.; Laroui, H.; Merlin, D.; Garg, P. Matrix metalloproteinase 9 (MMP9) limits reactive oxygen species (ROS) accumulation and DNA damage in colitis-associated cancer. Cell Death Dis. 2020, 11, 767. [Google Scholar] [CrossRef]
- Walter, L.; Pujada, A.; Bhatnagar, N.; Bialkowska, A.B.; Yang, V.W.; Laroui, H.; Garg, P. Epithelial derived-matrix metalloproteinase (MMP9) exhibits a novel defensive role of tumor suppressor in colitis associated cancer by activating MMP9-Notch1-ARF-p53 axis. Oncotarget 2017, 8, 364–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Fan, P.; Chou, H.; Li, J.; Wang, K.; Li, H. Herbacetin suppressed MMP9 mediated angiogenesis of malignant melanoma through blocking EGFR-ERK/AKT signaling pathway. Biochimie 2019, 162, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Kesari, P.; Pratap, S.; Dhankhar, P.; Dalal, V.; Mishra, M.; Singh, P.K.; Chauhan, H.; Kumar, P. Structural characterization and in-silico analysis of Momordica charantia 7S globulin for stability and ACE inhibition. Sci. Rep. 2020, 10, 1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Ouyang, S.; Yu, B.; Liu, Y.; Huang, K.; Gong, J.; Zheng, S.; Li, Z.; Li, H.; Jiang, H. PharmMapper server: A web server for potential drug target identification using pharmacophore mapping approach. Nucleic Acids Res. 2010, 38, W609–W614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saunders, L.R.; Bankovich, A.J.; Anderson, W.C.; Aujay, M.A.; Bheddah, S.; Black, K.; Desai, R.; Escarpe, P.A.; Hampl, J.; Laysang, A.; et al. A DLL3-targeted antibody-drug conjugate eradicates high-grade pulmonary neuroendocrine tumor-initiating cells in vivo. Sci. Transl. Med. 2015, 7, 302ra136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Wu, S.; Tong, L.; Liu, B.; Ai, Z.; Hong, Z.; You, P.; Wu, H.; Yang, Y. Bioactive ingredients obtained from Cortex Fraxini impair interactions between FAS and GPI. Free Radical. Bio. Med. 2020, 152, 504–515. [Google Scholar] [CrossRef]
- Xu, W.; Zhang, Y.; Yu, Y.; Li, B.; Liu, J.; Wang, P.; Wu, H.; Liu, Q.; Wei, Z.; Xiao, H.; et al. Dose-dependent target diversion of Danhong injection on the Glu-GLT-1/Gly-GlyRalpha dynamic balance module of cerebral ischemia. Pharmacol. Res. 2018, 135, 80–88. [Google Scholar] [CrossRef]
- Dennis, G.; Sherman, B.T.; Hosack, D.A.; Yang, J.; Gao, W.; Lane, H.C.; Lempicki, R.A. DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome Biol. 2003, 4, R60. [Google Scholar] [CrossRef] [Green Version]
- Pathan, M.; Keerthikumar, S.; Ang, C.S.; Gangoda, L.; Quek, C.Y.; Williamson, N.A.; Mouradov, D.; Sieber, O.M.; Simpson, R.J.; Salim, A.; et al. FunRich: An open access standalone functional enrichment and interaction network analysis tool. Proteomics 2015, 15, 2597–2601. [Google Scholar] [CrossRef]
- Pathan, M.; Keerthikumar, S.; Chisanga, D.; Alessandro, R.; Ang, C.S.; Askenase, P.; Batagov, A.O.; Benito-Martin, A.; Camussi, G.; Clayton, A.; et al. A novel community driven software for functional enrichment analysis of extracellular vesicles data. J. Extracell. Vesicles 2017, 6, 1321455. [Google Scholar] [CrossRef] [Green Version]
- Sanner, M.F. Python: A programming language for software integration and development. J. Mol. Graph. Model. 1999, 17, 57–61. [Google Scholar] [PubMed]
- Milburn, C.C.; Deak, M.; Kelly, S.M.; Price, N.C.; Alessi, D.R.; Aalten, D.M.F.V. Binding of phosphatidylinositol 3,4,5-trisphosphate to the pleckstrin homology domain of protein kinase B induces a conformational change. Biochem. J. 2003, 375, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Kawakita, Y.; Seto, M.; Ohashi, T.; Tamura, T.; Yusa, T.; Miki, H.; Iwata, H.; Kamiguchi, H.; Tanaka, T.; Sogabe, S.; et al. Design and synthesis of novel pyrimido[4,5-b]azepine derivatives as HER2/EGFR dual inhibitors. Bioorganic Med. Chem. 2013, 21, 2250–2261. [Google Scholar] [CrossRef] [PubMed]
- Antoni, C.; Vera, L.; Devel, L.; Catalani, M.P.; Czarny, B.; Cassar-Lajeunesse, E.; Nuti, E.; Rossello, A.; Dive, V.; Stura, E.A. Crystallization of bi-functional ligand protein complexes. J. Struct. Biol. 2013, 182, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The protein data bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Liu, J.; Liu, C.; Liu, B.; Yu, Y.; Zhang, T. Bifunctional peptides with antioxidant and angiotensin-converting enzyme inhibitory activity in vitro from egg white hydrolysates. J. Food Biochem. 2020, 44, e13347. [Google Scholar] [CrossRef]
- Momanyt, F.A.; Rone, R. Validation of the General Purpose QUANTAa3.2/CHARMma Force Field. J. Comput. Chem. 1992, 13, 888–900. [Google Scholar] [CrossRef]
- Dalal, V.; Golemi-Kotra, D.; Kumar, P. Quantum mechanics/molecular mechanics studies on the catalytic mechanism of a novel esterase (fmta) of Staphylococcus aureus. J. Chem. Inf. Model. 2022, 62, 2409–2420. [Google Scholar] [CrossRef]
- Wallace, A.C.; Laskowski, R.A.; Thornton, J.M. LIGPLOT: A program to generate schematic diagrams of protein-ligand interactions. Protein Eng. 1995, 8, 127–134. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Gene | Protein Target | UniProt ID |
|---|---|---|---|
| 1 | A1AT | Alpha-1-antitrypsin | P01009 |
| 2 | ACE | Angiotensin-converting enzyme | P12821 |
| 3 | AKT1 | RAC-alpha serine/threonine-protein kinase | P31749 |
| 4 | AKT2 | RAC-beta serine/threonine-protein kinase | P31751 |
| 5 | BTK | Tyrosine-protein kinase BTK | Q06187 |
| 6 | C1S | Complement C1s subcomponent | P09871 |
| 7 | CASP3 | Caspase-3 | P42574 |
| 8 | EGFR | Epidermal growth factor receptor | P00533 |
| 9 | ELNE | Neutrophil elastase | P08246 |
| 10 | FGFR1 | Fibroblast growth factor receptor 1 | P11362 |
| 11 | G6PD | Glucose-6-phosphate 1-dehydrogenase | P11413 |
| 12 | GALK1 | Galactokinase | P51570 |
| 13 | GCR | Glucocorticoid receptor | P04150 |
| 14 | GSTM1 | Glutathione S-transferase Mu 1 | P09488 |
| 15 | GSTP1 | Glutathione S-transferase P | P09211 |
| 16 | HMOX1 | Heme oxygenase 1 | P09601 |
| 17 | IGF1R | Insulin-like growth factor 1 receptor | P08069 |
| 18 | IL2 | Interleukin-2 | P60568 |
| 19 | JAK2 | Tyrosine-protein kinase JAK2· | O60674 |
| 20 | JAK3 | Tyrosine-protein kinase JAK3 | P52333 |
| 21 | KIT | Mast/stem cell growth factor receptor Kit | P10721 |
| 22 | LCK | Interleukin-2 | P06239 |
| 23 | MDM2 | E3 ubiquitin-protein ligase Mdm2 | Q00987 |
| 24 | MET | Hepatocyte growth factor receptor | P08581 |
| 25 | MIF | Macrophage migration inhibitory factor | P14174 |
| 26 | MMP13 | Collagenase 3 | P45452 |
| 27 | MMP2 | 72 kDa type IV collagenase | P08253 |
| 28 | MMP9 | Matrix metalloproteinase-9 | P14780 |
| 29 | NOS2 | Nitric oxide synthase, inducible | P35228 |
| 30 | NOS3 | Nitric oxide synthase, endothelial | P29474 |
| 31 | PPARG | Peroxisome proliferator-activated receptor gamma | P37231 |
| 32 | RAF1 | RAF proto-oncogene serine/threonine-protein kinase | P04049 |
| 33 | RARA | Retinoic acid receptor alpha | P10276 |
| 34 | SAHH | Adenosylhomocysteinase | P23526 |
| 35 | STAT1 | Signal transducer and activator of transcription 1-alpha/beta | P42224 |
| 36 | TGFB2 | Transforming growth factor beta-2 proprotein | P61812 |
| 37 | TGFR1 | TGF-beta receptor type-1 | P36897 |
| 38 | TGM2 | Protein-glutamine gamma-glutamyltransferase 2 | P21980 |
| 39 | VDR | Vitamin D3 receptor | P11473 |
| 40 | XIAP | E3 ubiquitin-protein ligase XIAP | P98170 |
| 41 | ZAP70 | Tyrosine-protein kinase ZAP-70 | P43403 |
| No. | Name | Betweenness Centrality | Closeness Centrality | Clustering Coefficient | Degree |
|---|---|---|---|---|---|
| 1 | AKT1 | 0.1690 | 0.8000 | 0.3471 | 30 |
| 2 | EGFR | 0.1477 | 0.7547 | 0.3276 | 29 |
| 3 | MMP9 | 0.0949 | 0.7143 | 0.4275 | 24 |
| 4 | CASP3 | 0.0423 | 0.6897 | 0.4819 | 24 |
| 5 | MMP2 | 0.0854 | 0.6349 | 0.4211 | 20 |
| 6 | IL2 | 0.0326 | 0.6250 | 0.4795 | 19 |
| 7 | STAT1 | 0.0221 | 0.6061 | 0.5294 | 18 |
| 8 | JAK2 | 0.0269 | 0.6250 | 0.4902 | 18 |
| 9 | PPARG | 0.0093 | 0.5882 | 0.6286 | 15 |
| 10 | HMOX1 | 0.0354 | 0.5797 | 0.5055 | 14 |
| 11 | NOS3 | 0.0094 | 0.5797 | 0.6154 | 14 |
| 12 | MDM2 | 0.0139 | 0.5797 | 0.5165 | 14 |
| 13 | LCK | 0.0094 | 0.5634 | 0.6264 | 14 |
| 14 | KIT | 0.0093 | 0.5634 | 0.5897 | 13 |
| 15 | AKT2 | 0.0066 | 0.5797 | 0.6795 | 13 |
| 16 | NOS2 | 0.0041 | 0.5479 | 0.6970 | 12 |
| 17 | IGF1R | 0.0060 | 0.5714 | 0.6970 | 12 |
| 18 | XIAP | 0.0658 | 0.5556 | 0.5636 | 11 |
| 19 | GCR | 0.0034 | 0.5405 | 0.6727 | 11 |
| 20 | RAF1 | 0.0036 | 0.5556 | 0.7273 | 11 |
| 21 | ACE | 0.0023 | 0.5479 | 0.8000 | 10 |
| 22 | VDR | 0.0032 | 0.5263 | 0.6111 | 9 |
| 23 | TGM2 | 0.0070 | 0.5479 | 0.7143 | 8 |
| 24 | TGFB2 | 0.0073 | 0.5405 | 0.7500 | 8 |
| 25 | JAK3 | 0.0007 | 0.5195 | 0.8214 | 8 |
| 26 | BTK | 0.0025 | 0.5195 | 0.7500 | 8 |
| 27 | RARA | 0.0008 | 0.5128 | 0.7619 | 7 |
| 28 | MMP13 | 0.0007 | 0.5195 | 0.8571 | 7 |
| 29 | FGFR1 | 0.0009 | 0.5195 | 0.8095 | 7 |
| 30 | ALAT | 0.0994 | 0.5195 | 0.3810 | 7 |
| 31 | MET | 0.0004 | 0.5195 | 0.9048 | 7 |
| 32 | GSTP1 | 0.0128 | 0.5263 | 0.4667 | 6 |
| 33 | ZAP70 | 0.0003 | 0.4819 | 0.9000 | 5 |
| 34 | MIF | 0.0000 | 0.4938 | 1.0000 | 4 |
| 35 | G6PD | 0.0001 | 0.5000 | 0.8333 | 4 |
| 36 | GSTM1 | 0.0015 | 0.4301 | 0.3333 | 3 |
| 37 | TGFR1 | 0.0000 | 0.4598 | 1.0000 | 2 |
| 38 | SAHH | 0.0000 | 0.3604 | 0.0000 | 1 |
| 39 | C1S | 0.0000 | 0.3448 | 0.0000 | 1 |
| 40 | ELNE | 0.0000 | 0.3922 | 0.0000 | 1 |
| 41 | GALK1 | 0.0000 | 0.3448 | 0.0000 | 1 |
| Ligands | AKT1 | EGFR | MMP9 | Ligands | AKT1 | EGFR | MMP9 |
|---|---|---|---|---|---|---|---|
| Binding Energy (kcal/mol) | Binding Energy (kcal/mol) | ||||||
| Ligand-1 | −14.30 | - | - | EEC | −13.13 | −10.14 | −9.27 |
| Ligand-2 | - | −14.54 | - | EED | −14.39 | −12.53 | −8.09 |
| Ligand-3 | - | - | −9.47 | EEE | −14.67 | −12.84 | −7.77 |
| CDD | −14.60 | −11.81 | −10.50 | EEG | −13.54 | −10.05 | −7.82 |
| DCD | −13.68 | −10.15 | −9.93 | EEH | −12.97 | −7.80 | −9.45 |
| DDD | −15.30 | −11.43 | −8.13 | EEV | −13.04 | −10.61 | −8.26 |
| DDE | −15.86 | −11.77 | −7.23 | EFD | −12.75 | −10.85 | −11.61 |
| DDG | −13.63 | −11.16 | −9.26 | EGE | −14.03 | −11.67 | −7.47 |
| DDQ | −13.31 | −11.52 | −6.67 | EMI | −11.62 | −9.64 | −8.11 |
| DDR | −12.74 | −10.25 | −11.60 | EVD | −14.25 | −11.80 | −8.83 |
| DDS | −14.35 | −12.20 | −10.54 | GDE | −15.25 | −12.09 | −7.79 |
| DEE | −13.21 | −10.56 | −6.35 | HDE | −12.24 | −11.46 | −9.48 |
| DGD | −14.34 | −12.07 | −9.37 | IDD | −13.81 | −12.83 | −8.86 |
| DGE | −12.72 | −11.47 | −7.13 | IMI | −7.88 | −7.57 | −8.09 |
| DIM | −10.90 | −9.63 | −9.12 | LYM | −8.09 | −8.43 | −9.84 |
| DMR | −12.48 | −11.43 | −9.76 | MDD | −11.99 | −10.67 | −10.36 |
| DNN | −13.33 | −11.49 | −9.91 | MEF | −10.29 | −9.66 | −9.23 |
| DTD | −14.34 | −12.42 | −8.84 | MEK | −11.28 | −10.59 | −12.31 |
| EAD | −13.78 | −11.17 | −9.32 | MWM | −8.68 | −8.95 | −9.44 |
| EDA | −12.78 | −10.60 | −9.20 | PEE | −11.16 | −10.09 | −6.71 |
| EDC | −14.01 | −8.53 | −9.29 | QEE | −13.54 | −11.57 | −8.12 |
| EDD | −15.52 | −13.60 | −8.62 | QQE | −13.05 | −11.09 | −9.47 |
| EDE | −13.95 | −12.26 | −6.95 | TNE | −12.95 | −11.56 | −9.33 |
| EDL | −12.29 | −11.09 | −9.00 | VRE | −11.61 | −11.35 | −10.79 |
| EDM | −12.23 | −11.25 | −9.78 | VYK | −7.71 | −9.92 | −11.63 |
| EDQ | −12.62 | −11.01 | −8.89 | WIY | −11.06 | −10.27 | −12.30 |
| EDT | −14.38 | −11.71 | −9.21 | WRM | −9.16 | −10.00 | −10.76 |
| EDV | −12.46 | −10.90 | −9.51 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ge, H.; Zhang, T.; Tang, Y.; Zhang, Y.; Yu, Y.; Men, F.; Liu, J.; Yu, Y. Computer-Aided Screening and Revealing Action Mechanism of Food-Derived Tripeptides Intervention in Acute Colitis. Int. J. Mol. Sci. 2022, 23, 13471. https://doi.org/10.3390/ijms232113471
Ge H, Zhang T, Tang Y, Zhang Y, Yu Y, Men F, Liu J, Yu Y. Computer-Aided Screening and Revealing Action Mechanism of Food-Derived Tripeptides Intervention in Acute Colitis. International Journal of Molecular Sciences. 2022; 23(21):13471. https://doi.org/10.3390/ijms232113471
Chicago/Turabian StyleGe, Huifang, Ting Zhang, Yuanhu Tang, Yan Zhang, Yue Yu, Fangbing Men, Jingbo Liu, and Yiding Yu. 2022. "Computer-Aided Screening and Revealing Action Mechanism of Food-Derived Tripeptides Intervention in Acute Colitis" International Journal of Molecular Sciences 23, no. 21: 13471. https://doi.org/10.3390/ijms232113471