
C-Series Coelenterazine-Driven Bioluminescence Signature Imaging
 , , and
, , and
Abstract
:1. Introduction
2. Experimental Method
2.1. Reagents and Instrumentation for the Synthesis of the C-Series CTZ Analogues
2.2. Preparation of Mammalian Cell Expression Vectors Encoding Each Marine Luciferase or Single-Chain Probe
2.3. Synthesis of C-Series CTZ Analogues
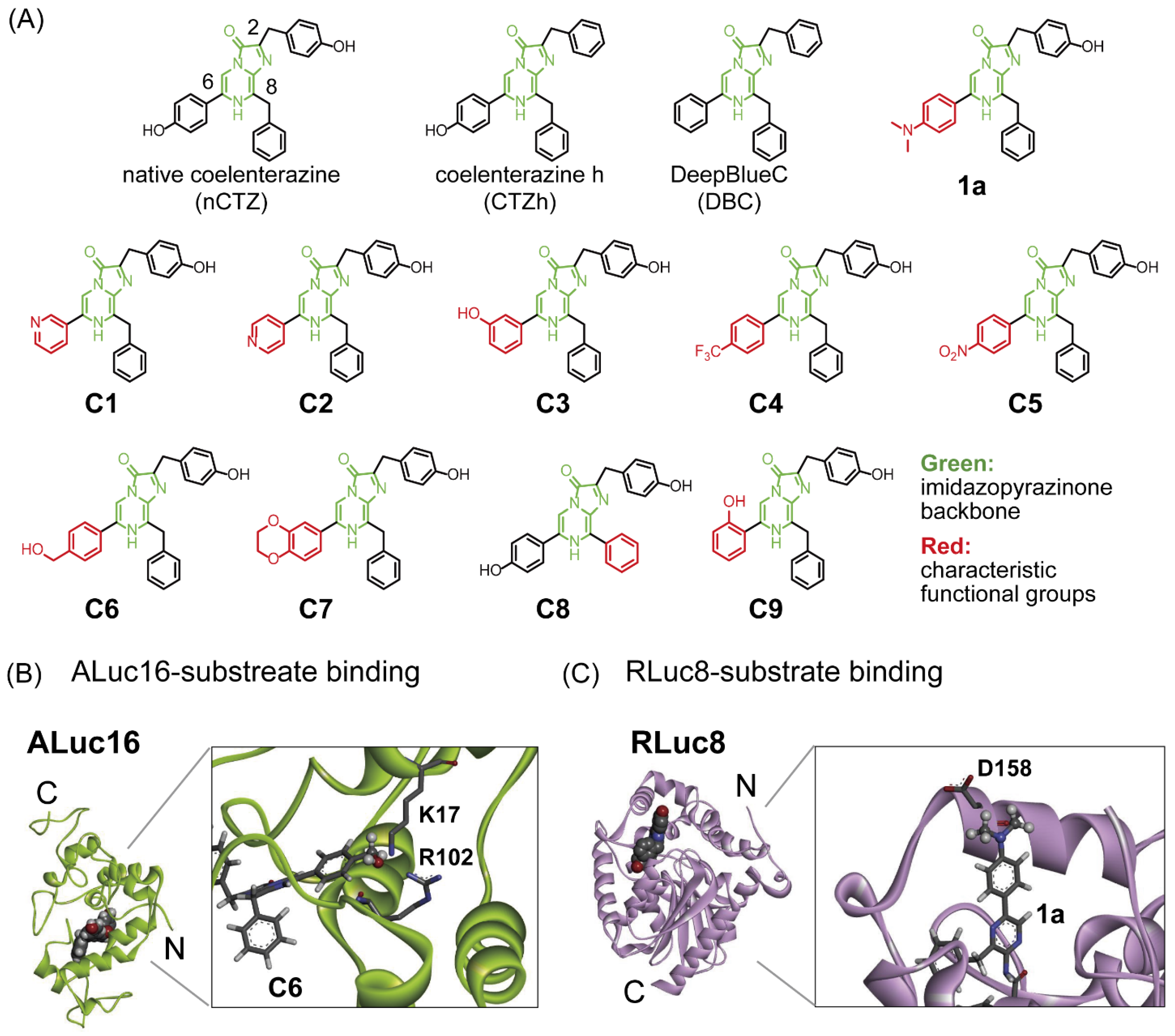
2.4. Modeling of the Binding Modes of Substrates in ALuc16 and RLuc8
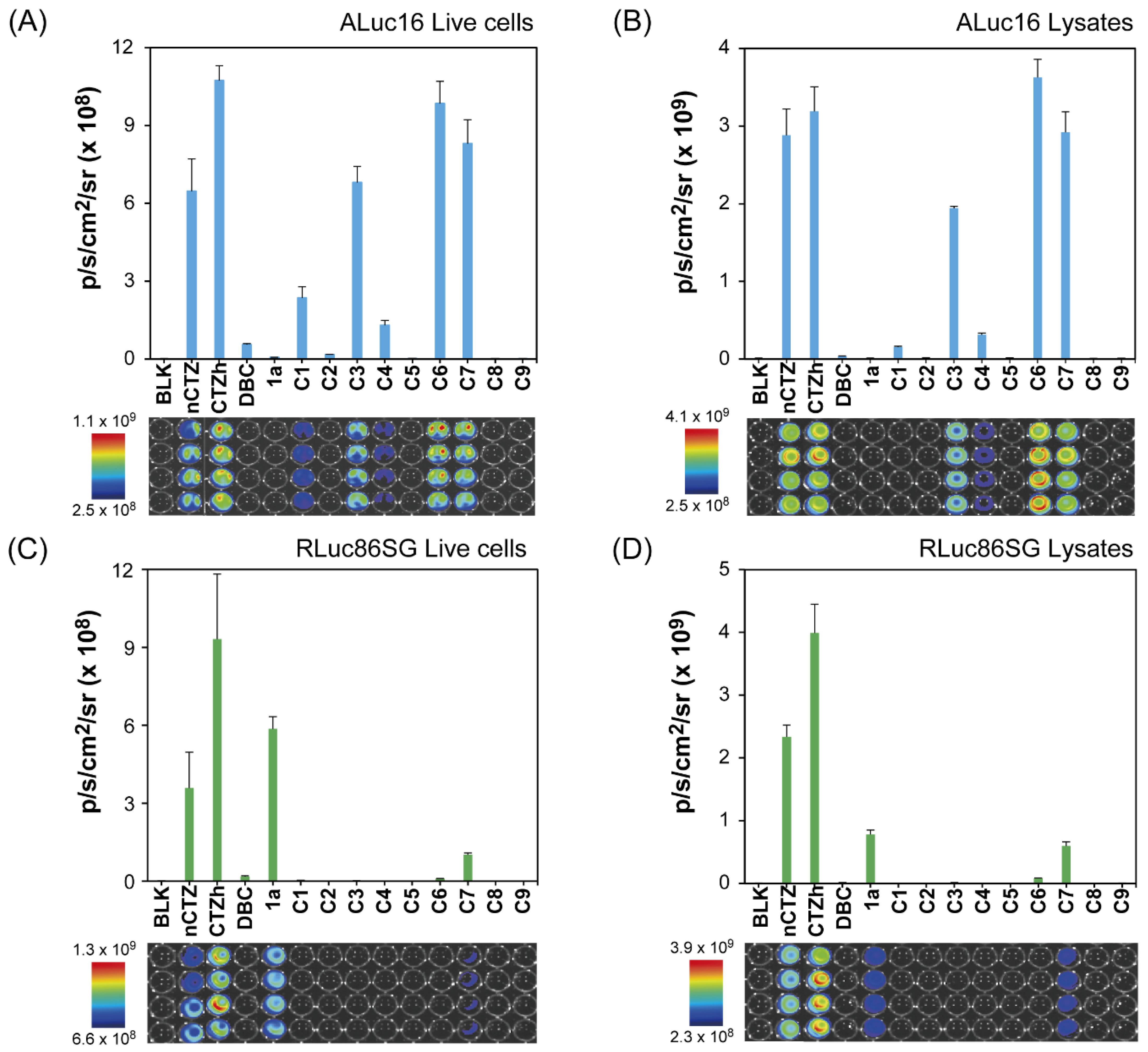
2.5. Absolute BL Intensities of the CTZ Analogues C1–C9 According to Marine Luciferases
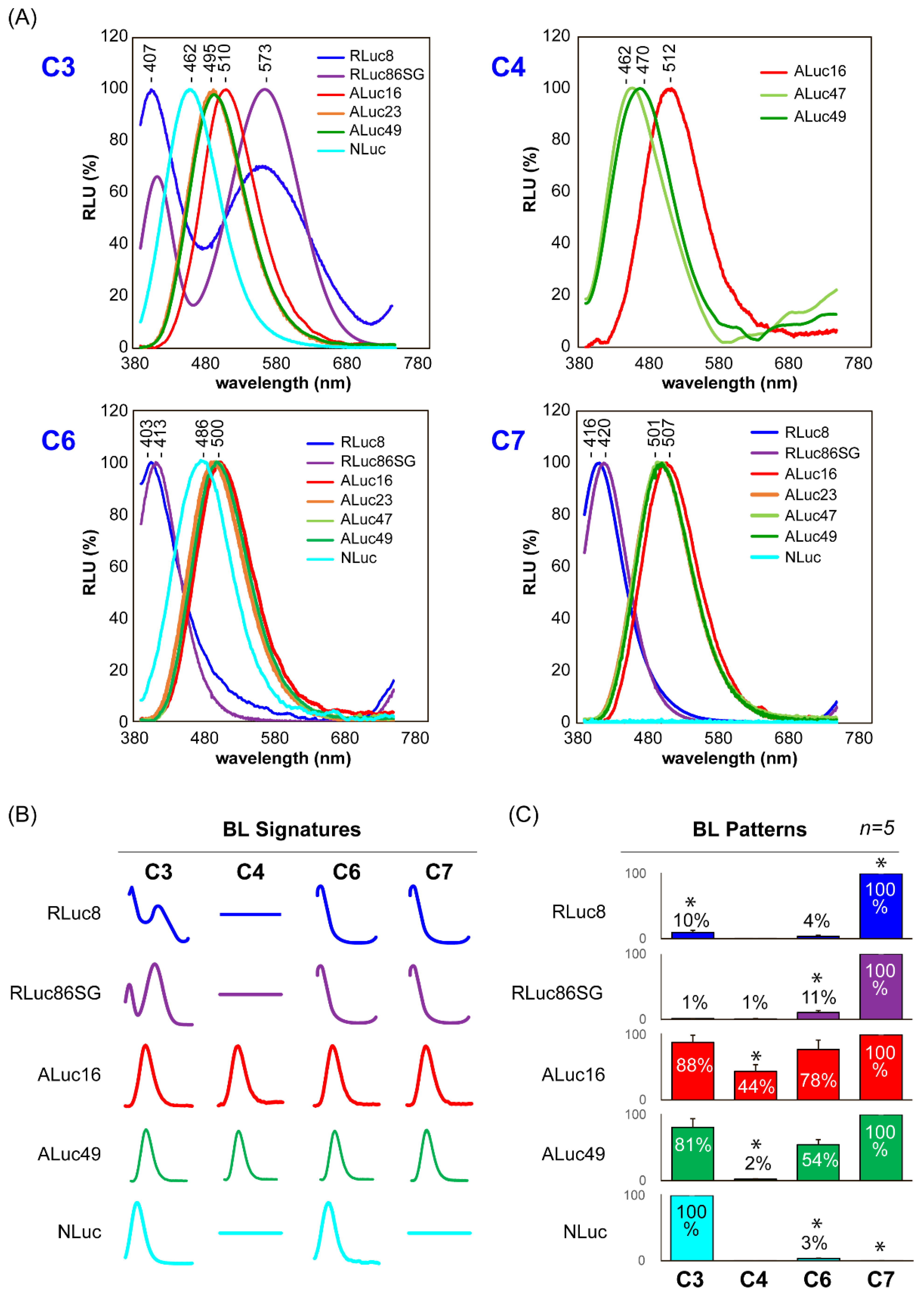
2.6. Characterization of the BL Spectra of the CTZ Analogues C1–C9
2.7. Characterization of the Chemical Stability of the CTZ Analogues C3, C4, C6, and C7
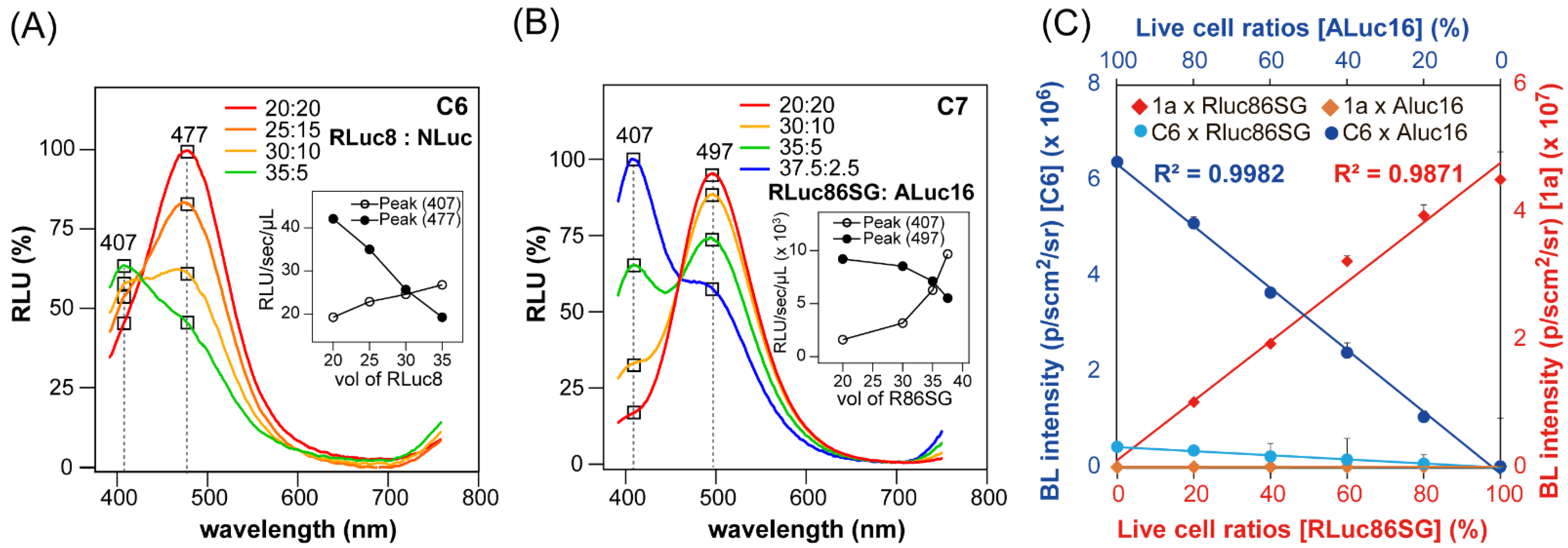
2.8. Quantitative Relationship between Multi-Reporter Systems in Live Cells
2.9. Multi-Probe BL Imaging of F-R8-F and F-A23-F in Live Cells
3. Results and Discussion
3.1. Molecular Designs of the Novel C-Series CTZ Analogues (C1–C9)
3.2. Characterization of the C-Series CTZ Analogues C1–C9
3.3. Characterization of the Autoluminescence Properties of the C-Series CTZ Analogues
3.4. BL Spectral Signatures of the Selected CTZ Analogues C3, C4, C6, and C7
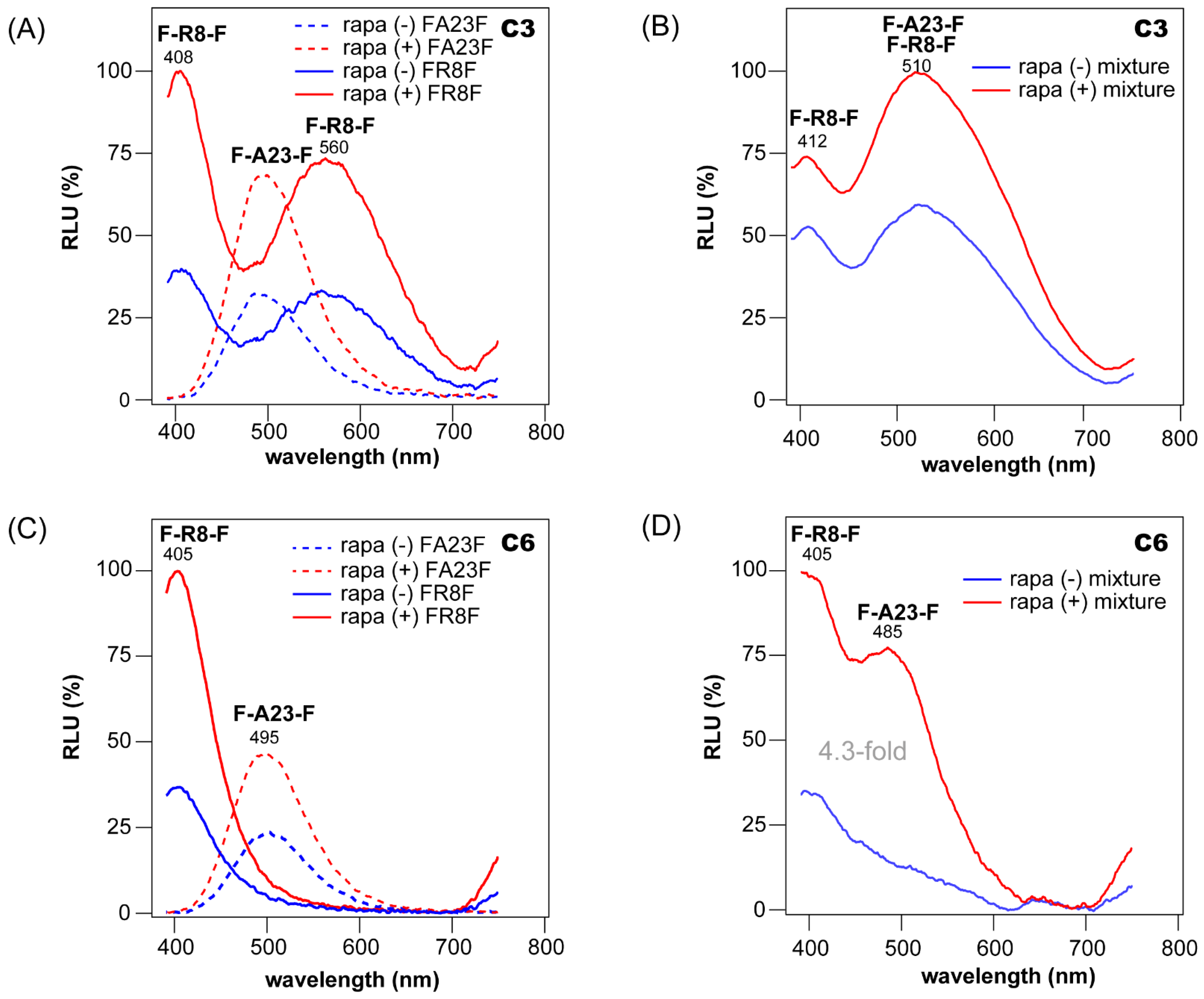
3.5. Quantitative Relationship between Reporters in Multi-Reporter Systems
3.6. Multi-Probe BL Imaging with COS-7 Cells Containing F-R8-F and F-A23-F
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Shimomura, O. Bioluminescence; World Scientific Publishing Co. Pte. Ltd.: Singapore, 2006. [Google Scholar]
- Kim, S.B.; Paulmurugan, R. Bioluminescent Imaging Systems for Assay Developments. Anal. Sci. 2021, 37, 233–247. [Google Scholar] [CrossRef]
- Ozawa, T.; Yoshimura, H.; Kim, S.B. Advances in Fluorescence and Bioluminescence Imaging. Anal. Chem. 2013, 85, 590–609. [Google Scholar] [CrossRef] [PubMed]
- Loening, A.M.; Fenn, T.D.; Wu, A.M.; Gambhir, S.S. Consensus guided mutagenesis of Renilla luciferase yields enhanced stability and light output. Protein Eng. Des. Sel. 2006, 19, 391–400. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.P.; Unch, J.; Binkowski, B.F.; Valley, M.P.; Butler, B.L.; Wood, M.G.; Otto, P.; Zimmerman, K.; Vidugiris, G.; Machleidt, T.; et al. Engineered Luciferase Reporter from a Deep Sea Shrimp Utilizing a Novel Imidazopyrazinone Substrate. ACS Chem. Biol. 2012, 7, 1848–1857. [Google Scholar] [CrossRef] [PubMed]
- Dixon, A.S.; Schwinn, M.K.; Hall, M.P.; Zimmerman, K.; Otto, P.; Lubben, T.H.; Butler, B.L.; Binkowski, B.F.; Machleidt, T.; Kirkland, T.A.; et al. NanoLuc Complementation Reporter Optimized for Accurate Measurement of Protein Interactions in Cells. ACS Chem. Biol. 2016, 11, 400–408. [Google Scholar] [CrossRef]
- Yeh, H.W.; Karmach, O.; Ji, A.; Carter, D.; Martins-Green, M.M.; Ai, H.W. Red-shifted luciferase-luciferin pairs for enhanced bioluminescence imaging. Nat. Methods 2017, 14, 971–974. [Google Scholar] [CrossRef] [PubMed]
- Hiblot, J.; Yu, Q.L.Y.; Sabbadini, M.D.B.; Reymond, L.; Xue, L.; Schena, A.; Sallin, O.; Hill, N.; Griss, R.; Johnsson, K. Luciferases with Tunable Emission Wavelengths. Angew. Chem. Int. Ed. 2017, 56, 14556–14560. [Google Scholar] [CrossRef] [PubMed]
- Westerhausen, S.; Nowak, M.; Torres-Vargas, C.E.; Bilitewski, U.; Bohn, E.; Grin, I.; Wagner, S. A NanoLuc luciferase-based assay enabling the real-time analysis of protein secretion and injection by bacterial type III secretion systems. Mol. Microbiol. 2020, 113, 1240–1254. [Google Scholar] [CrossRef] [Green Version]
- Tamaki, S.; Kitada, N.; Kiyama, M.; Fujii, R.; Hirano, T.; Kim, S.B.; Maki, S. Color-tunable bioluminescence imaging portfolio for cell imaging. Sci. Rep. 2021, 11, 2219. [Google Scholar] [CrossRef]
- Kim, S.B.; Torimura, M.; Tao, H. Creation of artificial luciferases for bioassays. Bioconjug. Chem. 2013, 24, 2067–2075. [Google Scholar] [CrossRef]
- Kim, S.B.; Nishihara, R.; Citterio, D.; Suzuki, K. Fabrication of a New Lineage of Artificial Luciferases from Natural Luciferase Pools. ACS Comb. Sci. 2017, 19, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Gammon, S.T.M.; Leevy, W.M.; Gross, S.; Gokel, G.W.; Piwnica-Worms, D. Spectral unmixing of multicolored bioluminescence emitted from heterogeneous biological sources. Anal. Chem. 2006, 78, 1520–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branchini, B.R.; Southworth, T.L.; Fontaine, D.M.; Kohrt, D.; Florentine, C.M.; Grossel, M.J. A Firefly Luciferase Dual Color Bioluminescence Reporter Assay Using Two Substrates to Simultaneously Monitor Two Gene Expression Events. Sci. Rep. 2018, 8, 5990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.T.; Bai, H.X.; Jiang, T.Y.; Ma, Z.; Cheng, Y.N.; Zhou, Y.B.; Du, L.P.; Li, M.Y. Quenching the firefly bioluminescence by various ions. Photochem. Photobiol. Sci. 2016, 15, 244–249. [Google Scholar] [CrossRef]
- Nishihara, R.; Abe, M.; Nishiyama, S.; Citterio, D.; Suzuki, K.; Kim, S.B. Luciferase-Specific Coelenterazine Analogues for Optical Contamination-Free Bioassays. Sci. Rep. 2017, 7, 908. [Google Scholar] [CrossRef] [Green Version]
- Nishihara, R.; Hoshino, E.; Kakudate, Y.; Kishigami, S.; Iwasawa, N.; Sasaki, S.-I.; Nakajima, T.; Sato, M.; Nishiyama, S.; Citterio, D.; et al. Azide- and Dye-Conjugated Coelenterazine Analogues for a Multiplex Molecular Imaging Platform. Bioconjug. Chem. 2018, 29, 1922–1931. [Google Scholar] [CrossRef]
- Los, G.V.; Encell, L.P.; McDougall, M.G.; Hartzell, D.D.; Karassina, N.; Zimprich, C.; Wood, M.G.; Learish, R.; Ohane, R.F.; Urh, M.; et al. HaloTag: A novel protein labeling technology for cell imaging and protein analysis. ACS Chem. Biol. 2008, 3, 373–382. [Google Scholar] [CrossRef]
- Sarrion-Perdigones, A.; Chang, L.; Gonzalez, Y.; Gallego-Flores, T.; Young, D.W.; Venken, K.J.T. Examining multiple cellular pathways at once using multiplex hextuple luciferase assaying. Nat. Commun. 2019, 10, 5710. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.B.; Sato, M.; Tao, H. Molecular Tension-Indexed Bioluminescent Probe for Determining Protein-Protein Interactions. Bioconjug. Chem. 2009, 20, 2324–2330. [Google Scholar] [CrossRef]
- Kim, S.B.; Nishihara, R.; Citterio, D.; Suzuki, K. Genetically Encoded Molecular Tension Probe for Tracing Protein—Protein Interactions in Mammalian Cells. Bioconjugate Chem. 2016, 27, 354–362. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, N.; Kobayashi, N.; Tsuda, K.; Unzai, S.; Saotome, T.; Kuroda, Y.; Yamazaki, T. Solution structure of Gaussia Luciferase with five disulfide bonds and identification of a putative coelenterazine binding cavity by heteronuclear NMR. Sci. Rep. 2020, 10, 20069. [Google Scholar] [CrossRef] [PubMed]
- Ohmuro-Matsuyama, Y.; Furuta, T.; Matsui, H.; Kanai, M.; Ueda, H. Miniaturization of Bright Light-Emitting Luciferase ALuc: PicALuc. Acs Chem. Biol. 2022, 17, 864–872. [Google Scholar] [CrossRef] [PubMed]
- Schenkmayerova, A.; Toul, M.; Pluskal, D.; Baatallah, R.; Gagnot, G.; Pinto, G.P.; Santana, V.T.; Stuchla, M.; Neugebauer, P.; Chaiyen, P.; et al. A catalytic mechanism for Renilla-type bioluminescence. bioRxiv 2022. bioRxiv:2022.02.09.479090. [Google Scholar]
- Loening, A.M.; Fenn, T.D.; Gambhir, S.S. Crystal structures of the luciferase and green fluorescent protein from Renilla reniformis. J. Mol. Biol. 2007, 374, 1017–1028. [Google Scholar] [CrossRef]
- Nishihara, R.; Paulmurugan, R.; Nakajima, T.; Yamamoto, E.; Natarajan, A.; Afjei, R.; Hiruta, Y.; Iwasawa, N.; Nishiyama, S.; Citterio, D.; et al. Highly bright and stable NIR-BRET with blue-shifted coelenterazine derivatives for deep-tissue imaging of molecular events in vivo. Theranostics 2019, 9, 2646–2661. [Google Scholar] [CrossRef]
- Nishihara, R.; Niwa, K.; Tomita, T.; Kurita, R. Coelenterazine Analogue with Human Serum Albumin-Specific Bioluminescence. Bioconjug. Chem. 2020, 31, 2679–2684. [Google Scholar] [CrossRef]
- Shimomura, O. Cause of Spectral Variation in the Luminescence of Semisynthetic Aequorins. Biochem. J. 1995, 306, 537–543. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Luciferase | Substrates (nm) | ||||
|---|---|---|---|---|---|
| nCTZ | C3 | C4 | C6 | C7 | |
| ALuc16 | 496 | 510 | 512 | 500 | 507 |
| ALuc23 | 500 | 495 | - | 496 | 499 |
| F-A23-F | 500 | 499 | 500 | 498 | 499 |
| ALuc47 | 490 | 490 | 462 | 502 | 501 |
| ALuc49 | 490 | 495 | 470 | 500 | 501 |
| RLuc8 | 480 | 407, 567 | - | 403 | 416 |
| F-R8-F | 490 | 407, 564 | - | 403 | 412 |
| RLuc8.6SG | 535 | 408, 573 | 415 | 413 | 420 |
| NanoLuc | 462 | - | 465 | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamiya, G.; Kitada, N.; Furuta, T.; Hirano, T.; Maki, S.; Kim, S.B. C-Series Coelenterazine-Driven Bioluminescence Signature Imaging. Int. J. Mol. Sci. 2022, 23, 13047. https://doi.org/10.3390/ijms232113047
Kamiya G, Kitada N, Furuta T, Hirano T, Maki S, Kim SB. C-Series Coelenterazine-Driven Bioluminescence Signature Imaging. International Journal of Molecular Sciences. 2022; 23(21):13047. https://doi.org/10.3390/ijms232113047
Chicago/Turabian StyleKamiya, Genta, Nobuo Kitada, Tadaomi Furuta, Takashi Hirano, Shojiro Maki, and Sung Bae Kim. 2022. "C-Series Coelenterazine-Driven Bioluminescence Signature Imaging" International Journal of Molecular Sciences 23, no. 21: 13047. https://doi.org/10.3390/ijms232113047