

A Plasma Circular RNA Profile Differentiates Subjects with Alzheimer’s Disease and Mild Cognitive Impairment from Healthy Controls
 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Results
2.1. CircRNAs Profile by Microarray
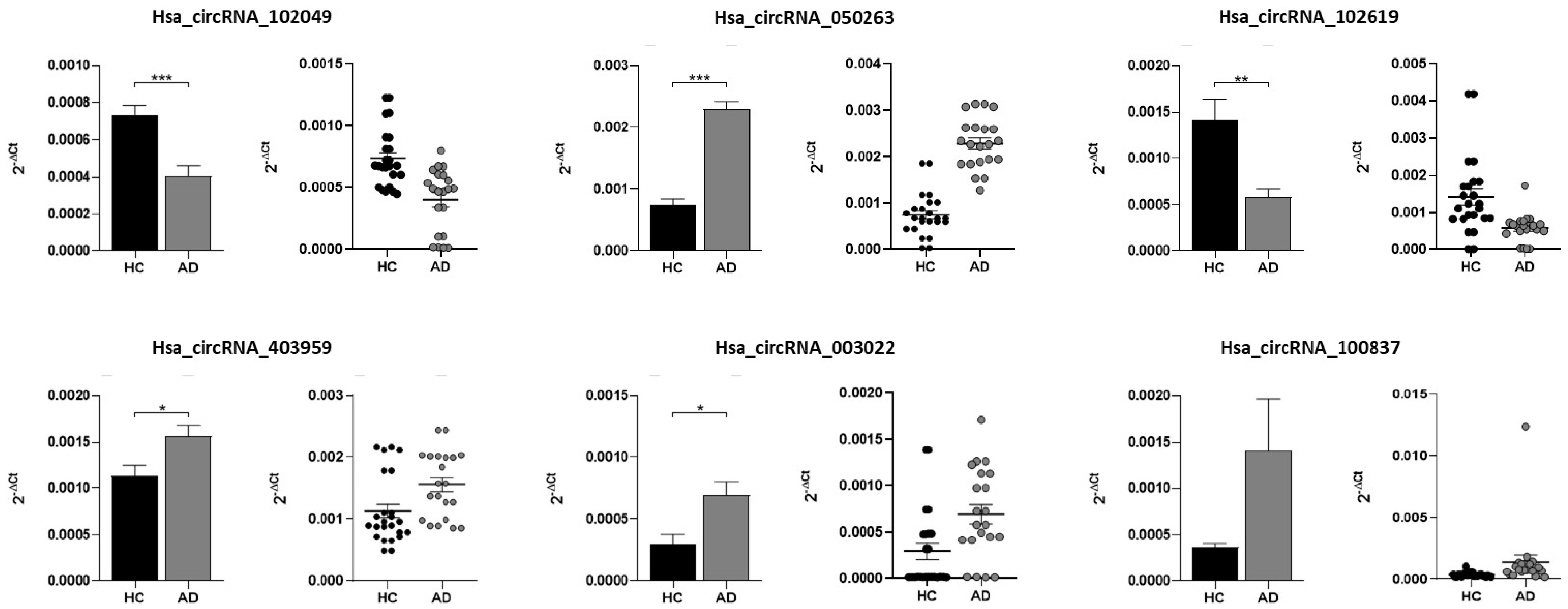
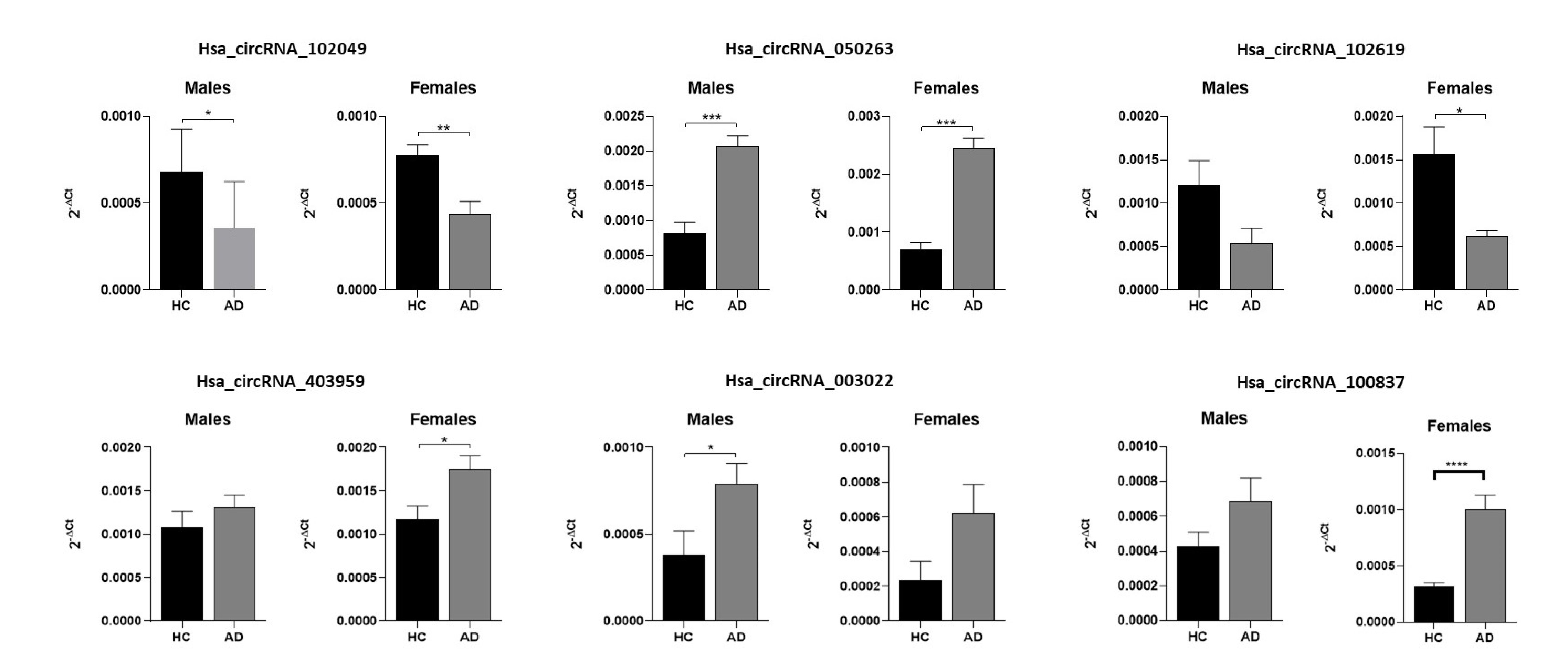
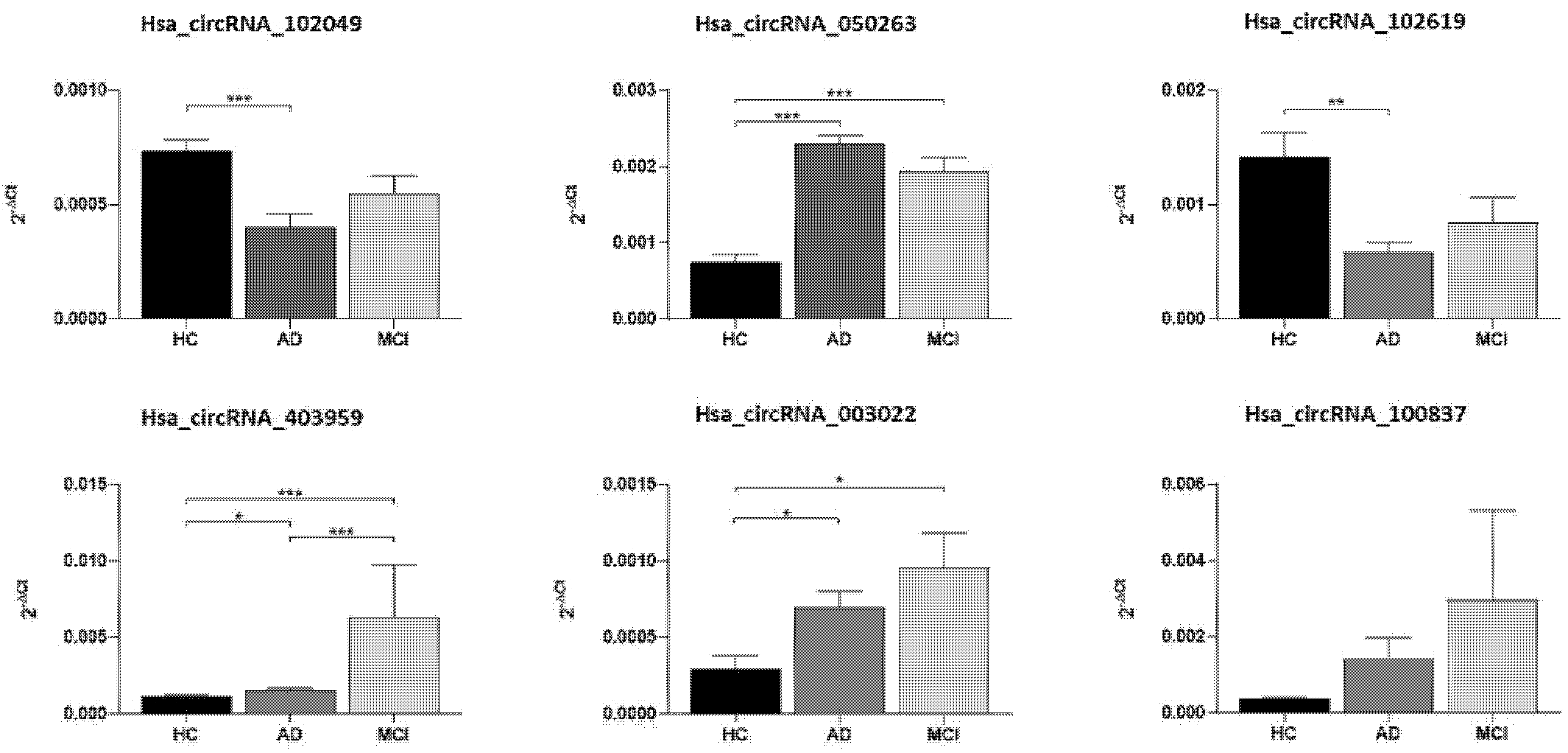
2.2. Validation of Candidate circRNAs by qRT-PCR
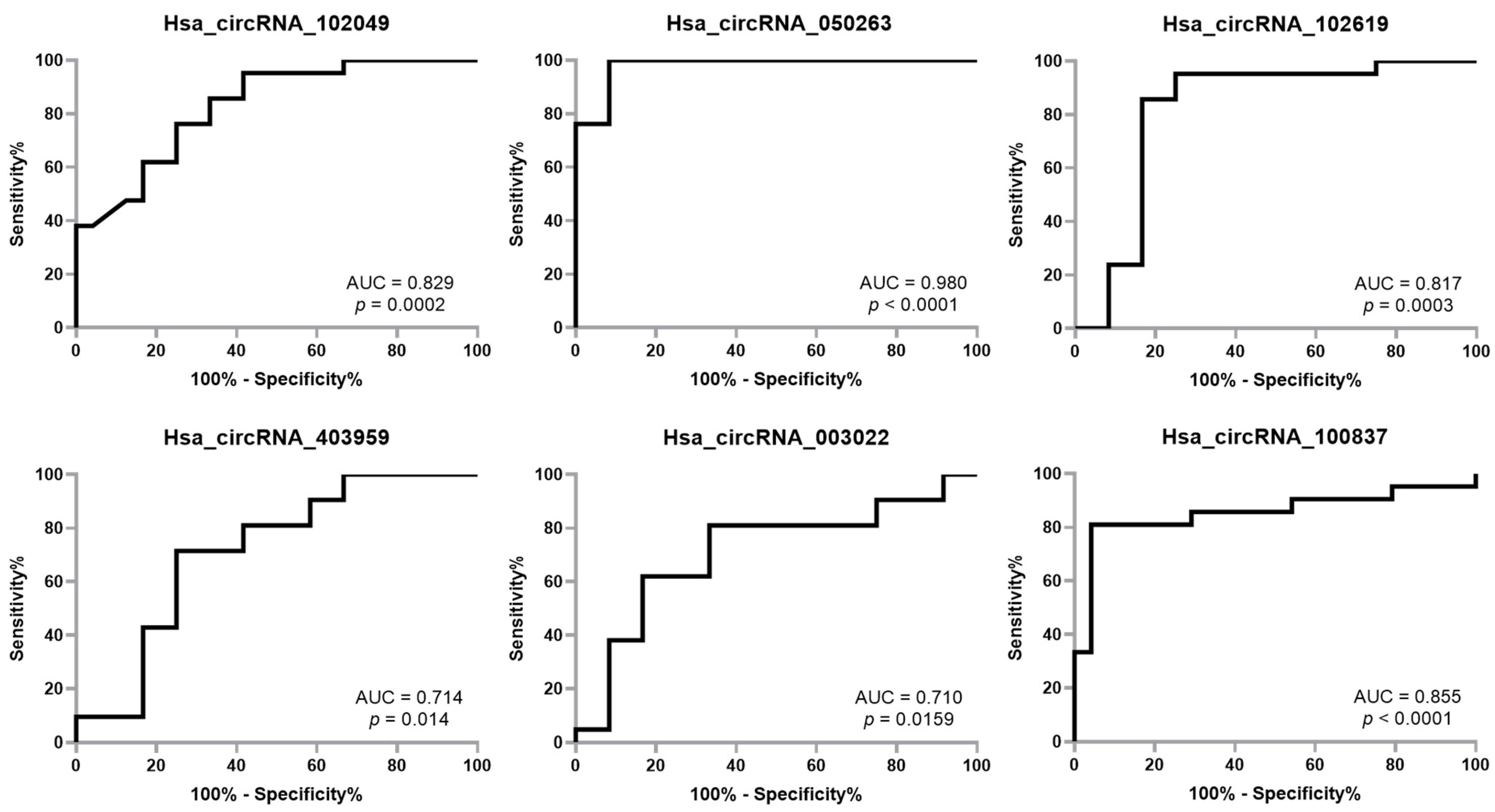
2.3. ROC Showed Good Diagnostic Accuracy for All Six miRNAs Analyzed
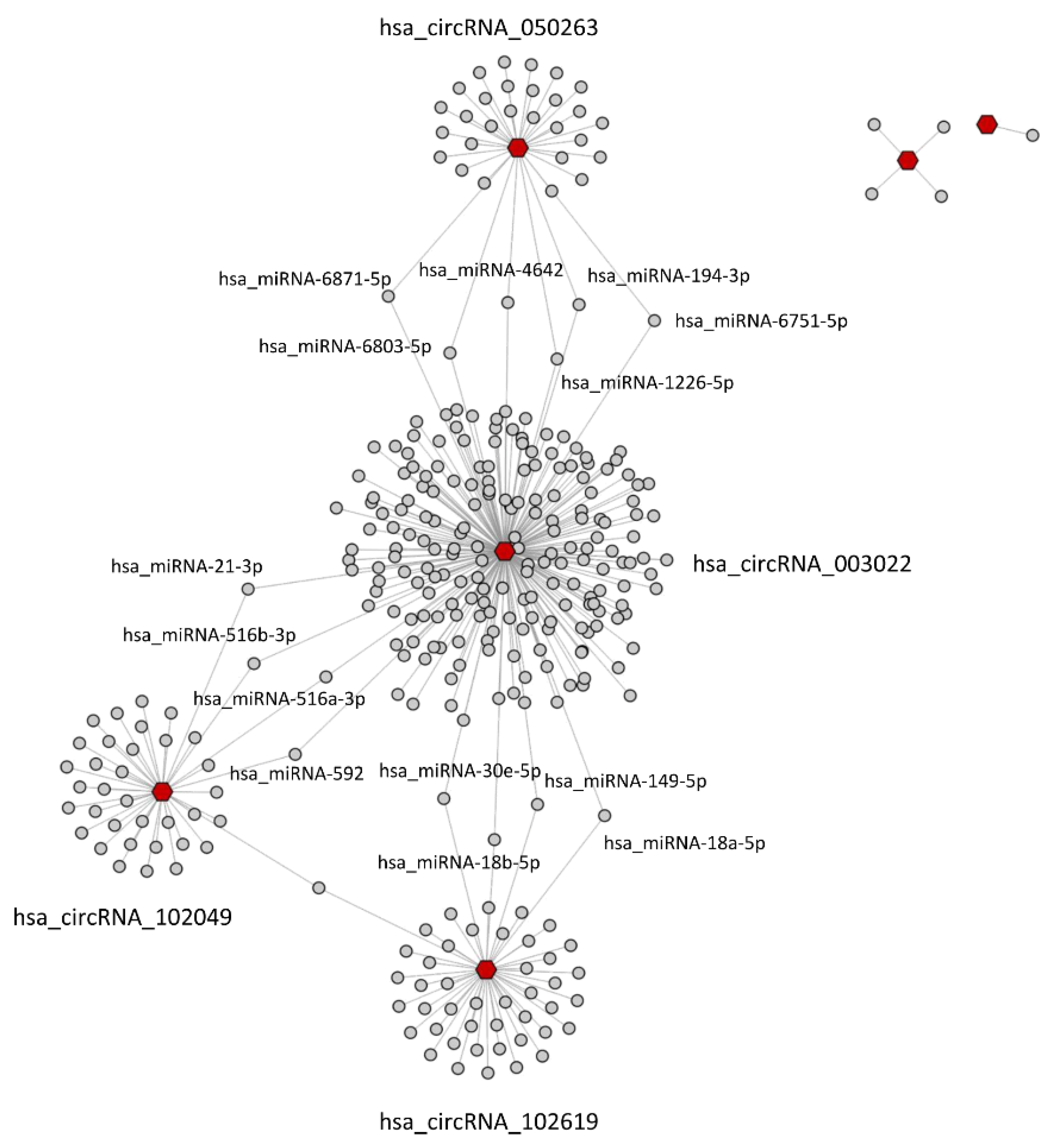
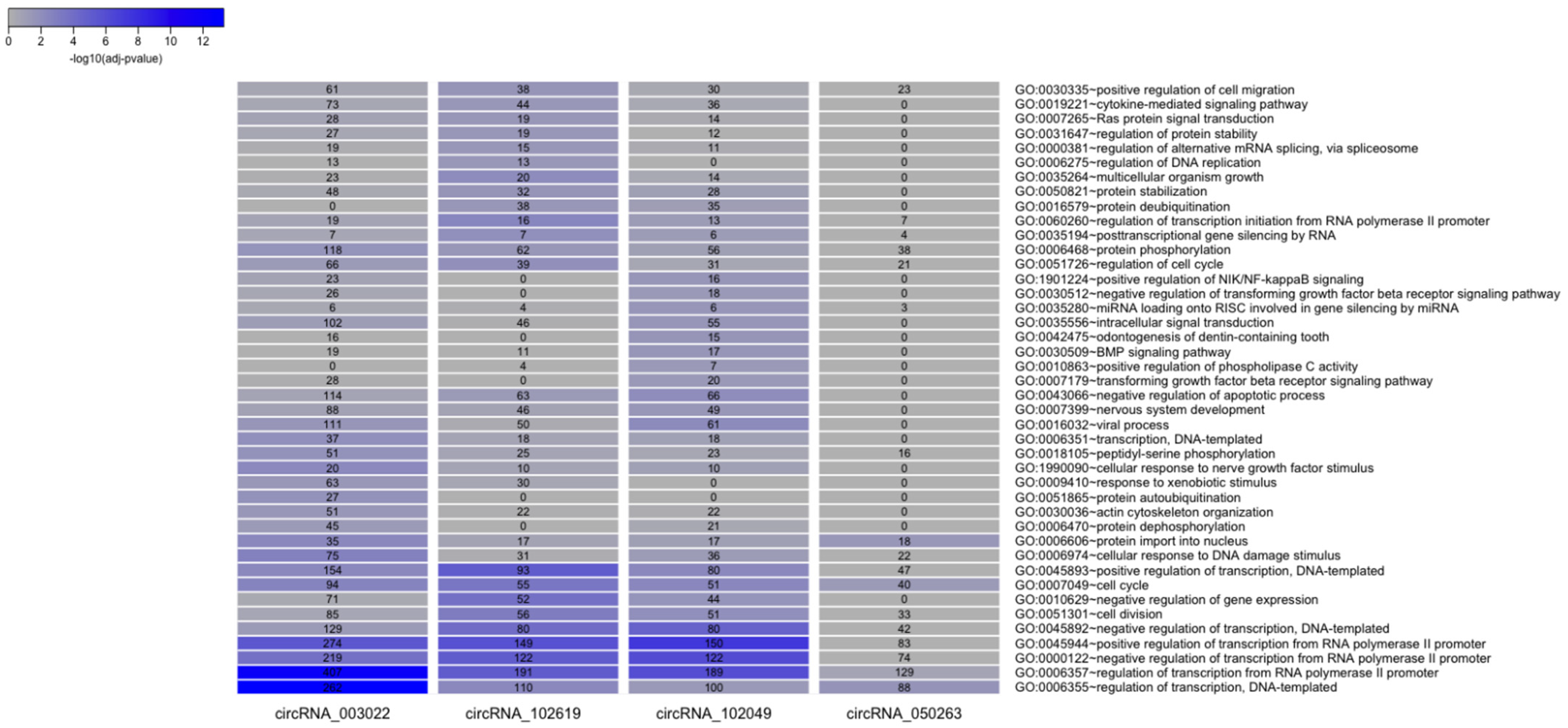
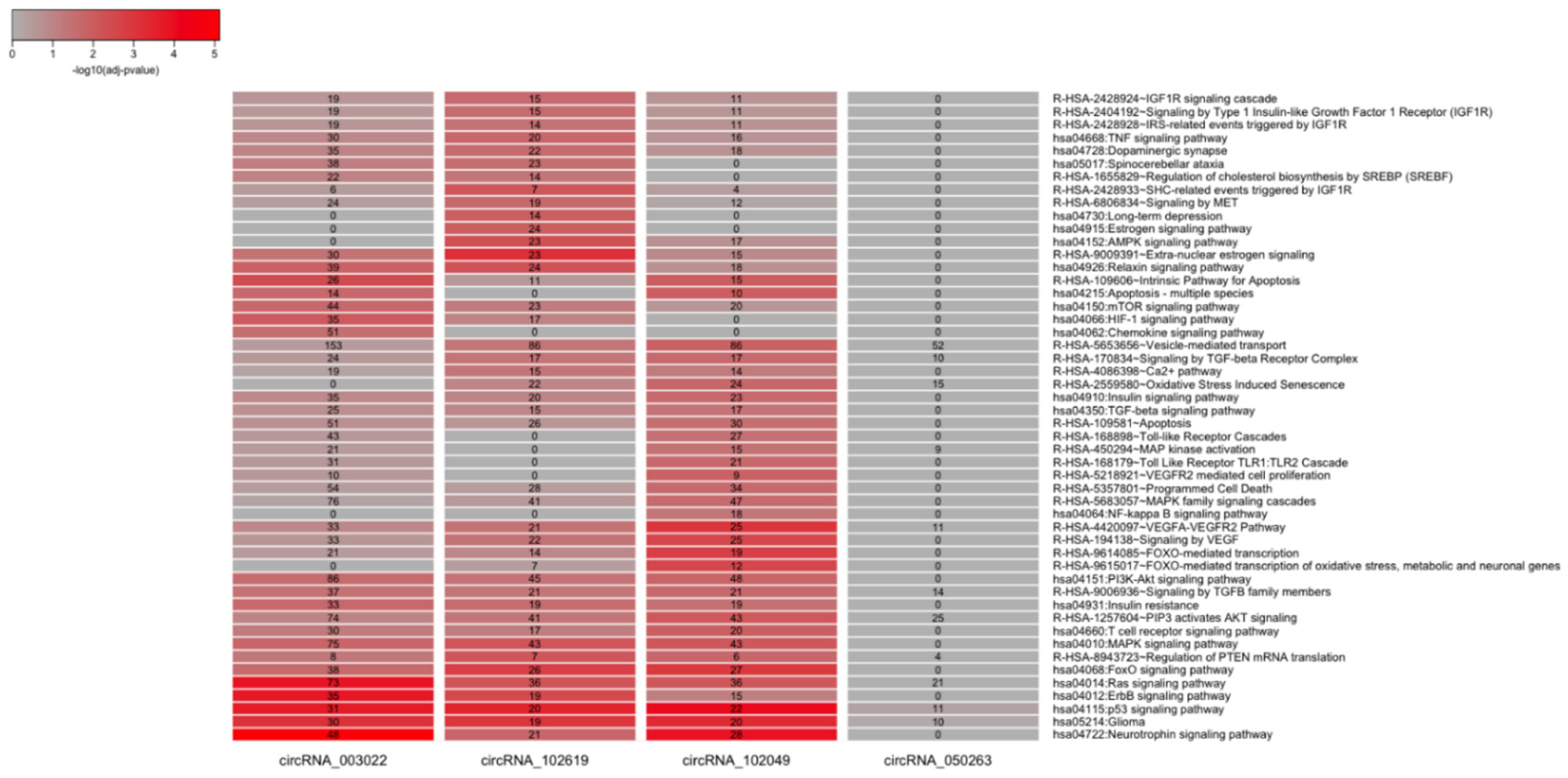
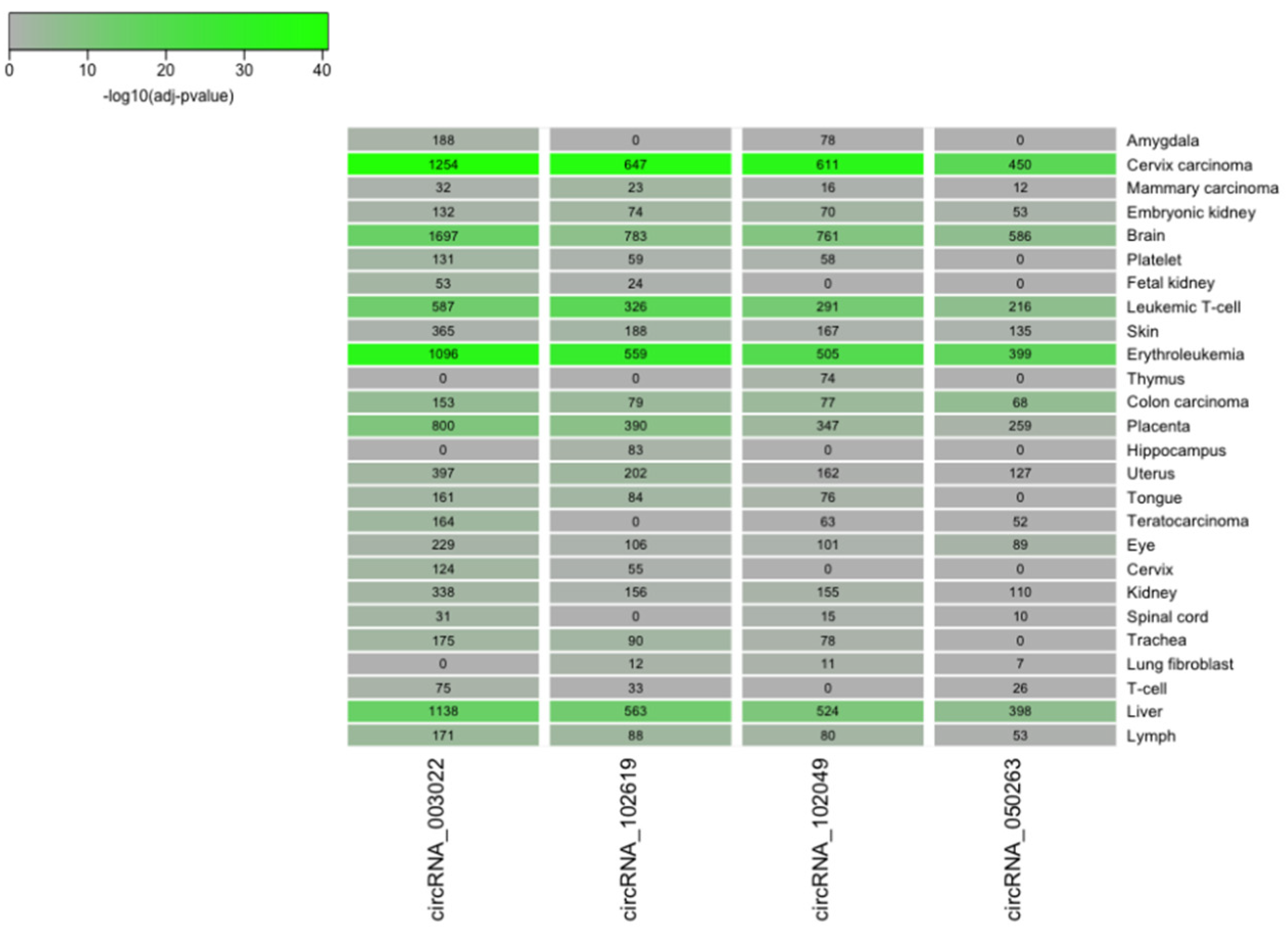
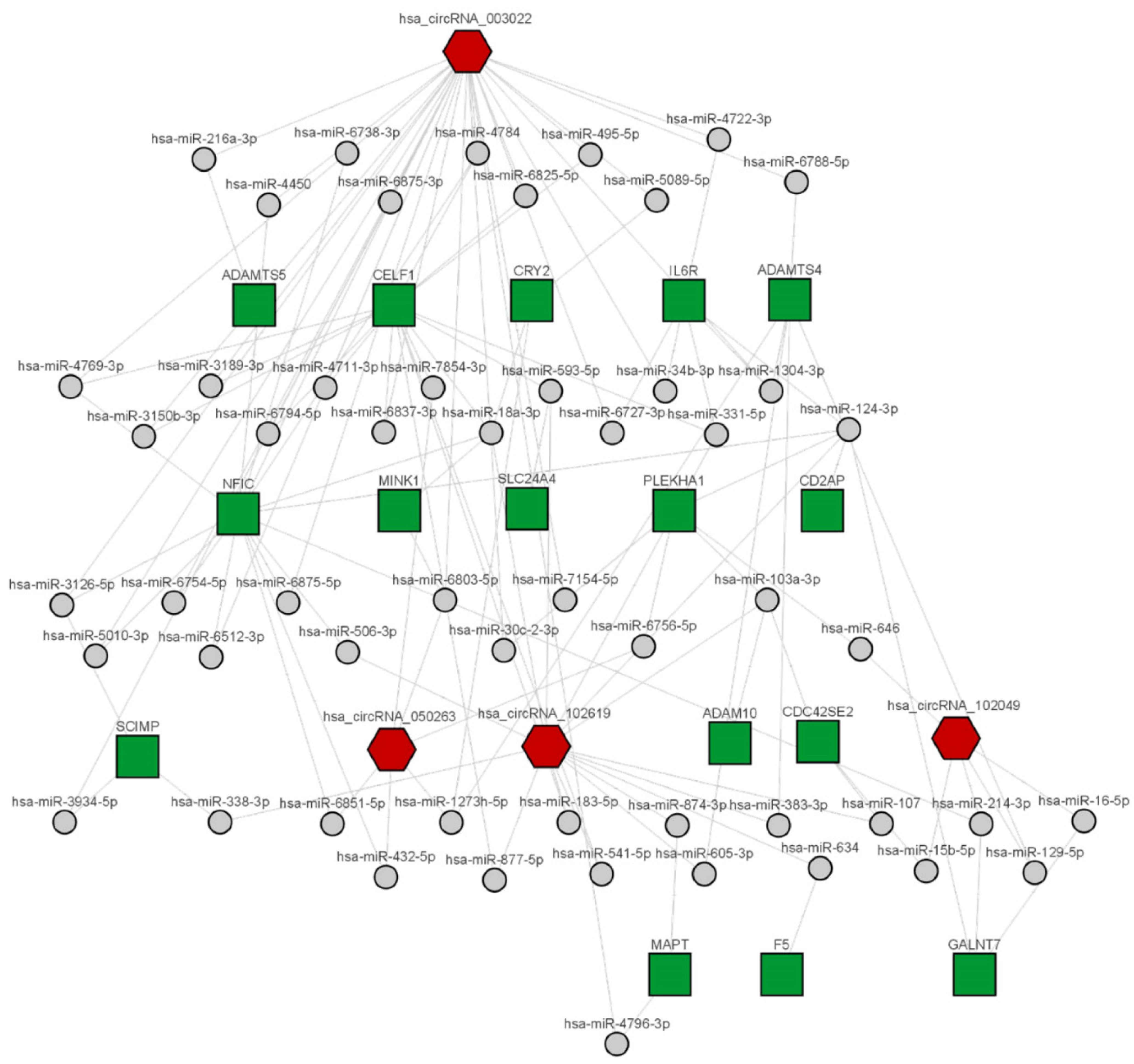
2.4. CircRNA/miRNA Network
3. Discussion
4. Materials and Methods
4.1. Recruited Population
4.2. Sample Preparation
4.3. SIMOA analysis
4.4. Microarray Analysis
4.5. Quantitative Real-Time PCR Analysis
4.6. CircRNA-microRNA-mRNA Network
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jack, C.R.; Petersen, R.C., Jr.; Xu, Y.C.; O’Brien, P.C.; Smith, G.E.; Ivnik, R.J.; Boeve, B.F.; Waring, S.C.; Tangalos, E.G.; Kokmen, E. Prediction of AD with MRI-based hippocampal volume in mild cognitive impairment. Neurology 1999, 52, 1397–1403. [Google Scholar] [CrossRef] [Green Version]
- Choo, I.H.; Ni, R.; Schöll, M.; Wall, A.; Almkvist, O.; Nordberg, A. Combination of 18F-FDG PET and cerebrospinal fluid biomarkers as a better predictor of the progression to Alzheimer’s disease in mild cognitive impairment patients. JAD 2013, 33, 929–939. [Google Scholar] [CrossRef]
- Grimmer, T.; Wutz, C.; Alexopoulos, P.; Drzezga, A.; Förster, S.; Förstl, H.; Goldhardt, O.; Ortner, M.; Sorg, C.; Kurz, A. Visual Versus Fully Automated Analyses of 18F-FDG and Amyloid PET for Prediction of Dementia Due to Alzheimer Disease in Mild Cognitive Impairment. J. Nucl. Med. 2016, 57, 204–207. [Google Scholar] [CrossRef] [Green Version]
- Jack, C.R.; Bennett, D.A., Jr.; Blennow, K.; Carrillo, M.C.; Dunn, B.; Haeberlein, S.B.; Holtzman, D.M.; Jagust, W.; Jessen, F.; Karlawish, J.; et al. NIA-AA Research Framework: Toward a biological definition of Alzheimer’s disease. Alzheimers Dement. 2018, 14, 535–562. [Google Scholar] [CrossRef]
- Hansson, O.; Zetterberg, H.; Buchhave, P.; Londos, E.; Blennow, K.; Minthon, L. Association between CSF biomarkers and incipient Alzheimer’s disease in patients with mild cognitive impairment: A follow-up study. Lancet Neurol. 2006, 5, 228–234. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Yang, T.; Xiao, J. Circular RNAs: Promising Biomarkers for Human Diseases. EBioMedicine 2018, 34, 267–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Yang, Y.; Li, Z.Q.; Li, L.C.; Zhu, X.H. Circular RNAs: From biogenesis and function to diseases. Chin. Med. J. 2019, 132, 2457–2464. [Google Scholar] [CrossRef]
- Huang, J.L.; Su, M.; Wu, D.P. Functional roles of circular RNAs in Alzheimer’s disease. Ageing Res. Rev. 2020, 60, 101058. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Yang, Z.; Jia, R.; Ge, S. The novel roles of circRNAs in human cancer. Mol. Cancer 2019, 18, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhter, R. Circular RNA and Alzheimer’s Disease. Adv. Exp. Med. Biol. 2018, 1087, 239–243. [Google Scholar]
- Li, Y.; Lv, Z.; Zhang, J.; Ma, Q.; Li, Q.; Song, L.; Gong, L.; Zhu, Y.; Li, X.; Hao, Y.; et al. Profiling of differentially expressed circular RNAs in peripheral blood mononuclear cells from Alzheimer’s disease patients. Metab. Brain Dis. 2020, 35, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Su, P.; Liu, Y.; Zhang, X.; Yan, J.; An, X.; Wang, X.; Gu, S. Abnormal expression of circRNA_089763 in the plasma exosomes of patients with post-operative cognitive dysfunction after coronary artery bypass grafting. Mol. Med. Rep. 2019, 20, 2549–2562. [Google Scholar] [CrossRef] [Green Version]
- Vogrinc, D.; Goričar, K.; Dolžan, V. Genetic Variability in Molecular Pathways Implicated in Alzheimer’s Disease: A Comprehensive Review. Front. Aging Neurosci. 2021, 13, 646901. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M.A.; Mohan, A.; Mather, K.A. Going around in circles: Deciphering the role of circular RNAs in neurodegenerative disease. Curr. Opin. Psychiatry 2020, 33, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Safari, M.; Taheri, M.; Samadian, M. Expression of Linear and Circular lncRNAs in Alzheimer’s Disease. J. Mol. Neurosci. 2022, 72, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Cochran, K.R.; Veeraraghavan, K.; Kundu, G.; Mazan-Mamczarz, K.; Coletta, C.; Thambisetty, M.; Gorospe, M.; De, S. Systematic Identification of circRNAs in Alzheimer’s Disease. Genes 2021, 12, 1258. [Google Scholar] [CrossRef]
- Ramirez, A.; Heimbach, A.; Gründemann, J.; Stiller, B.; Hampshire, D.; Cid, L.P.; Goebel, I.; Mubaidin, A.F.; Wriekat, A.-L.; Roeper, J.; et al. Hereditary parkinsonism with dementia is caused by mutations in ATP13A2, encoding a lysosomal type 5 P-type ATPase. Nat. Genet. 2006, 38, 1184–1191. [Google Scholar] [CrossRef]
- Sørensen, D.M.; Holemans, T.; van Veen, S.; Martin, S.; Arslan, T.; Haagendahl, I.W.; Holen, H.W.; Hamouda, N.N.; Eggermont, J.; Palmgren, M.; et al. Parkinson disease related ATP13A2 evolved early in animal evolution. PLoS ONE 2018, 13, e0193228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbe, C.; Amaral, T.; Peris, K.; Hauschild, A.; Arenberger, P.; Bastholt, L.; Bataille, V.; Del Marmol, V.; Dréno, B.; Fargnoli, M.C.; et al. European consensus-based interdisciplinary guideline for melanoma. Part 1: Diagnostics: Update 2022. Eur. J. Cancer 2020, 126, 141–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Liu, N.; Wei, W.; Zhang, Z.; Li, H. Integrated Analysis of Weighted Gene Coexpression Network Analysis Identifying Six Genes as Novel Biomarkers for Alzheimer’s Disease. Oxidative Med. Cell. Longev. 2021, 2021, 9918498. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, D.; Catania, A.; Viscomi, C.; Deleidi, M.; Bindoff, L.A.; Ghezzi, D.; Zeviani, M. Role of PITRM1 in Mitochondrial Dysfunction and Neurodegeneration. Biomedicines 2021, 9, 833. [Google Scholar] [CrossRef]
- Zhang, Y.; Yao, H.; Li, Y.; Yang, L.; Zhang, L.; Chen, J.; Wang, Y.; Li, X. Circular RNA TADA2A promotes proliferation and migration via modulating of miR-638/KIAA0101 signal in non-small cell lung cancer. Oncol. Rep. 2021, 46, 201. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, P.; Huang, J.; Liao, Y.; Pan, C.; Liu, J.; Du, Q.; Liu, T.; Shang, C.; Ooi, S.; et al. Circular RNA hsa_circ_0043280 inhibits cervical cancer tumor growth and metastasis via miR-203a-3p/PAQR3 axis. Cell Death Dis. 2021, 12, 888. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Kim, H.; Jo, A.; Khang, R.; Park, C.-H.; Park, S.-J.; Kwag, E.; Shin, J.-H. α-Synuclein A53T Binds to Transcriptional Adapter 2-Alpha and Blocks Histone H3 Acetylation. Int. J. Mol. Sci. 2021, 22, 5392. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.M.; Giuffre, E.; Moseley, P.; Hitchcock, P.F. The MicroRNA, miR-18a, Regulates NeuroD and Photoreceptor Differentiation in the Retina of Zebrafish. Dev. Neurobiol. 2019, 79, 202–219. [Google Scholar] [CrossRef] [Green Version]
- Magner, E.; Sandoval-Sanchez, P.; Kramer, A.C.; Thummel, R.; Hitchcock, P.F.; Taylor, S.M. Disruption of miR-18a Alters Proliferation, Photoreceptor Replacement Kinetics, Inflammatory Signaling, and Microglia/Macrophage Numbers During Retinal Regeneration in Zebrafish. Mol. Neurobiol. 2022, 59, 2910–2931. [Google Scholar] [CrossRef]
- Soleimani, T.; Falsafi, N.; Fallahi, H. Dissection of Regulatory Elements During Direct Conversion of Somatic Cells Into Neurons. J. Cell Biochem. 2017, 118, 3158–3170. [Google Scholar] [CrossRef]
- Nasirishargh, A.; Kumar, P.; Ramasubramanian, L.; Clark, K.; Hao, D.; Lazar, S.V.; Wang, A. Exosomal microRNAs from mesenchymal stem/stromal cells: Biology and applications in neuroprotection. World J. Stem Cells. 2021, 13, 776–794. [Google Scholar] [CrossRef]
- Shu, S.Y.; Qing, D.; Wang, B.; Zeng, Q.-Y.; Chen, Y.-C.; Jin, Y.; Zeng, C.-C.; Bao, R. Comparison of microRNA expression in hippocampus and the marginal division (MrD) of the neostriatum in rats. J. Biomed. Sci. 2013, 20, 9. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhou, Y.; Wu, Y.-J.; Ma, L.; Wang, R.-J.; Huang, S.-Q.; Gao, R.-R.; Liu, L.-H.; Shao, Z.-H.; Shi, H.-J.; et al. Novel cerebellum-enriched miR-592 may play a role in neural progenitor cell differentiation and neuronal maturation through regulating Lrrc4c and Nfasc in rat. Curr. Mol. Med. 2013, 13, 1432–1445. [Google Scholar] [CrossRef] [PubMed]
- Vreugdenhil, E.; Verissimo, C.S.L.; Mariman, R.; Kamphorst, J.T.; Barbosa, J.S.; Zweers, T.; Champagne, D.L.; Schouten, T.; Meijer, O.; De Kloet, E.R.; et al. MicroRNA 18 and 124a down-regulate the glucocorticoid receptor: Implications for glucocorticoid responsiveness in the brain. Endocrinology 2009, 150, 2220–2228. [Google Scholar] [CrossRef]
- Shimizu, S.; Tanaka, T.; Takeda, T.; Tohyama, M.; Miyata, S. The Kampo Medicine Yokukansan Decreases MicroRNA-18 Expression and Recovers Glucocorticoid Receptors Protein Expression in the Hypothalamus of Stressed Mice. Biomed. Res. Int. 2015, 2015, 797280. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.; Jin, H.-J.; Zhu, Y.-Y.; Fang, Z.; Mao, L.; He, Q.; Xia, Y.-P.; Li, M.; Li, Y.; Chen, X.; et al. MicroRNA-149-5p regulates blood-brain barrier permeability after transient middle cerebral artery occlusion in rats by targeting S1PR2 of pericytes. FASEB J. 2018, 32, 3133–3148. [Google Scholar] [CrossRef] [Green Version]
- Delalle, I. MicroRNAs as Candidates for Bipolar Disorder Biomarkers. Psychiatr. Danub. 2021, 33, 451–455. [Google Scholar]
- Choi, J.L.; Kao, P.F.; Itriago, E.; Zhan, Y.; Kozubek, J.A.; Hoss, A.G.; Banigan, M.G.; Vanderburg, C.R.; Rezvani, A.H.; Latourelle, J.C.; et al. miR-149 and miR-29c as candidates for bipolar disorder biomarkers. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2017, 174, 315–323. [Google Scholar] [CrossRef]
- Khandelwal, N.; Dey, S.K.; Chakravarty, S.; Kumar, A. miR-30 Family miRNAs Mediate the Effect of Chronic Social Defeat Stress on Hippocampal Neurogenesis in Mouse Depression Model. Front. Mol. Neurosci. 2019, 12, 188. [Google Scholar] [CrossRef] [Green Version]
- Amar, L.; Benoit, C.; Beaumont, G.; Vacher, C.M.; Crepin, D.; Taouis, M.; Baroin-Tourancheau, A. MicroRNA expression profiling of hypothalamic arcuate and paraventricular nuclei from single rats using Illumina sequencing technology. J. Neurosci. Methods. 2012, 209, 134–143. [Google Scholar] [CrossRef]
- Bai, X.; Bian, Z. MicroRNA-21 Is a Versatile Regulator and Potential Treatment Target in Central Nervous System Disorders. Front. Mol. Neurosci. 2022, 15, 842288. [Google Scholar] [CrossRef]
- Wu, G.D.; Li, Z.H.; Li, X.; Zheng, T.; Zhang, D.K. microRNA-592 blockade inhibits oxidative stress injury in Alzheimer’s disease astrocytes via the KIAA0319-mediated Keap1/Nrf2/ARE signaling pathway. Exp. Neurol. 2020, 324, 113128. [Google Scholar] [CrossRef]
- Barh, D.; García-Solano, M.E.; Tiwari, S.; Bhattacharya, A.; Jain, N.; Torres-Moreno, D.; Ferri, B.; Silva, A.; Azevedo, V.; Ghosh, P.; et al. BARHL1 Is Downregulated in Alzheimer’s Disease and May Regulate Cognitive Functions through ESR1 and Multiple Pathways. Genes 2017, 8, 245. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.R.; Cui, X.L.; Zheng, Y.; Zhang, G.R.; Li, P.; Huang, H.; Zhao, Y.Y.; Bo, X.C.; Wang, S.Q.; Zhou, W.X.; et al. Nodes and biological processes identified on the basis of network analysis in the brain of the senescence accelerated mice as an Alzheimer’s disease animal model. Front. Aging Neurosci. 2013, 5, 65. [Google Scholar] [CrossRef] [Green Version]
- Du, W.; Lei, C.; Dong, Y. MicroRNA-149 is downregulated in Alzheimer’s disease and inhibits β-amyloid accumulation and ameliorates neuronal viability through targeting BACE1. Genet. Mol. Biol. 2021, 44, e20200064. [Google Scholar] [CrossRef]
- Ghasemloo, E.; Mostafavi, H.; Hosseini, M.; Forouzandeh, M.; Eskandari, M.; Mousavi, S.S. Neuroprotective effects of coenzyme Q10 in Parkinson’s model via a novel Q10/miR-149-5p/MMPs pathway. Metab. Brain Dis. 2021, 36, 2089–2100. [Google Scholar] [CrossRef]
- Lv, Z.; Hu, L.; Yang, Y.; Zhang, K.; Sun, Z.; Zhang, J.; Zhang, L.; Hao, Y. Comparative study of microRNA profiling in one Chinese Family with PSEN1 G378E mutation. Metab. Brain Dis. 2018, 33, 1711–1720. [Google Scholar] [CrossRef]
- Dube, U.; Del-Aguila, J.L.; Li, Z.; Budde, J.P.; Jiang, S.; Hsu, S.; Ibanez, L.; Fernandez, M.V.; Farias, F.; Norton, J.; et al. An atlas of cortical circular RNA expression in Alzheimer disease brains demonstrates clinical and pathological associations. Nat. Neurosci. 2019, 22, 1903–1912. [Google Scholar] [CrossRef]
- Verduci, L.; Tarcitano, E.; Strano, S.; Yarden, Y.; Blandino, G. CircRNAs: Role in human diseases and potential use as biomarkers. Cell Death Dis. 2021, 12, 468. [Google Scholar] [CrossRef]
- Pietraforte, D.; Straface, E.; Piscopo, P.; Vona, R.; Confaloni, A. Sex-related biomarkers in cardiovascular and neurodegenerative disorders. Ann. Ist. Super Sanita. 2016, 52, 230–239. [Google Scholar]
- Piscopo, P.; Rivabene, R.; Galimberti, D.; Crestini, A.; Talarico, G.; Vanacore, N.; Scarpini, E.; Bruno, G.; Confaloni, A. Gender effects on plasma PGRN levels in patients with Alzheimer’s disease: A preliminary study. J. Alzheimers Dis. 2013, 35, 313–318. [Google Scholar] [CrossRef]
- Piscopo, P.; Bellenghi, M.; Manzini, V.; Crestini, A.; Pontecorvi, G.; Corbo, M.; Ortona, E.; Carè, A.; Confaloni, A. A Sex Perspective in Neurodegenerative Diseases: MicroRNAs as Possible Peripheral Biomarkers. Int. J. Mol. Sci. 2021, 22, 4423. [Google Scholar] [CrossRef]
- McKhann, G.; Drachman, D.; Folstein, M.; Katzman, R.; Price, D.; Stadlan, E.M. Clinical diagnosis of Alzheimer’s disease: Report of the NINCDS-ADRDA Work Group under the auspices of Department of Health and Human Services Task Force on Alzheimer’s Disease. Neurology 1984, 34, 939–944. [Google Scholar] [CrossRef] [Green Version]
- Petersen, R.C.; Doody, R.; Kurz, A.; Mohs, R.C.; Morris, J.C.; Rabins, P.V.; Ritchie, K.; Rossor, M.; Thal, L.; Winblad, B. Current concepts in mild cognitive impairment. Arch. Neurol. 2001, 58, 1985–1992. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Enright, A.J.; John, B.; Gaul, U.; Tuschl, T.; Sander, C.; Marks, D.S. MicroRNA targets in Drosophila. Genome Biol. 2003, 5, R1. [Google Scholar] [CrossRef]
- Huang, H.Y.; Lin, Y.C.; Li, J.; Huang, K.Y.; Shrestha, S.; Hong, H.C.; Tang, Y.; Chen, Y.G.; Jin, C.N.; Yu, Y.; et al. miRTarBase 2020: Updates to the experimentally validated microRNA-target interaction database. Nucleic Acids Res. 2020, 48, D148–D154. [Google Scholar] [CrossRef] [Green Version]
- Sticht, C.; De La Torre, C.; Parveen, A.; Gretz, N. An online resource for prediction of microRNA binding sites. PLoS ONE 2018, 13, e0206239. [Google Scholar] [CrossRef]
- Huang, D.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Fabregat, A.; Jupe, S.; Matthews, L.; Sidiropoulos, K.; Gillespie, M.; Garapati, P.; Haw, R.; Jassal, B.; Korninger, F.; May, B.; et al. The Reactome Pathway Knowledgebase. Nucleic Acids Res. 2018, 46, D649–D655. [Google Scholar] [CrossRef]
- The Uniprot Consortium. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D1. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pilot Study | Validation Study | ||||||
|---|---|---|---|---|---|---|---|
| HC | AD | HC | AD | MCI | |||
| N. | 5 | 5 | 24 | 21 | 5 | ||
| Sex (M/F) | 2/3 | 2/3 | p = n.s. | 10/14 | 9/12 | 3/2 | p = n.s. |
| Age at the evaluation | 73.8 ± 4.3 | 74.4 ± 2.3 | p = n.s. | 69.9 ± 6.6 | 74.4 ± 6.1 | 70.0 ± 7.3 | p = n.s. |
| Age at onset | - | 66.40 ± 6.5 | - | 70.6 ± 6.8 | 66.4 ± 6.5 | ||
| MMSE | - | 21.4 ± 1.4 | - | 21.1 ± 1.7 | 25.6 ± 0.9 | ||
| Aβ42/Aβ40 | NA | 0.04 ± 0.007 | 0.05 ± 0.001 | ||||
| Total-tau | NA | 2.31± 1.9 | 2.98 ± 0.6 | ||||
| p-tau | NA | 2.78 ± 0.5 | 2.37 ± 0.6 | ||||
| CircRNA | p-Value | FC (abs) | Regulation | Chrom. | Strand | circRNA_Type | Gene Symbol |
|---|---|---|---|---|---|---|---|
| hsa_circRNA_100837 | 0.0037 | 2.40 | up | chr11 | − | exonic | STX5 |
| hsa_circRNA_100760 | 0.0042 | 2.23 | up | chr11 | − | exonic | DENND5A |
| hsa_circRNA_403959 | 0.0051 | 2.23 | up | chr7 | − | exonic | BRAF |
| hsa_circRNA_001131 | 0.0122 | 2.55 | up | chr2 | − | intronic | TLR5 |
| hsa_circRNA_405788 | 0.0104 | 2.38 | up | chr19 | − | Exonic | CADM4 |
| hsa_circRNA_050263 | 0.0155 | 2.19 | up | chr19 | − | exonic | ATP13A1 |
| hsa_circRNA_003022 | 0.0180 | 2.14 | up | chr10 | − | exonic | PITRM1 |
| hsa_circRNA_407191 | 0.0126 | 2.00 | up | chr9 | + | Sense overlapping | AL161626.1 |
| hsa_circRNA_102750 | 0.0425 | 1.55 | up | chr2 | + | exonic | MEIS1 |
| hsa_circRNA_105042 | 0.0128 | 1.90 | up | chrX | − | exonic | GAB3 |
| hsa_circRNA_090183 | 0.0114 | 1.74 | up | chrX | + | exonic | PRRG1 |
| hsa_circRNA_401844 | 0.0111 | 1.92 | up | chr17 | − | exonic | TUBD1 |
| hsa_circRNA_080099 | 0.0073 | 1.66 | up | chr7 | − | exonic | MYO1G |
| hsa_circRNA_004907 | 0.0316 | 1.60 | up | chr10 | + | exonic | ZEB1 |
| hsa_circRNA_101222 | 0.0343 | 1.57 | up | chr13 | − | exonic | TPTE2 |
| hsa_circRNA_002165 | 0.0340 | 1.60 | up | chr6 | − | exonic | SRPK1 |
| hsa_circRNA_003022 | 0.0380 | 2.14 | up | chr10 | − | exonic | PITRM1 |
| hsa_circRNA_003574 | 0.0104 | 1.88 | up | chr20 | + | exonic | GID8 |
| hsa_circRNA_102885 | 0.0273 | 1.77 | up | chr2 | − | exonic | SATB2 |
| hsa_circRNA_104671 | 0.0283 | 1.82 | up | chr8 | − | exonic | UBR5 |
| hsa_circRNA_103618 | 0.0255 | 1.78 | up | chr4 | − | exonic | ARAP2 |
| hsa_circRNA_104220 | 0.0109 | 1.67 | up | chr6 | + | exonic | PCMT1 |
| hsa_circRNA_101752 | 0.0181 | 1.56 | up | chr16 | − | exonic | LOC100271836 |
| hsa_circRNA_100759 | 0.0206 | 1.72 | up | chr11 | − | exonic | DENND5A |
| hsa_circRNA_102049 | 0.0279 | 2.11 | down | chr17 | + | exonic | TADA2A |
| hsa_circRNA_102619 | 0.0036 | 2.00 | down | chr2 | − | exonic | NOL10 |
| hsa_circRNA_102645 | 0.0454 | 1.62 | down | chr2 | − | exonic | HADHA |
| circRNA | Mean Value (∆Ct) | Standard Error | p_Values | p Values Adjusted | ||
|---|---|---|---|---|---|---|
| Hsa_circRNA_102049 | HC | 0.000737 | 0.000048 | HC/AD | <0.001 | <0.001 |
| AD | 0.000404 | 0.000056 | HC/MCI | 0.355 | 0.474 | |
| MCI | 0.000549 | 0.000077 | AD/MCI | 0.687 | 0.687 | |
| Hsa_circRNA_050263 | HC | 0.000750 | 0.000093 | HC/AD | <0.001 | <0.001 |
| AD | 0.002293 | 0.000120 | HC/MCI | <0.001 | <0.001 | |
| MCI | 0.001945 | 0.000179 | AD/MCI | 0.494 | 0.668 | |
| Hsa_circRNA_102619 | HC | 0.001418 | 0.000215 | HC/AD | 0.003 | 0.006 |
| AD | 0.000585 | 0.000082 | HC/MCI | 0.430 | 0.548 | |
| MCI | 0.000839 | 0.000228 | AD/MCI | 0.217 | 1.000 | |
| Hsa_circRNA_403959 | HC | 0.001136 | 0.000114 | HC/AD | 0.013 | 0.026 |
| AD | 0.001559 | 0.000116 | HC/MCI | <0.001 | <0.001 | |
| MCI | 0.006260 | 0.003491 | AD/MCI | 0.001 | 0.001 | |
| Hsa_circRNA_003022 | HC | 0.000295 | 0.000085 | HC/AD | 0.015 | 0.094 |
| AD | 0.000695 | 0.000106 | HC/MCI | 0.015 | 0.016 | |
| MCI | 0.000954 | 0.000227 | AD/MCI | 0.777 | 0.451 | |
| Hsa_circRNA_100837 | HC | 0.000364 | 0.000039 | HC/AD | 0.389 | 0.072 |
| AD | 0.001407 | 0.000555 | HC/MCI | 0.070 | 0.055 | |
| MCI | 0.002972 | 0.002349 | AD/MCI | 0.513 | 1.000 | |
| Gene Name | Sequence (5′ to 3′) | Tm (°C) | Length of Product (bp) |
|---|---|---|---|
| hsa_circRNA_100837 | Forward:5′ AAGCAGTGGAAATTGAAGAGC3′ Reverse:5′ GCTGGCTTATTTGTCTGGATT3′ | 60 | 67 |
| hsa_circRNA_403959 | Forward:5′ AGAAGACAGGAATCGAATGGACT3′ Reverse:5′ CAGGTAATGAGGCAGGGGG3′ | 60 | 96 |
| hsa_circRNA_102619 | Forward:5′ GGGCATCTATTACATTCCATTCT3′ Reverse:5′ ATTATTCTCCGCAGCATCAGT3′ | 60 | 95 |
| hsa_circRNA_003022 | Forward:5′ GATGAAGGGAGCGTTTACAGA3′ Reverse:5′ GGGAACAGATGTCACCTAGCA3′ | 60 | 192 |
| hsa_circRNA_050263 | Forward:5′ CAAGCTCTCATCCATCCAGTG3′ Reverse:5′ ATGGGCGTACTCTCGTCCTC3′ | 60 | 73 |
| hsa_circRNA_102049 | Forward:5′ CACAGCCATTCCATTTCACTACT3′ Reverse:5′ CAAAGCCACAGTCCATCACAG3′ | 60 | 105 |
| β-actin | Forward:5′ GTGGCCGAGGACTTTGATTG3′ Reverse:5′ CCTGTAACAACGCATCTCATATT3′ | 60 | 73 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piscopo, P.; Manzini, V.; Rivabene, R.; Crestini, A.; Le Pera, L.; Pizzi, E.; Veroni, C.; Talarico, G.; Peconi, M.; Castellano, A.E.; et al. A Plasma Circular RNA Profile Differentiates Subjects with Alzheimer’s Disease and Mild Cognitive Impairment from Healthy Controls. Int. J. Mol. Sci. 2022, 23, 13232. https://doi.org/10.3390/ijms232113232
Piscopo P, Manzini V, Rivabene R, Crestini A, Le Pera L, Pizzi E, Veroni C, Talarico G, Peconi M, Castellano AE, et al. A Plasma Circular RNA Profile Differentiates Subjects with Alzheimer’s Disease and Mild Cognitive Impairment from Healthy Controls. International Journal of Molecular Sciences. 2022; 23(21):13232. https://doi.org/10.3390/ijms232113232
Chicago/Turabian StylePiscopo, Paola, Valeria Manzini, Roberto Rivabene, Alessio Crestini, Loredana Le Pera, Elisabetta Pizzi, Caterina Veroni, Giuseppina Talarico, Martina Peconi, Anna Elisa Castellano, and et al. 2022. "A Plasma Circular RNA Profile Differentiates Subjects with Alzheimer’s Disease and Mild Cognitive Impairment from Healthy Controls" International Journal of Molecular Sciences 23, no. 21: 13232. https://doi.org/10.3390/ijms232113232