

Genome-Wide Identification and Expression Analysis of Senescence-Associated Genes in Grapevine (Vitis vinifera L.) Reveal Their Potential Functions in Leaf Senescence Order

Abstract
:1. Introduction
2. Results
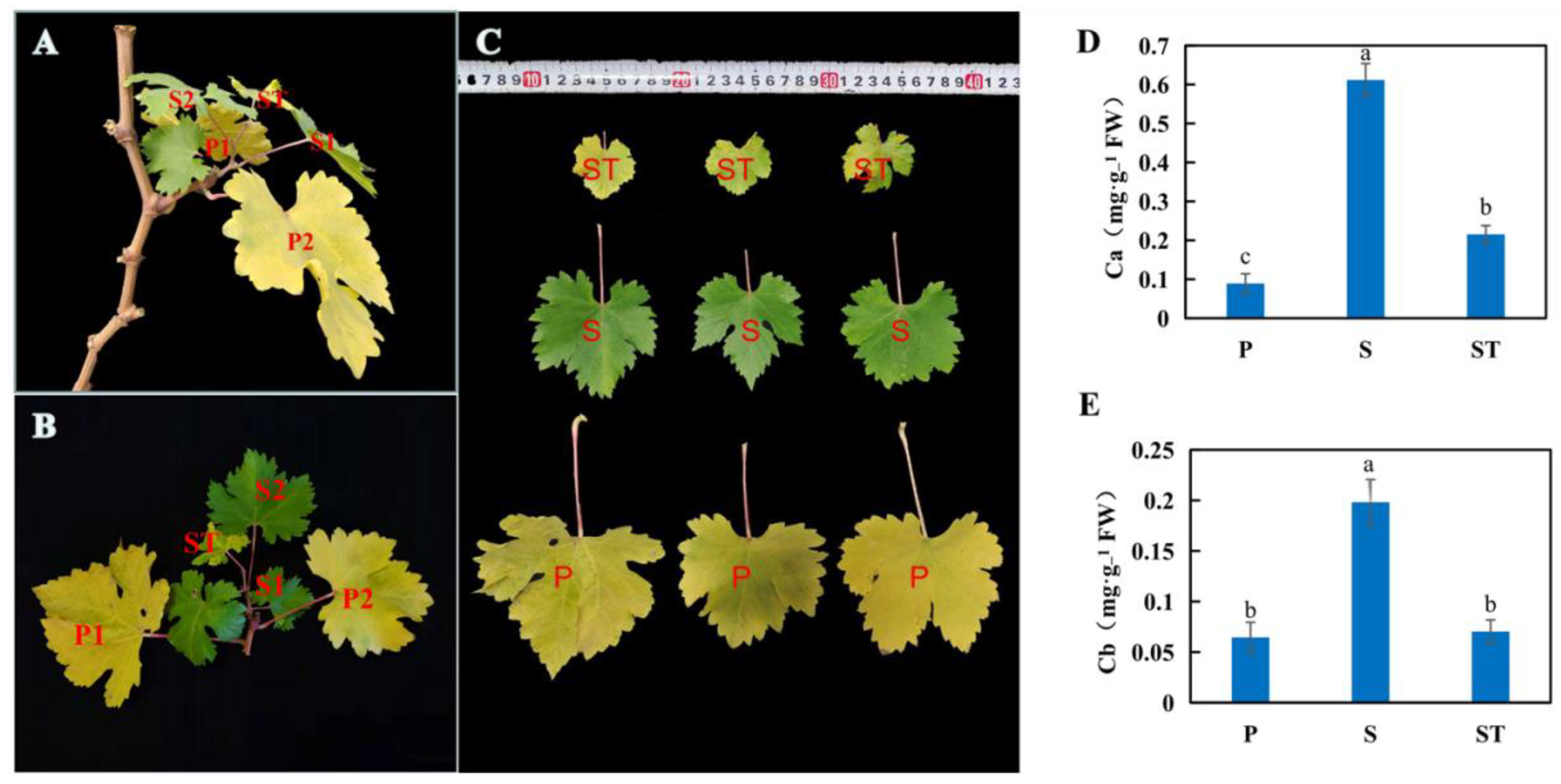
2.1. The Impact of Leaf Position on Senescence of Grapevine Leaves
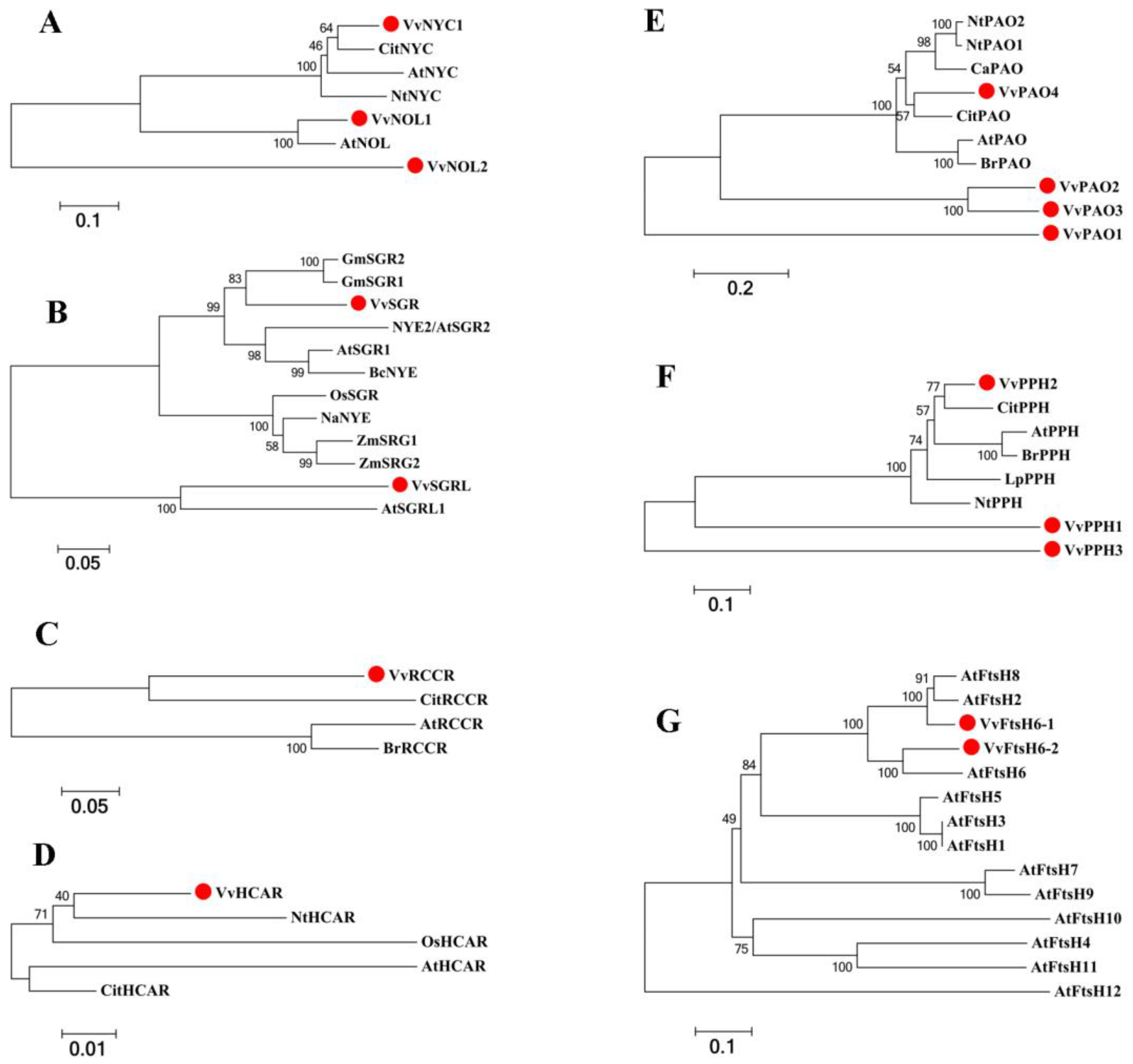
2.2. Genome-Wide Identification and Characterization of Chl Degradation-Related SAGs in Grapevine Leaves
2.3. Cis-Acting Regulatory Element Analysis of SAG Promoters
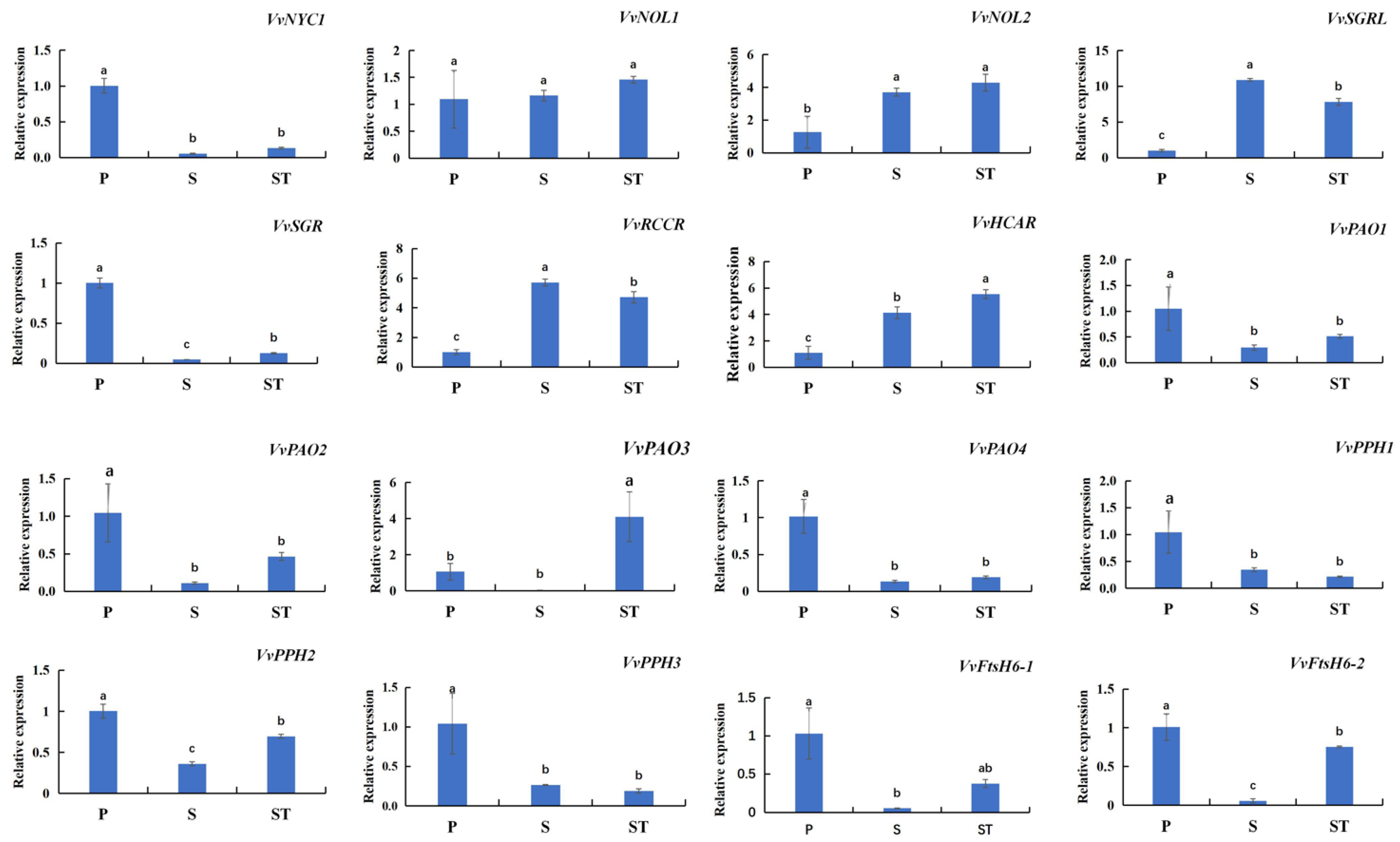
2.4. Chl Degradation-Related SAGs Are Characterized by Diverse Expression Profiles in Different Leaf Positions of Grape Shoots
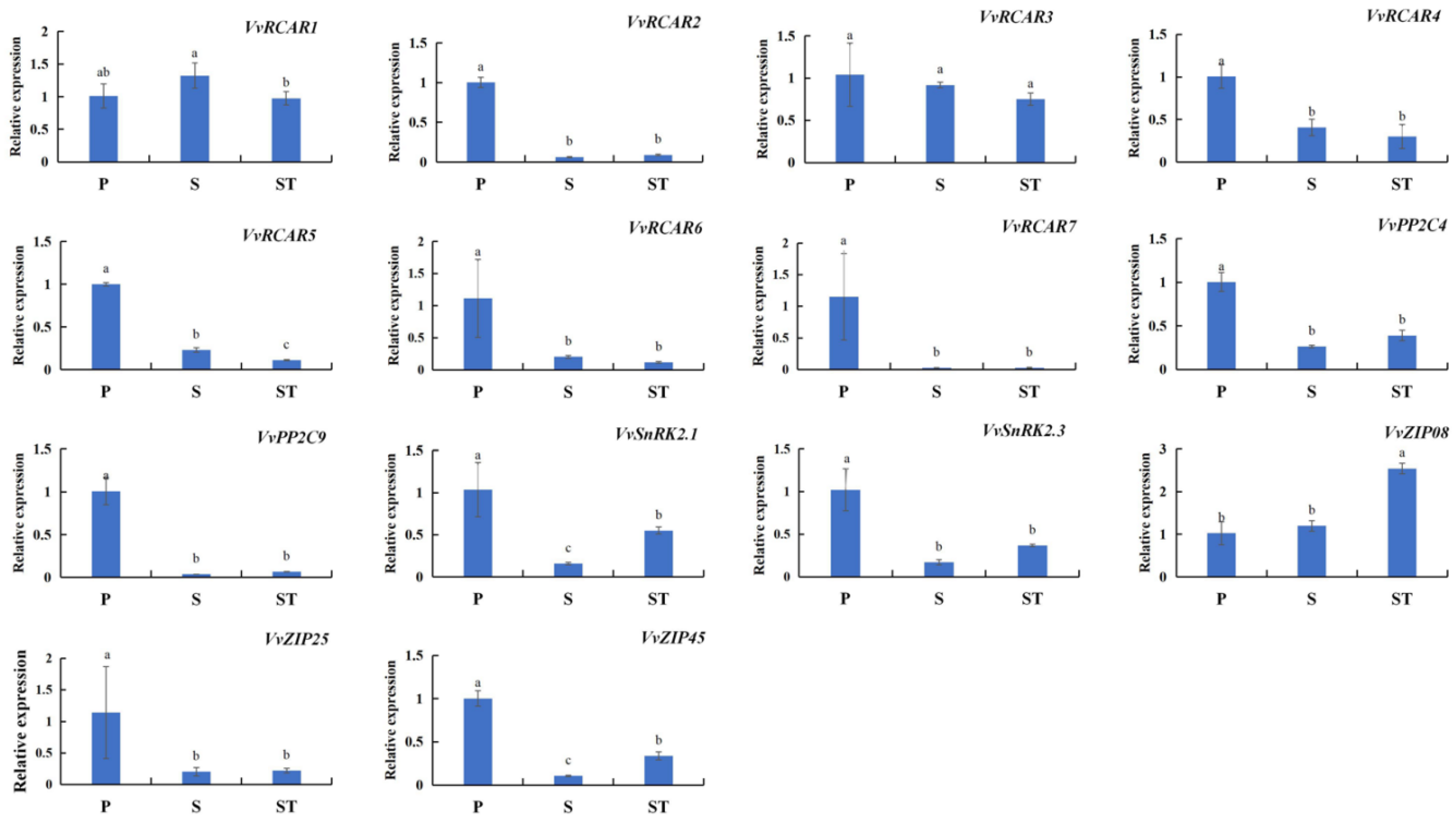
2.5. ABA-Related SAGs Are Characterized by Consistent Expression Profiles with Chl Degradation-Related SAGs in Different Leaf Positions of Grape Shoots
3. Discussion
4. Materials and Methods
4.1. Plant Material and Sampling
4.2. Measurement of Chl Content
4.3. Identification of Chl Degradation-Related SAGs in Grapevine Leaves
4.4. Phylogenetic Analysis, Gene Structure Analysis, and Promoter Analysis
4.5. Expression Analysis by Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schippers, J.H.; Schmidt, R.; Wagstaff, C.; Jing, H.C. Living to Die and Dying to Live: The Survival Strategy behind Leaf Senescence. Plant Physiol. 2015, 169, 914–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schippers, J.H. Transcriptional networks in leaf senescence. Curr. Opin. Plant Biol. 2015, 27, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Noodén, L.D. Plant Cell Death Processes; Academic Press: London, UK, 2004. [Google Scholar]
- Tamary, E.; Nevo, R.; Naveh, L.; Levin-Zaidman, S.; Kiss, V.; Savidor, A.; Levin, Y.; Eyal, Y.; Reich, Z.; Adam, Z. Chlorophyll catabolism precedes changes in chloroplast structure and proteome during leaf senescence. Plant Direct 2019, 3, e00127. [Google Scholar] [CrossRef] [PubMed]
- Horie, Y.; Ito, H.; Kusaba, M.; Tanaka, R.; Tanaka, A. Participation of chlorophyll b reductase in the initial step of the degradation of light-harvesting chlorophyll a/b-protein complexes in Arabidopsis. J. Biol. Chem. 2009, 284, 17449–17456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meguro, M.; Ito, H.; Takabayashi, A.; Tanaka, R.; Tanaka, A. Identification of the 7-hydroxymethyl chlorophyll a reductase of the chlorophyll cycle in Arabidopsis. Plant Cell 2011, 23, 3442–3453. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Morita, R.; Katsuma, S.; Nishimura, M.; Tanaka, A.; Kusaba, M. Two short-chain dehydrogenase/reductases, NON-YELLOW COLORING 1 and NYC1-LIKE, are required for chlorophyll b and light-harvesting complex II degradation during senescence in rice. Plant J. 2009, 57, 120–131. [Google Scholar] [CrossRef]
- Morita, R.; Sato, Y.; Masuda, Y.; Nishimura, M.; Kusaba, M. Defect in non-yellow coloring 3, an alpha/beta hydrolase-fold family protein, causes a stay-green phenotype during leaf senescence in rice. Plant J. 2009, 59, 940–952. [Google Scholar] [CrossRef]
- Pruzinska, A.; Anders, I.; Aubry, S.; Schenk, N.; Tapernoux-Luthi, E.; Muller, T.; Krautler, B.; Hortensteiner, S. In Vivo participation of red chlorophyll catabolite reductase in chlorophyll breakdown. Plant Cell 2007, 19, 369–387. [Google Scholar] [CrossRef] [Green Version]
- Piao, W.; Han, S.H.; Sakuraba, Y.; Paek, N.C. Rice 7-Hydroxymethyl Chlorophyll a Reductase Is Involved in the Promotion of Chlorophyll Degradation and Modulates Cell Death Signaling. Mol. Cell 2017, 40, 773–786. [Google Scholar]
- Sakuraba, Y.; Kim, Y.S.; Yoo, S.C.; Hortensteiner, S.; Paek, N.C. 7-Hydroxymethyl chlorophyll a reductase functions in metabolic channeling of chlorophyll breakdown intermediates during leaf senescence. Biochem. Biophys. Res. Commun. 2013, 430, 32–37. [Google Scholar] [CrossRef] [Green Version]
- Schelbert, S.; Aubry, S.; Burla, B.; Agne, B.; Kessler, F.; Krupinska, K.; Hortensteiner, S. Pheophytin Pheophorbide Hydrolase (Pheophytinase) Is Involved in Chlorophyll Breakdown during Leaf Senescence in Arabidopsis. Plant Cell 2009, 21, 767–785. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.H.; Xie, Z.I.; Zhang, J.; Lei, S.S.; Lin, W.J.; Xu, B.; Huang, B.R. NOL-mediated functional stay-green traits in perennial ryegrass (Lolium perenne L.) involving multifaceted molecular factors and metabolic pathways regulating leaf senescence. Plant J. 2021, 106, 1219–1232. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.D.; An, K.; Liao, Y.; Zhou, X.; Cao, Y.J.; Zhao, H.F.; Ge, X.C.; Kuai, B.K. Identification of a novel chloroplast protein AtNYE1 regulating chlorophyll degradation during leaf senescence in arabidopsis. Plant Physiol. 2007, 144, 1429–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakuraba, Y.; Schelbert, S.; Park, S.Y.; Han, S.H.; Lee, B.D.; Andres, C.B.; Kessler, F.; Hortensteiner, S.; Paek, N.C. STAY-GREEN and Chlorophyll Catabolic Enzymes Interact at Light-Harvesting Complex II for Chlorophyll Detoxification during Leaf Senescence in Arabidopsis. Plant Cell 2012, 24, 507–518. [Google Scholar] [CrossRef] [Green Version]
- Sakuraba, Y.; Kim, D.; Kim, Y.S.; Hortensteiner, S.; Paek, N.C. Arabidopsis STAYGREEN-LIKE (SGRL) promotes abiotic stress-induced leaf yellowing during vegetative growth. FEBS Lett. 2014, 588, 3830–3837. [Google Scholar] [CrossRef] [Green Version]
- Sakuraba, Y.; Park, S.Y.; Kim, Y.S.; Wang, S.H.; Yoo, S.C.; Hortensteiner, S.; Paek, N.C. Arabidopsis STAY-GREEN2 Is a Negative Regulator of Chlorophyll Degradation during Leaf Senescence. Mol. Plant 2014, 7, 1288–1302. [Google Scholar] [CrossRef] [Green Version]
- Wagner, R.; Aigner, H.; Funk, C. FtsH proteases located in the plant chloroplast. Physiol. Plant 2012, 145, 203–214. [Google Scholar] [CrossRef]
- Zelisko, A.; Garcia-Lorenzo, M.; Jackowski, G.; Jansson, S.; Funk, C. AtFtsH6 is involved in the degradation of the light-harvesting complex II during high-light acclimation and senescence. Proc. Natl. Acad. Sci. USA 2005, 102, 13699–13704. [Google Scholar] [CrossRef] [Green Version]
- Ueda, H.; Ito, T.; Inoue, R.; Masuda, Y.; Nagashima, Y.; Kozuka, T.; Kusaba, M. Genetic Interaction Among Phytochrome, Ethylene and Abscisic Acid Signaling During Dark-Induced Senescence in Arabidopsis thaliana. Front. Plant Sci. 2020, 11, 564. [Google Scholar] [CrossRef]
- Gao, S.; Gao, J.; Zhu, X.; Song, Y.; Li, Z.; Ren, G.; Zhou, X.; Kuai, B. ABF2, ABF3, and ABF4 Promote ABA-Mediated Chlorophyll Degradation and Leaf Senescence by Transcriptional Activation of Chlorophyll Catabolic Genes and Senescence-Associated Genes in Arabidopsis. Mol. Plant 2016, 9, 1272–1285. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Chan, Z.; Gao, J.; Xing, L.; Cao, M.; Yu, C.; Hu, Y.; You, J.; Shi, H.; Zhu, Y.; et al. ABA receptor PYL9 promotes drought resistance and leaf senescence. Proc. Natl. Acad. Sci. USA 2016, 113, 1949–1954. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.Y.; Kuai, B.K.; Jia, J.Z.; Jing, H.C. Regulation of leaf senescence and crop genetic improvement. J. Integr. Plant Biol. 2012, 54, 936–952. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.G.; Li, G.R.; Yan, C.H.; Liu, L.; Zhang, Q.T.; Han, Z.; Li, B. DRL1, Encoding A NAC Transcription Factor, Is Involved in Leaf Senescence in Grapevine. Int. J. Mol. Sci. 2019, 20, 2678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, X.R.; Xie, X.L.; Xia, X.J.; Yu, J.Q.; Ferguson, I.B.; Giovannoni, J.J.; Chen, K.S. Involvement of an ethylene response factor in chlorophyll degradation during citrus fruit degreening. Plant J. 2016, 86, 403–412. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhang, Y.; Zou, D.; Zhao, Y.; Wang, H.L.; Zhang, Y.; Xia, X.; Luo, J.; Guo, H.; Zhang, Z. LSD 3.0: A comprehensive resource for the leaf senescence research community. Nucleic Acids Res. 2020, 48, D1069–D1075. [Google Scholar] [CrossRef]
- Tanaka, R.; Kobayashi, K.; Masuda, T. Tetrapyrrole metabolism in arabidopsis thaliana. Arab. Book 2011, 9, e0145. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, R.; Hirashima, M.; Satoh, S.; Tanaka, A. The Arabidopsis-accelerated cell death gene ACD1 is involved in oxygenation of pheophorbide a: Inhibition of the pheophorbide a oxygenase activity does not lead to the“Stay-Green” phenotype in Arabidopsis. Plant Cell Physiol. 2003, 44, 1266–1274. [Google Scholar] [CrossRef] [Green Version]
- Mach, J.M.; Castillo, A.R.; Hoogstraten, R.; Greenberg, J.T. The Arabidopsis-accelerated cell death gene ACD2 encodes red chlorophyll catabolite reductase and suppresses the spread of disease symptoms. Proc. Natl. Acad. Sci. USA 2001, 98, 771–776. [Google Scholar] [CrossRef]
- Zhang, J.; Yu, G.H.; Wen, W.W.; Ma, X.Q.; Xu, B.; Huang, B.R. Functional characterization and hormonal regulation of the PHEOPHYTINASE gene LpPPH controlling leaf senescence in perennial ryegrass. J. Exp. Bot. 2016, 67, 935–945. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.I.; Hong, J.H.; Ha, J.O.; Kang, J.Y.; Kim, S.Y. ABFs, a family of ABA-responsive element binding factors. J. Biol. Chem. 2000, 275, 1723–1730. [Google Scholar] [CrossRef] [Green Version]
- Uno, Y.; Furihata, T.; Abe, H.; Yoshida, R.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis basic leucine zipper transcription factors involved in an abscisic acid-dependent signal transduction pathway under drought and high-salinity conditions. Proc. Natl. Acad Sci. USA 2000, 97, 11632–11637. [Google Scholar] [CrossRef] [PubMed]
- Boneh, U.; Biton, I.; Zheng, C.; Schwartz, A.; Ben-Ari, G. Characterization of potential ABA receptors in Vitis vinifera. Plant Cell Rep. 2012, 31, 311–321. [Google Scholar] [CrossRef]
- Liu, J.; Chen, N.; Chen, F.; Cai, B.; Dal Santo, S.; Tornielli, G.B.; Pezzotti, M.; Cheng, Z.M. Genome-wide analysis and expression profile of the bZIP transcription factor gene family in grapevine (Vitis vinifera). BMC Genom. 2014, 15, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boneh, U.; Biton, I.; Schwartz, A.; Ben-Ari, G. Characterization of the ABA signal transduction pathway in Vitis vinifera. Plant Sci. 2012, 187, 89–96. [Google Scholar] [CrossRef]
- Burnison, B.K. Modified dimethyl sulfoxide (dmso) extraction for chlorophyll analysis of phytoplankton. Can. J. Fish. Aquat. 2011, 37, 729–733. [Google Scholar] [CrossRef]
- Jaillon, O.; Aury, J.M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463. [Google Scholar]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.Y.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Gambino, G.; Perrone, I.; Gribaudo, I. A Rapid and effective method for RNA extraction from different tissues of grapevine and other woody plants. Phytochem. Anal. 2008, 19, 520–525. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Location | Peptide (aa) |
|---|---|---|---|
| VvNYC1 | VIT_11s0016g03890.2 | chr11:3174337..3179203− | 518 |
| VvNOL1 | VIT_01s0010g00590.1 | chr1:15587022..15619072− | 317 |
| VvNOL2 | VIT_12s0035g01780.1 | chr12:21916237..21924762− | 265 |
| VvSGR | VIT_02s0025g04660.1 | chr2:4225612..4232509+ | 228 |
| VvSGRL | VIT_18s0001g01210.1 | chr18:1811441..1814300− | 252 |
| VvRCCR | VIT_07s0031g00680.1 | chr7:16847972..1685168+ | 323 |
| VvHCAR | VIT_05s0051g00070.2 | chr5:10300193..10315728− | 458 |
| VvPAO1 | VIT_04s0008g07020.1 | chr4:7106897..7110246− | 545 |
| VvPAO2 | VIT_06s0004g00610.1 | chr6:769345..772779− | 524 |
| VvPAO3 | VIT_06s0004g00620.1 | chr6:780506..783167− | 464 |
| VvPAO4 | VIT_06s0061g00790.1 | chr6:18321351..18327161− | 540 |
| VvPPH1 | VIT_04s0023g02010.1 | chr4:18547350..18550496+ | 368 |
| VvPPH2 | VIT_13s0158g00180.2 | chr13:21072015..21076223− | 525 |
| VvPPH3 | VIT_16s0022g01340.2 | chr6:780506..783167− | 464 |
| VvFtsH6-1 | VIT_12s0028g01600.1 | chr12:2304815..2308584+ | 695 |
| VvFtsH6-2 | VIT_14s0108g00590.1 | chr14:29333100..29335715+ | 677 |
| Element | Function | Cis-Element on SAGs Promoters | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NYC1 | NOL1 | NOL2 | SGRL | SGR | RCCR | HCAR | PAO1 | PAO2 | PAO3 | PAO4 | PPH1 | PPH2 | PPH3 | FtsH6-1 | FtsH6-2 | ||
| ABRE | Abscisic acid responsiveness | + | + | + | + | + | + | + | + | + | + | + | + | ||||
| ACE | Light responsiveness | + | + | ||||||||||||||
| AE-box | Light response | + | + | + | + | + | + | ||||||||||
| ARE | Anaerobic induction | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| CAAT-box | Common cis-acting element in promoter and enhancer regions | + | + | + | + | + | + | + | + | + | + | + | + | ||||
| AuxRR-core | Auxin responsiveness | + | + | ||||||||||||||
| CGTCA-motif | MeJA-responsiveness | + | + | + | + | + | |||||||||||
| G-box | Light responsiveness | + | + | + | + | + | + | + | + | + | + | ||||||
| TATC-box | Gibberellin-responsiveness | + | + | + | + | ||||||||||||
| TC-rich repeats | Defense and stress responsiveness | + | + | + | |||||||||||||
| TCA-element | Salicylic acid responsiveness | + | + | + | + | + | + | + | + | + | + | ||||||
| TCT-motif | Part of a light-responsive element | + | + | + | + | + | + | + | + | + | |||||||
| TGACG-motif | MeJA-responsiveness | + | + | + | + | + | + | ||||||||||
| Box4 | Light responsiveness | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| GARE-motif | Gibberellin-responsive element | + | + | + | + | + | + | + | |||||||||
| P-box | Gibberellin-responsive element | + | + | + | + | + | + | ||||||||||
| GATA-motif | Part of a light-responsive element | + | + | + | |||||||||||||
| MRE | MYB binding site involved in light responsiveness | + | + | + | + | ||||||||||||
| AT1-motif | Part of a light-responsive module | + | + | + | |||||||||||||
| ATCT-motif | Light responsiveness | + | + | ||||||||||||||
| I-box | Light-responsive element | + | + | ||||||||||||||
| chs-CMA2a | Light-responsive element | + | + | + | |||||||||||||
| ABRE3a | + | + | + | + | + | + | + | + | |||||||||
| ABRE4 | + | + | + | + | + | + | + | + | |||||||||
| TCCC-motif | Light-responsive element | + | + | + | + | + | |||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.-M.; Sun, M.-H.; Tang, X.-S.; Wang, C.-P.; Xie, Z.-S. Genome-Wide Identification and Expression Analysis of Senescence-Associated Genes in Grapevine (Vitis vinifera L.) Reveal Their Potential Functions in Leaf Senescence Order. Int. J. Mol. Sci. 2022, 23, 12731. https://doi.org/10.3390/ijms232112731
Li Y-M, Sun M-H, Tang X-S, Wang C-P, Xie Z-S. Genome-Wide Identification and Expression Analysis of Senescence-Associated Genes in Grapevine (Vitis vinifera L.) Reveal Their Potential Functions in Leaf Senescence Order. International Journal of Molecular Sciences. 2022; 23(21):12731. https://doi.org/10.3390/ijms232112731
Chicago/Turabian StyleLi, You-Mei, Meng-Hao Sun, Xuan-Si Tang, Chao-Ping Wang, and Zhao-Sen Xie. 2022. "Genome-Wide Identification and Expression Analysis of Senescence-Associated Genes in Grapevine (Vitis vinifera L.) Reveal Their Potential Functions in Leaf Senescence Order" International Journal of Molecular Sciences 23, no. 21: 12731. https://doi.org/10.3390/ijms232112731