
Effects and Mechanism of Enhanced UV-B Radiation on the Flag Leaf Angle of Rice

Abstract
:1. Introduction
2. Results
2.1. The Leaf Angle Was Increased by High-Intensity UV-B Radiation
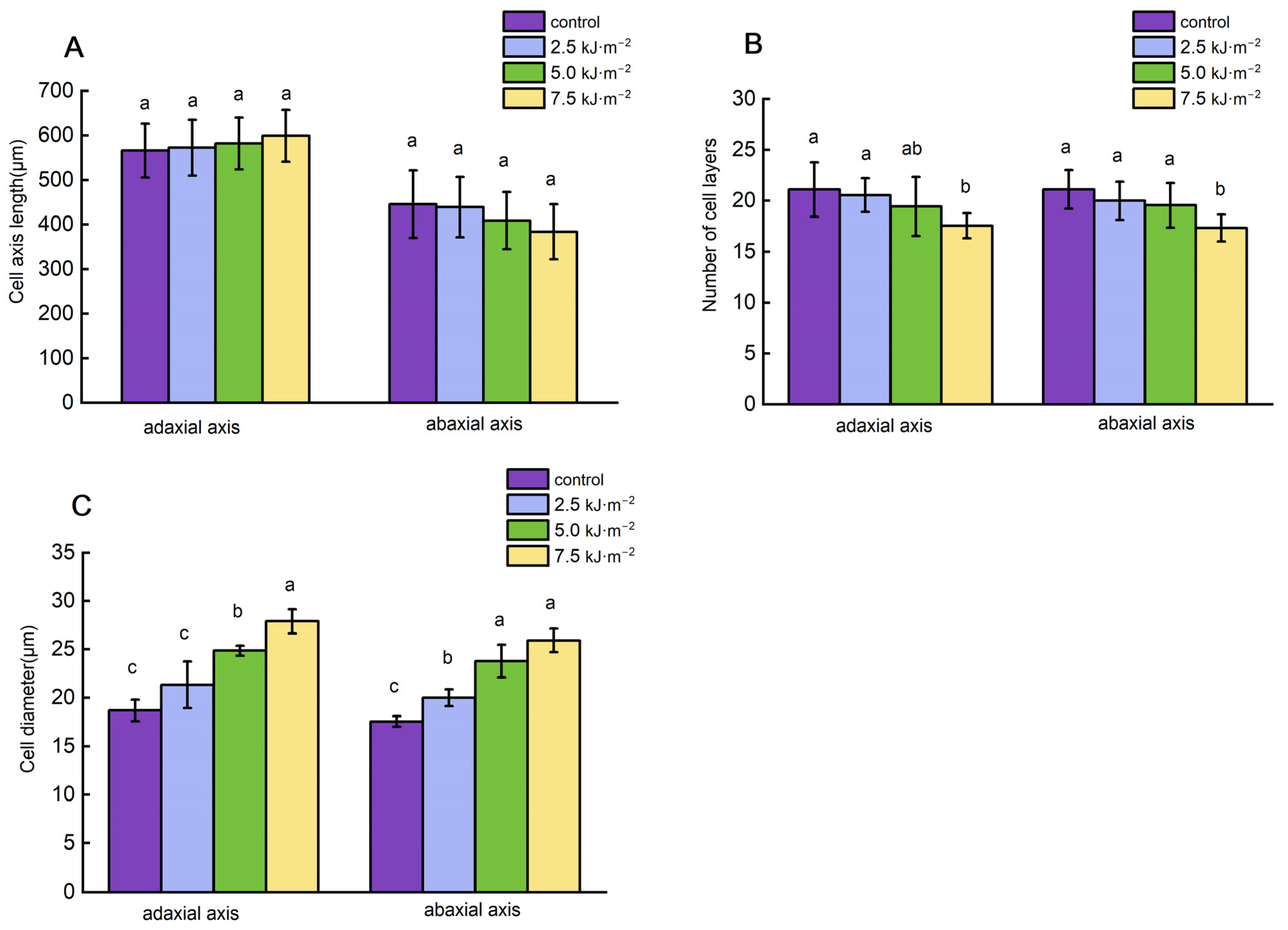
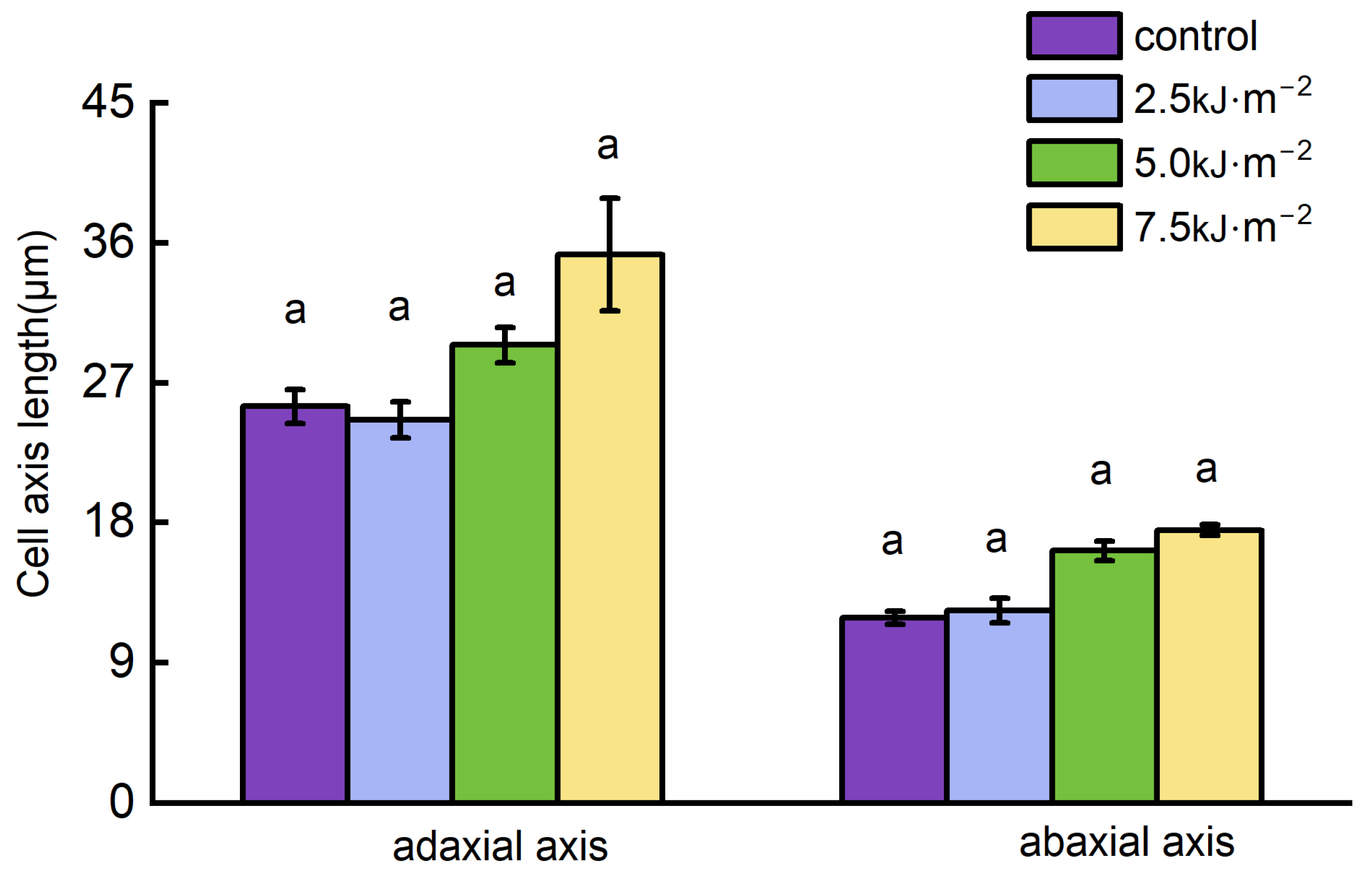
2.2. High-Intensity UV-B Radiation Promotes Cell Elongation and Inhibits Cellulose Synthesis
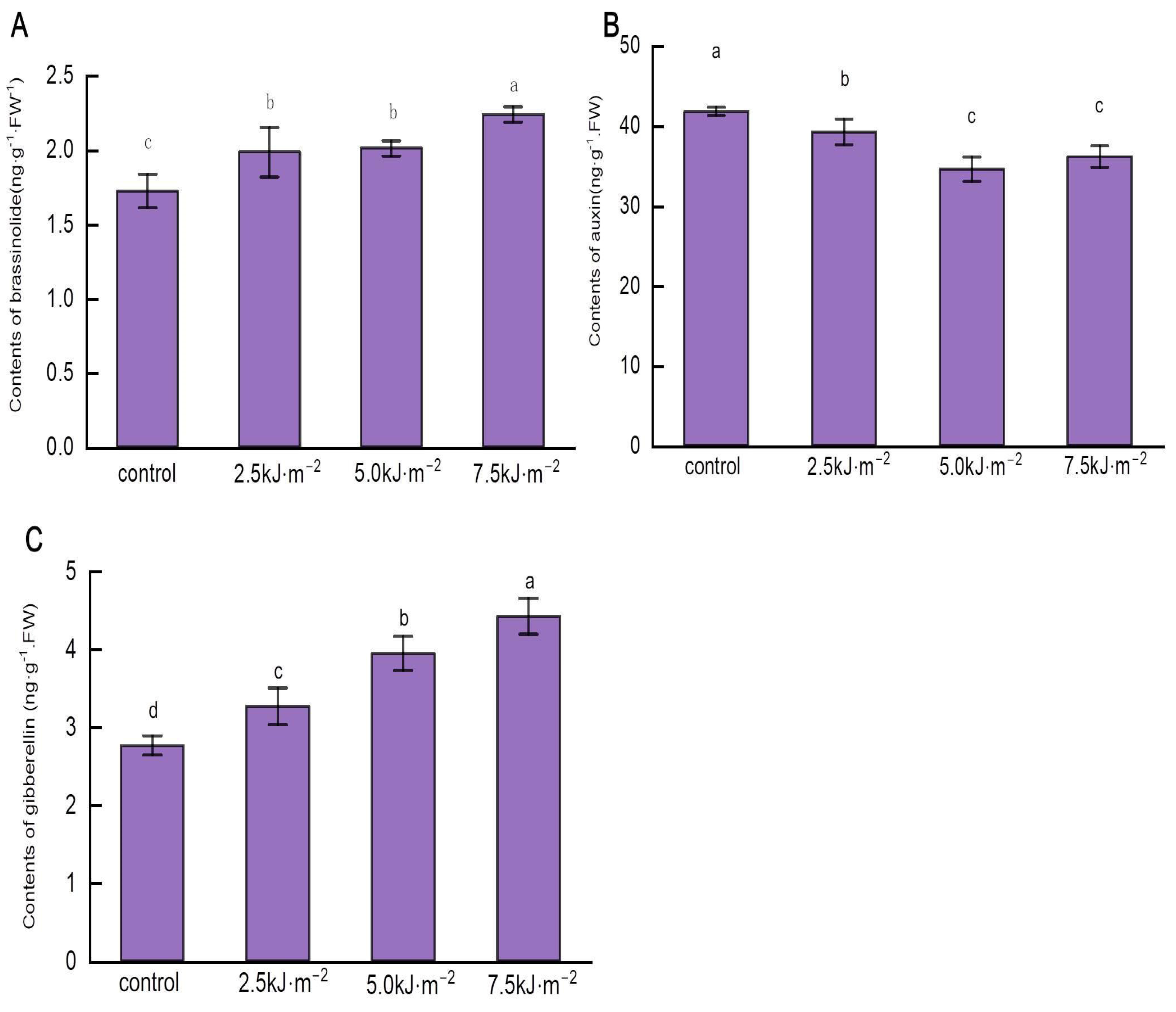
2.3. Radiation with High-Intensity UV-B Positively Regulates the BR and GA Content of the Cusp and Negatively Regulates the IAA Content of the Point
2.4. UV-B Radiation Changes the Content of Hormone Synthase and Cellulose Synthase
2.5. UV-B Radiation Alters the Expression of Hormone-Regulated Genes and Mechano-Tissue-Regulated Genes
3. Discussion
3.1. High-Intensity UV-B Radiation May Damage the Rice Tolerance Mechanism When It Promotes Rice Flag Leaf Angle
3.2. Rice Flag Leaf Angle Is Modulated by UV-B Radiation by Affecting Leaf Pulvini Cell’s Development
3.3. Rice Flag Leaves Are Modulated by UV-B Radiation through a Mechanical Effect on the Leaf Pulvini
3.4. Linkage between the Phytohormonal Pathway and the Mechanical Support Pathway in Regulating the Flag Leaf Angle of Rice under UV-B Radiation
4. Materials and Methods
4.1. Growing Conditions and Plant Material
4.2. UV-B Radiation Treatment
4.3. Measurement of the Flag Leaf Angle
4.4. Preparation of Rice Leaf Pulvini Paraffin Sections
4.5. Determination of Cellulose and Phytohormone Contents, and Their Synthase Activity in Leaf Pulvini
4.6. RNA Extraction and RT-qPCR
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jenkins, G.I. Signal transduction in responses to UV-B radiation. Annu. Rev. Plant Biol. 2009, 60, 407–431. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, L.; Jin, D.; Nezames, C.D.; Terzaghi, W.; Deng, X.W. UV-B-induced photomorphogenesis in Arabidopsis. Protein Cell 2013, 4, 485–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, M.A. Ultraviolet-B radiation effects on plants: Induction of morphogenic responses. Physiol. Plant. 2002, 116, 423–429. [Google Scholar] [CrossRef]
- Hectors, K.; Prinsen, E.; De Coen, W.; Jansen, M.A.K.; Guisez, Y. Arabidopsis thaliana plants acclimated to low dose rates of ultraviolet B radiation show specific changes in morphology and gene expression in the absence of stress symptoms. New Phytol. 2007, 175, 255–270. [Google Scholar] [CrossRef] [PubMed]
- Wargent, J.J.; Gegas, V.C.; Jenkins, G.I.; Doonan, J.H.; Paul, N.D. UVR8 in Arabidopsis thaliana regulates multiple aspects of cellular differentiation during leaf development in response to ultraviolet B radiation. New Phytol. 2009, 183, 315–326. [Google Scholar] [CrossRef]
- Tossi, V.; Lamattina, L.; Jenkins, G.I.; Cassia, R.O. Ultraviolet-B-induced stomatal closure in Arabidopsis is regulated by the UV RESISTANCE LOCUS8 photoreceptor in a nitric oxide-dependent mechanism. Plant Physiol. 2014, 164, 2220–2230. [Google Scholar] [CrossRef] [Green Version]
- Yadav, A.; Singh, D.; Lingwan, M.; Yadukrishnan, P.; Masakapalli, S.K.; Datta, S. Light signaling and UV-B-mediated plant growth regulation. J. Integr. Plant Biol. 2020, 62, 1270–1292. [Google Scholar] [CrossRef]
- Yin, R.; Ulm, R. How plants cope with UV-B: From perception to response. Curr. Opin. Plant Biol. 2017, 37, 42–48. [Google Scholar] [CrossRef] [Green Version]
- Qian, C.; Chen, Z.; Liu, Q.; Mao, W.; Chen, Y.; Tian, W.; Liu, Y.; Han, J.; Ouyang, X.; Huang, X. Coordinated Transcriptional Regulation by the UV-B Photoreceptor and Multiple Transcription Factors for Plant UV-B Responses. Mol. Plant 2020, 13, 777–792. [Google Scholar] [CrossRef]
- Gardner, G.; Lin, C.; Tobin, E.M.; Loehrer, H.; Brinkman, D. Photobiological properties of the inhibition of etiolated Arabidopsis seedling growth by ultraviolet-B irradiation. Plant Cell Environ. 2009, 32, 1573–1583. [Google Scholar] [CrossRef]
- Biever, J.J.; Brinkman, D.; Gardner, G. UV-B inhibition of hypocotyl growth in etiolated Arabidopsis thaliana seedlings is a consequence of cell cycle arrest initiated by photodimer accumulation. J. Exp. Bot. 2014, 65, 2949–2961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumagai, T.; Hidema, J.; Kang, H.-S.; Sato, T. Effects of supplemental UV-B radiation on the growth and yield of two cultivars of Japanese lowland rice (Oryza sativa L.) under the field in a cool rice-growing region of Japan. Agric. Ecosyst. Environ. 2001, 83, 201–208. [Google Scholar] [CrossRef]
- Mohammed, A.; Tarpley, L. Effects of Elevated Ultraviolet-B Radiation on Productive Tillers, Spikelet Sterility and Grain Characteristics of Southern US Rice (Oryza sativa L.) Cultivars. J. Agron. Crop Sci. 2009, 195, 292–300. [Google Scholar] [CrossRef]
- He, Y.M.; Zhan, F.D.; Zu, Y.Q.; Xu, W.W.; Li, Y. Effects of enhanced UV-B radiation on culm charateristics and lodging index of two local rice varieties in Yuanvang terraces under field condition. Ying Yong Sheng Tai Xue Bao 2015, 26, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.; Li, X.; Zhang, Y.; He, Y.; Xie, C.; Li, Y. Effect of UV-B radiation on grain morphology and proanthocyanidins content and distribution of Yuanyang red rice. J. Agro-Environ. Sci. 2019, 38, 2689–2696. [Google Scholar] [CrossRef]
- Bao, L.; He, Y.; Zu, Y.; Li, Y.; Gao, G.; He, Y. Effects of enhanced UV-B radiation on the leaf morphology and anatomical structure of two local rice varieties in Yuanyang terraced fields, Yunnan Province of Southwest China. Chin. J. Ecol. 2013, 32, 882–889. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, X.; Ling, C.; Zu, Y.; He, Y.M.; Yang, Y.; Li, Y. Effects of UV-B radiation on phenolic content and antioxidant capacity of Yuanyang red rice seeds. J. Agro-Environ. Sci. 2022, 41, 10–18. [Google Scholar] [CrossRef]
- Mantilla-Perez, M.B.; Salas Fernandez, M.G. Differential manipulation of leaf angle throughout the canopy: Current status and prospects. J. Exp. Bot. 2017, 68, 5699–5717. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wu, P.; Lu, Y.; Guo, S.; Zhong, Z.; Shen, R.; Xie, Q. Synergistic Interaction of Phytohormones in Determining Leaf Angle in Crops. Int. J. Mol. Sci. 2020, 21, 5052. [Google Scholar] [CrossRef]
- Hu, J.; Lin, H.; Xu, N.; Jiao, R.; Lu, C.; Rao, Y.; Wang, Y. Advances in molecular mechanisms of rice leaf inclination and its application in breeding. Chin. J. Rice Sci. 2019, 33, 391–400. [Google Scholar]
- Das, A.; Pramanik, K.; Sharma, R.; Gantait, S.; Banerjee, J. In-silico study of biotic and abiotic stress-related transcription factor binding sites in the promoter regions of rice germin-like protein genes. PLoS ONE 2019, 14, e0211887. [Google Scholar] [CrossRef] [PubMed]
- He, Z.D.; Tao, M.L.; Leung, D.W.M.; Yan, X.Y.; Chen, L.; Peng, X.X.; Liu, E.E. The rice germin-like protein OsGLP1 participates in acclimation to UV-B radiation. Plant Physiol. 2021, 186, 1254–1268. [Google Scholar] [CrossRef] [PubMed]
- Hong, Z.; Ueguchi-Tanaka, M.; Matsuoka, M. Brassinosteroids and rice architecture. J. Pestic. Sci. 2004, 29, 184–188. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Zheng, J.; Huang, R.; Huang, Y.; Wang, H.; Jiang, L.; Fang, X. Phytohormones signaling and crosstalk regulating leaf angle in rice. Plant Cell Rep. 2016, 35, 2423–2433. [Google Scholar] [CrossRef]
- Qu, L.; Lin, L.B.; Xue, H.W. Rice miR394 suppresses leaf inclination through targeting an F-box gene, leaf inclination 4. J. Integr. Plant Biol. 2019, 61, 406–416. [Google Scholar] [CrossRef]
- Chen, Y.; Dan, Z.; Gao, F.; Chen, P.; Fan, F.; Li, S. Rice Growth-Regulating Factor7 Modulates Plant Architecture through Regulating GA and Indole-3-Acetic Acid Metabolism. Plant Physiol. 2020, 184, 393–406. [Google Scholar] [CrossRef]
- Xu, J.; Wang, L.; Qian, Q.; Zhang, G.-H. Research Advance in Molecule Regulation Mechanism of Leaf Morphogenesis in Rice (Oryza sativa L.). Acta Agron. Sin. 2013, 39, 767. [Google Scholar] [CrossRef]
- Hong, Z.; Ueguchi-Tanaka, M.; Umemura, K.; Uozu, S.; Fujioka, S.; Takatsuto, S.; Yoshida, S.; Ashikari, M.; Kitano, H.; Matsuoka, M. A rice brassinosteroid-deficient mutant, ebisu dwarf (d2), is caused by a loss of function of a new member of cytochrome P450. Plant Cell 2003, 15, 2900–2910. [Google Scholar] [CrossRef] [Green Version]
- Jang, S. A novel trimeric complex in plant cells that contributes to the lamina inclination of rice. Plant Signal. Behav. 2017, 12, e1274482. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.Q.; Xiang, J.J.; Xue, H.W. Studies on the rice LEAF INCLINATION1 (LC1), an IAA-amido synthetase, reveal the effects of auxin in leaf inclination control. Mol. Plant. 2013, 6, 174–187. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Wang, S.; Xu, Y.; Yu, C.; Shen, C.; Qian, Q.; Geisler, M.; de Jiang, A.; Qi, Y. The auxin response factor, OsARF19, controls rice leaf angles through positively regulating OsGH3-5 and OsBRI1. Plant Cell Environ. 2015, 38, 638–654. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, S.; Ashikari, M.; Fujioka, S.; Takatsuto, S.; Yoshida, S.; Yano, M.; Yoshimura, A.; Kitano, H.; Matsuoka, M.; Fujisawa, Y.; et al. A novel cytochrome P450 is implicated in brassinosteroid biosynthesis via the characterization of a rice dwarf mutant, dwarf11, with reduced seed length. Plant Cell 2005, 17, 776–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Wang, Z.; Xu, Y.; Joo, S.H.; Kim, S.K.; Xue, Z.; Xu, Z.; Wang, Z.; Chong, K. OsGSR1 is involved in crosstalk between gibberellins and brassinosteroids in rice. Plant J. 2009, 57, 498–510. [Google Scholar] [CrossRef] [PubMed]
- Ning, J.; Zhang, B.; Wang, N.; Zhou, Y.; Xiong, L. Increased leaf angle1, a Raf-like MAPKKK that interacts with a nuclear protein family, regulates mechanical tissue formation in the Lamina joint of rice. Plant Cell 2011, 23, 4334–4347. [Google Scholar] [CrossRef] [Green Version]
- Robson, T.M.; Klem, K.; Urban, O.; Jansen, M.A. Re-interpreting plant morphological responses to UV-B radiation. Plant Cell Environ. 2015, 38, 856–866. [Google Scholar] [CrossRef]
- Callahan, H.S.; Pigliucci, M. Shade-induced plasticity and its ecological significance in wild populations of arabidopsis thaliana. Ecology 2002, 83, 1965–1980. [Google Scholar] [CrossRef]
- Deckmyn, G.; Cayenberghs, E.; Ceulemans, R. UV-B and PAR in single and mixed canopies grown under different UV-B exclusions in the field. Plant Ecol. 2001, 154, 123–133. [Google Scholar] [CrossRef]
- Shi, C.; Liu, H. How plants protect themselves from ultraviolet-B radiation stress. Plant Physiol. 2021, 187, 1096–1103. [Google Scholar] [CrossRef]
- Sasaki, T.; Akutsu, H.; Shimada, H.; Miura, S. A rice cytochrome P450 OsCYP84A that may interact with the UV tolerance pathway. Biosci. Biotechnol. Biochem. 2010, 74, 1045–1049. [Google Scholar] [CrossRef] [Green Version]
- Nawkar, G.M.; Maibam, P.; Park, J.H.; Sahi, V.P.; Lee, S.Y.; Kang, C.H. UV-Induced cell death in plants. Int. J. Mol. Sci. 2013, 14, 1608–1628. [Google Scholar] [CrossRef] [Green Version]
- Tena, G.; Asai, T.; Chiu, W.L.; Sheen, J. Plant mitogen-activated protein kinase signaling cascades. Curr. Opin. Plant Biol. 2001, 4, 392–400. [Google Scholar] [CrossRef]
- Nakagami, H.; Pitzschke, A.; Hirt, H. Emerging MAP kinase pathways in plant stress signalling. Trends Plant Sci. 2005, 10, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Jonak, C.; Okrész, L.; Bögre, L.; Hirt, H. Complexity, cross talk and integration of plant MAP kinase signalling. Curr. Opin. Plant Biol. 2002, 5, 415–424. [Google Scholar] [CrossRef]
- Sugimoto-Shirasu, K.; Roberts, K. “Big it up”: Endoreduplication and cell-size control in plants. Curr. Opin. Plant Biol. 2003, 6, 544–553. [Google Scholar] [CrossRef]
- Lake, J.A.; Field, K.J.; Davey, M.P.; Beerling, D.J.; Lomax, B.H. Metabolomic and physiological responses reveal multi-phasic acclimation of Arabidopsis thaliana to chronic UV radiation. Plant Cell Environ. 2009, 32, 1377–1389. [Google Scholar] [CrossRef]
- Fina, J.; Casadevall, R.; AbdElgawad, H.; Prinsen, E.; Markakis, M.N.; Beemster, G.T.S.; Casati, P. UV-B Inhibits Leaf Growth through Changes in Growth Regulating Factors and Gibberellin Levels. Plant Physiol. 2017, 174, 1110–1126. [Google Scholar] [CrossRef] [Green Version]
- Wan, J.; Zhang, P.; Wang, R.; Sun, L.; Wang, W.; Zhou, H.; Xu, J. UV-B Radiation Induces Root Bending Through the Flavonoid-Mediated Auxin Pathway in Arabidopsis. Front. Plant Sci. 2018, 9, 618. [Google Scholar] [CrossRef]
- Idris, M.; Seo, N.; Jiang, L.; Kiyota, S.; Hidema, J.; Iino, M. UV-B signalling in rice: Response identification, gene expression profiling and mutant isolation. Plant Cell Environ. 2021, 44, 1468–1485. [Google Scholar] [CrossRef]
- Mitrović, A.L.; Radosavljević, J.S.; Prokopijević, M.; Spasojević, D.; Kovačević, J.; Prodanović, O.; Todorović, B.; Matović, B.; Stanković, M.; Maksimović, V.; et al. Cell wall response to UV radiation in needles of Picea omorika. Plant. Physiol. Biochem. 2021, 161, 176–190. [Google Scholar] [CrossRef]
- Huang, G.; Hu, H.; van de Meene, A.; Zhang, J.; Dong, L.; Zheng, S.; Zhang, F.; Betts, N.S.; Liang, W.; Bennett, M.J.; et al. Auxin response factors 6 and 17 control the flag leaf angle in rice by regulating secondary cell wall biosynthesis of lamina joints. Plant Cell 2021, 33, 3120–3133. [Google Scholar] [CrossRef]
- Liu, S.; Liu, N.; Lu, H.; Zhu, L. Disturbed phospholipid metabolism by three polycyclic aromatic hydrocarbons in Oryza sativa. Environ. Pollut. 2021, 283, 117073. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | 5′--3′ | Tm (°C) | Product (bp) |
|---|---|---|---|
| ILAL-F | GCGGAAGGTCAGGAGAGAAC | 58.6 | 142 |
| ILAL-R | ACTGGCGTTGAAGCGGTG | 59.9 | |
| D2-F | TGGAGGTGGAAGGAGAAGGA | 59.2 | 172 |
| D2-R | TGGGGAAGTTGACGATGTGG | 61.1 | |
| OsBUL1-F | GGTTTTTCCCTCTCCCTTCCA | 62 | 91 |
| OsBUL1-R | GTAGTGGTCGGTCGGTTGTC | 57.4 | |
| OsARF19-F | CGAAGGATGCCCAGCAAGAA | 63.4 | 348 |
| OsARF19-R | CGTCGCCAAGAAGTAGGATGT | 59.4 | |
| OsGSR1-F | CTGCCCCTGCTACAACAACT | 57.7 | 296 |
| OsGSR1-R | CTCACTCTCCTCTCTCGCAC | 54.2 | |
| TBC1 | TGGTCATGTTCCTTCAGCAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ling, C.; Wang, X.; Li, Z.; He, Y.; Li, Y. Effects and Mechanism of Enhanced UV-B Radiation on the Flag Leaf Angle of Rice. Int. J. Mol. Sci. 2022, 23, 12776. https://doi.org/10.3390/ijms232112776
Ling C, Wang X, Li Z, He Y, Li Y. Effects and Mechanism of Enhanced UV-B Radiation on the Flag Leaf Angle of Rice. International Journal of Molecular Sciences. 2022; 23(21):12776. https://doi.org/10.3390/ijms232112776
Chicago/Turabian StyleLing, Chengting, Xiupin Wang, Zuran Li, Yongmei He, and Yuan Li. 2022. "Effects and Mechanism of Enhanced UV-B Radiation on the Flag Leaf Angle of Rice" International Journal of Molecular Sciences 23, no. 21: 12776. https://doi.org/10.3390/ijms232112776