
Effects of Sub-Minimum Inhibitory Concentrations of Imipenem and Colistin on Expression of Biofilm-Specific Antibiotic Resistance and Virulence Genes in Acinetobacter baumannii Sequence Type 1894
 , , and
, , and
Abstract
:1. Introduction
2. Results
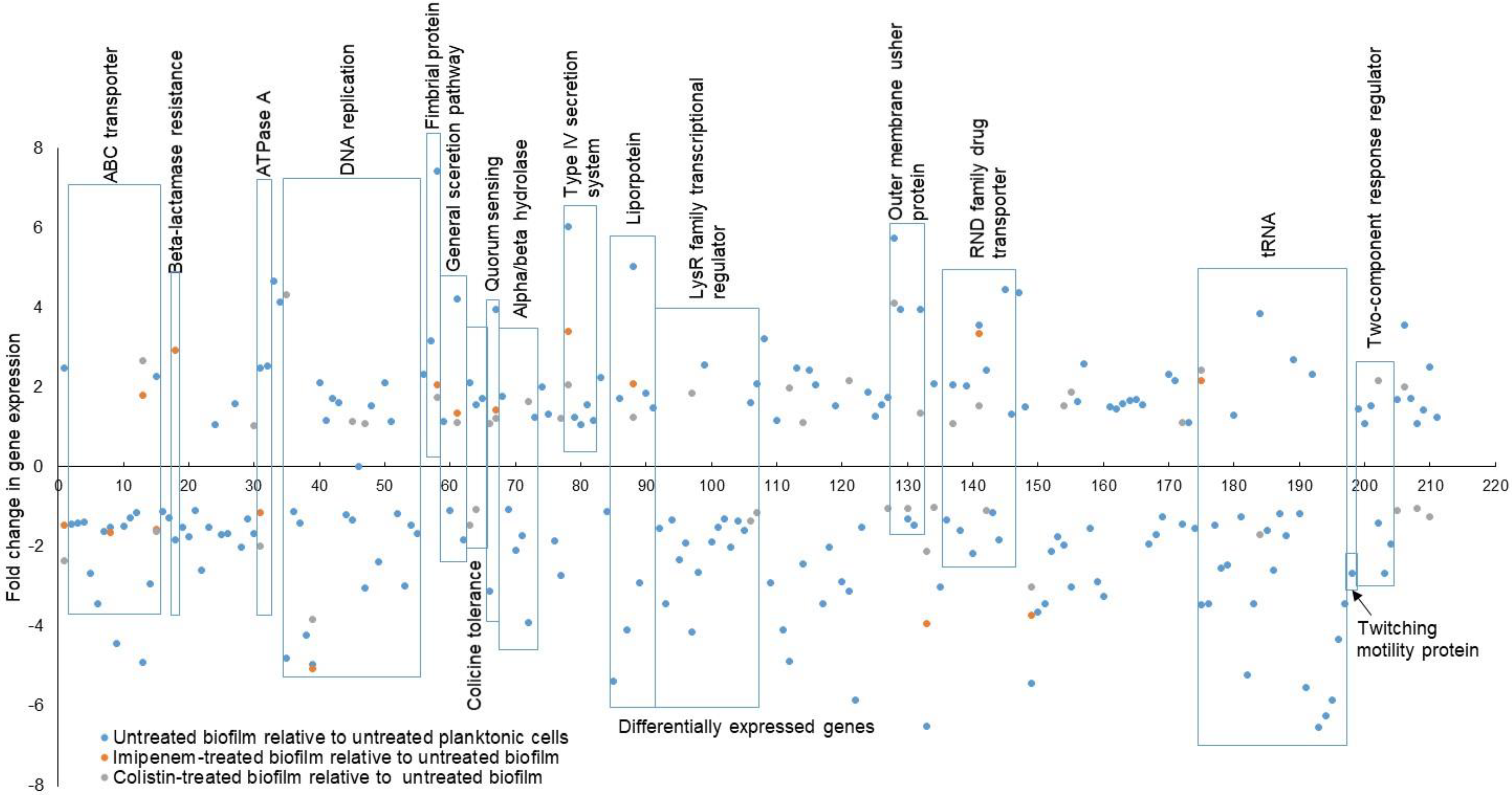
2.1. Gene Expression Profile Associated with Biofilm Formation
2.2. Gene Expression Profile of Biofilm Treated with Sub-MIC of Imipenem
2.3. Gene Expression Profile of Biofilm Treated with Sub-MIC of Colistin
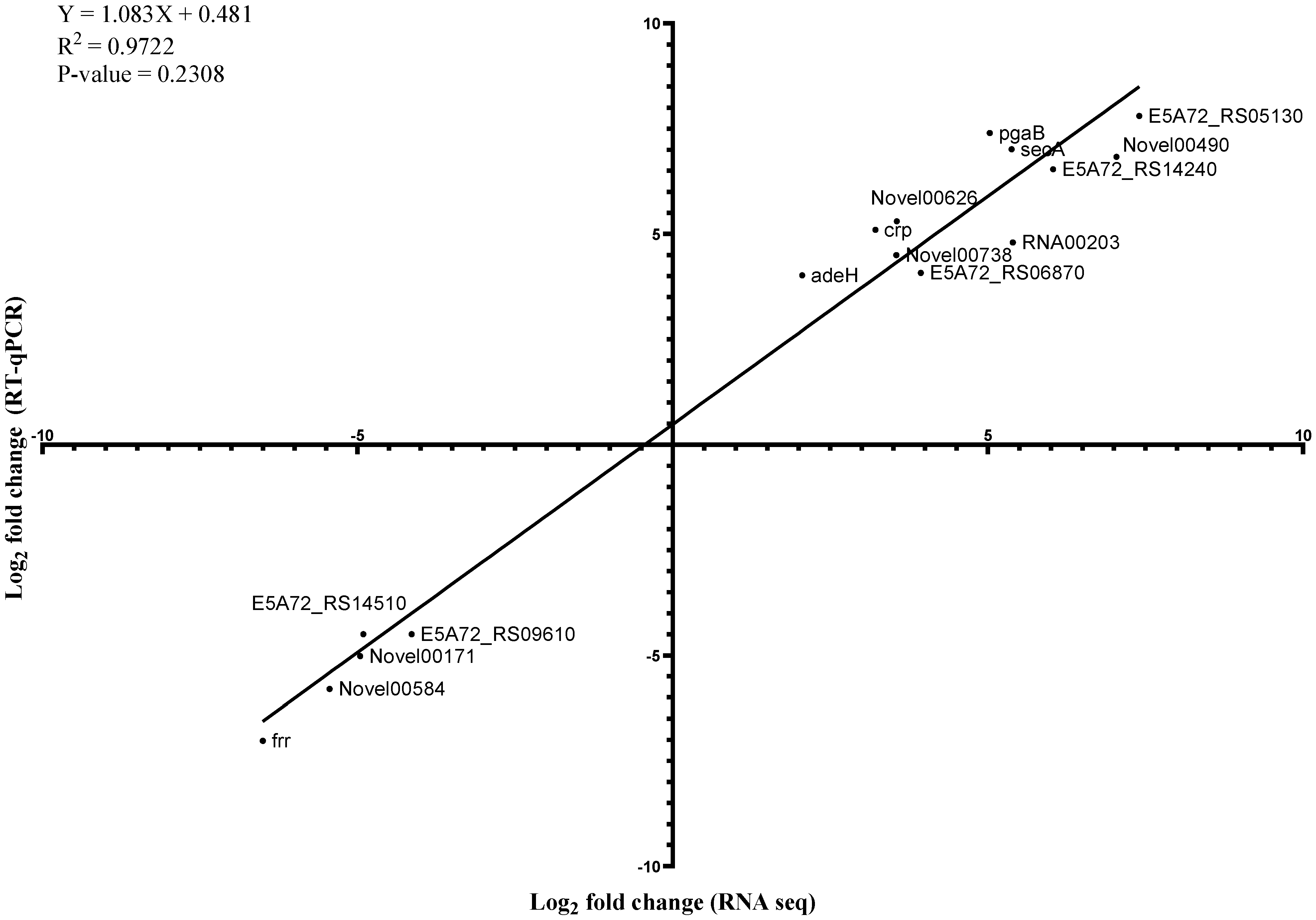
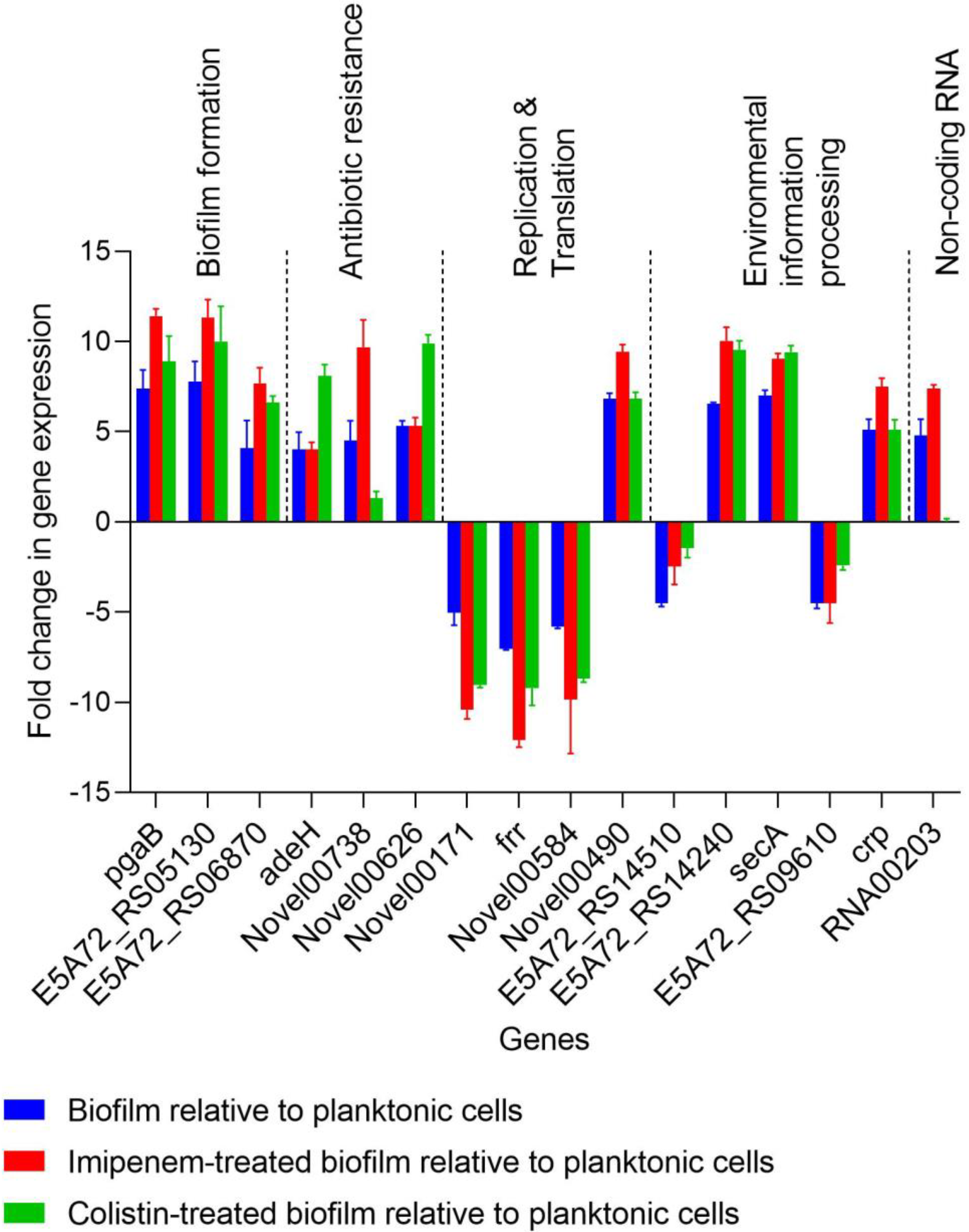
2.4. Verification of Genes Induced or Repressed by the Sub-MICs of Imipenem and Colistin
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain and Culture Conditions
4.2. Biofilm Preparation and Exposure to Antibiotics
4.3. RNA Extraction
4.4. RNA Sequencing
4.5. Bioinformatics Analysis
4.5.1. Novel Gene and Gene Structure Analysis
4.5.2. The GO and KEGG Enrichment Analyses
4.6. Verification of Genes of Induced or Repressed by Exposure to Sub-MICs of Imipenem and Colistin
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gedefie, A.; Demsis, W.; Ashagrie, M.; Kassa, Y.; Tesfaye, M.; Tilahun, M.; Bisetegn, H.; Sahle, Z. Acinetobacter Baumannii Biofilm Formation and Its Role in Disease Pathogenesis: A Review. Infect. Drug Resist. 2021, 14, 3711. [Google Scholar] [CrossRef] [PubMed]
- Espinal, P.; Martí, S.; Vila, J. Effect of Biofilm Formation on the Survival of Acinetobacter Baumannii on Dry Surfaces. J. Hosp. Infect. 2012, 80, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.H.; Su, P.W.; Moi, S.H.; Chuang, L.Y. Biofilm Formation in Acinetobacter Baumannii: Genotype-Phenotype Correlation. Molecules 2019, 24, 1849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harding, C.M.; Hennon, S.W.; Feldman, M.F. Uncovering the Mechanisms of Acinetobacter Baumannii Virulence. Nat. Rev. Microbiol. 2018, 16, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Harding, C.M.; Tracy, E.N.; Carruthers, M.D.; Rather, P.N.; Actis, L.A.; Munson, R.S. Acinetobacter Baumannii Strain M2 Produces Type IV Pili Which Play a Role in Natural Transformation and Twitching Motility but Not Surface-Associated Motility. MBio 2013, 4, e00360-13. [Google Scholar] [CrossRef] [Green Version]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial Biofilms: A Common Cause of Persistent Infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [Green Version]
- Penesyan, A.; Nagy, S.S.; Kjelleberg, S.; Gillings, M.R.; Paulsen, I.T. Rapid Microevolution of Biofilm Cells in Response to Antibiotics. NPJ Biofilms Microbiomes 2019, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Ababneh, Q.; Abu Laila, S.; Jaradat, Z. Prevalence, Genetic Diversity, Antibiotic Resistance and Biofilm Formation of Acinetobacter Baumannii Isolated from Urban Environments. J. Appl. Microbiol. 2022; In press. [Google Scholar]
- Cheng, V.C.C.; Wong, S.C.; Chen, J.H.K.; So, S.Y.C.; Wong, S.C.Y.; Ho, P.L.; Yuen, K.Y. Control of Multidrug-Resistant Acinetobacter Baumannii in Hong Kong: Role of Environmental Surveillance in Communal Areas after a Hospital Outbreak. Am. J. Infect. Control 2018, 46, 60–66. [Google Scholar] [CrossRef]
- Jung, J.Y.; Park, M.S.; Kim, S.E.; Park, B.H.; Son, J.Y.; Kim, E.Y.; Lim, J.E.; Lee, S.K.; Lee, S.H.; Lee, K.J. Risk Factors for Multi-Drug Resistant Acinetobacter Baumannii bacteremia in Patients with Colonization in the Intensive Care Unit. BMC Infect. Dis. 2010, 10, 228. [Google Scholar] [CrossRef] [Green Version]
- El Kheloui Raja, E.M.S.; Asma, L.; Rachida, M.; Fatima, H. Acinetobacter Baumannii Extracellular Matrix as An Antibiofilm and Anti-Infection Target. World J. Pharm. Res. 2022, 11, 10–35. [Google Scholar]
- Shenkutie, A.M.; Yao, M.Z.; Siu, G.K.H.; Wong, B.K.C.; Leung, P.H.M. Biofilm-Induced Antibiotic Resistance in Clinical Acinetobacter Baumannii Isolates. Antibiotics 2020, 9, 817. [Google Scholar] [CrossRef] [PubMed]
- García-Quintanilla, M.; Pulido, M.R.; López-Rojas, R.; Pachón, J.; McConnell, M.J. Emerging Therapies for Multidrug Resistant Acinetobacter Baumannii. Trends Microbiol. 2013, 21, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Paul, M.; Carrara, E.; Retamar, P.; Tängdén, T.; Bitterman, R.; Bonomo, R.A.; De Waele, J.; Daikos, G.L.; Akova, M.; Harbarth, S. European Society of Clinical Microbiology and Infectious Diseases (ESCMID) Guidelines for the Treatment of Infections Caused by Multidrug-Resistant Gram-Negative Bacilli (Endorsed by European Society of Intensive Care Medicine). Clin. Microbiol. Infect. 2022, 28, 521–547. [Google Scholar] [CrossRef] [PubMed]
- Høiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic Resistance of Bacterial Biofilms. Int. J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, J.B. Antibiotic-Induced Biofilm Formation. Int. J. Artif. Organs 2011, 34, 737–751. [Google Scholar] [CrossRef]
- Tarín-Pelló, A.; Suay-García, B.; Pérez-Gracia, M.-T. Antibiotic Resistant Bacteria: Current Situation and Treatment Options to Accelerate the Development of a New Antimicrobial Arsenal. Expert Rev. Anti. Infect. Ther. 2022, 20, 1095–1108. [Google Scholar] [CrossRef]
- Jolivet-Gougeon, A.; Bonnaure-Mallet, M. Biofilms as a Mechanism of Bacterial Resistance. Drug Discov. Today Technol. 2014, 11, 49–56. [Google Scholar] [CrossRef]
- Hathroubi, S.; Mekni, M.A.; Domenico, P.; Nguyen, D.; Jacques, M. Biofilms: Microbial Shelters against Antibiotics. Microb. Drug Resist. 2017, 23, 147–156. [Google Scholar] [CrossRef]
- Choi, A.H.K.; Slamti, L.; Avci, F.Y.; Pier, G.B.; Maira-Litrán, T. The PgaABCD Locus of Acinetobacter Baumannii Encodes the Production of Poly-β-1-6-N-Acetylglucosamine, Which Is Critical for Biofilm Formation. J. Bacteriol. 2009, 191, 5953–5963. [Google Scholar] [CrossRef] [Green Version]
- Law, S.K.K.; Tan, H.S. The Role of Quorum Sensing, Biofilm Formation, and Iron Acquisition as Key Virulence Mechanisms in Acinetobacter Baumannii and the Corresponding Anti-Virulence Strategies. Microbiol. Res. 2022, 260, 127032. [Google Scholar] [CrossRef]
- Vu, B.; Chen, M.; Crawford, R.J.; Ivanova, E.P. Bacterial Extracellular Polysaccharides Involved in Biofilm Formation. Molecules 2009, 14, 2535–2554. [Google Scholar] [CrossRef] [PubMed]
- Mah, T.F. Biofilm-Specific Antibiotic Resistance. Future Microbiol. 2012, 7, 1061–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branda, S.S.; Vik, Å.; Friedman, L.; Kolter, R. Biofilms: The Matrix Revisited. Trends Microbiol. 2005, 13, 20–26. [Google Scholar] [CrossRef]
- Kaushik, V.; Tiwari, M.; Tiwari, V. Interaction of RecA Mediated SOS Response with Bacterial Persistence, Biofilm Formation, and Host Response. Int. J. Biol. Macromol. 2022, 217, 931–943. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, K.K.; Goldmann, D.A.; Pier, G.B. Use of Confocal Microscopy to Analyze the Rate of Vancomycin Penetration through Staphylococcus Aureus Biofilms. Antimicrob. Agents Chemother. 2005, 49, 2467–2473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Ray, P.; Das, A.; Sharma, M. Penetration of Antibiotics through Staphylococcus Aureus and Staphylococcus Epidermidis Biofilms. J. Antimicrob. Chemother. 2010, 65, 1955–1958. [Google Scholar] [CrossRef] [Green Version]
- Ronish, L.A.; Lillehoj, E.; Fields, J.K.; Sundberg, E.J.; Piepenbrink, K.H. The Structure of PilA from Acinetobacter Baumannii AB5075 Suggests a Mechanism for Functional Specialization in Acinetobacter Type IV Pili. J. Biol. Chem. 2019, 294, 218–230. [Google Scholar] [CrossRef] [Green Version]
- Yamabe, K.; Arakawa, Y.; Shoji, M.; Miyamoto, K.; Tsuchiya, T.; Minoura, K.; Akeda, Y.; Tomono, K.; Onda, M. Enhancement of Acinetobacter Baumannii Biofilm Growth by Cephem Antibiotics via Enrichment of Protein and Extracellular DNA in the Biofilm Matrices. J. Appl. Microbiol. 2022, 133, 2002–2013. [Google Scholar] [CrossRef]
- Laureti, L.; Matic, I.; Gutierrez, A. Bacterial Responses and Genome Instability Induced by Subinhibitory Concentrations of Antibiotics. Antibiotics 2013, 2, 100–114. [Google Scholar] [CrossRef] [Green Version]
- Baharoglu, Z.; Garriss, G.; Mazel, D. Multiple Pathways of Genome Plasticity Leading to Development of Antibiotic Resistance. Antibiotics 2013, 2, 288–315. [Google Scholar] [CrossRef]
- Tomaras, A.P.; Dorsey, C.W.; Edelmann, R.E.; Actis, L.A. Attachment to and Biofilm Formation on Abiotic Surfaces by Acinetobacter Baumannii: Involvement of a Novel Chaperone-Usher Pili Assembly System. Microbiology 2003, 149, 3473–3484. [Google Scholar] [CrossRef] [PubMed]
- Peleg, A.Y.; de Breij, A.; Adams, M.D.; Cerqueira, G.M.; Mocali, S.; Galardini, M.; Nibbering, P.H.; Earl, A.M.; Ward, D.V.; Paterson, D.L.; et al. The Success of Acinetobacter Species; Genetic, Metabolic and Virulence Attributes. PLoS ONE 2012, 7, e46984. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.S.; Rezende, I.M.; Luehring, T.A.F.M.; Vieira, C.D.; Drumond, B.P.; de Macêdo Farias, L.; Nobre, V.; Gonçalves, R.; dos Santos, S.G. Sub-Inhibitory Concentrations of Polymyxin B Modulates Pathogenicity Factors and Transcriptional Regulators Genes in Multi-Resistant Acinetobacter Baumannii Strains. Braz. J. Dev. 2021, 7, 8463–8480. [Google Scholar] [CrossRef]
- Antunes, L.C.M.; Ferreira, R.B.R.; Buckner, M.M.C.; Finlay, B.B. Quorum Sensing in Bacterial Virulence. Microbiology 2010, 156, 2271–2282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breijyeh, Z.; Jubeh, B.; Karaman, R. Resistance of Gram-Negative Bacteria to Current Antibacterial Agents and Approaches to Resolve It. Molecules 2020, 25, 1340. [Google Scholar] [CrossRef] [Green Version]
- Raju, D.V.; Nagarajan, A.; Pandit, S.; Nag, M.; Lahiri, D.; Upadhye, V. Effect of Bacterial Quorum Sensing and Mechanism of Antimicrobial Resistance. Biocatal. Agric. Biotechnol. 2022, 43, 102409. [Google Scholar] [CrossRef]
- Papenfort, K.; Bassler, B.L. Quorum Sensing Signal–Response Systems in Gram-Negative Bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.-H.; Tian, X. Quorum Sensing and Bacterial Social Interactions in Biofilms. Sensors 2012, 12, 2519–2538. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; Coppola, R. Quorum Sensing and Phytochemicals. Int. J. Mol. Sci. 2013, 14, 12607–12619. [Google Scholar] [CrossRef] [Green Version]
- Passos da Silva, D.; Schofield, M.C.; Parsek, M.R.; Tseng, B.S. An Update on the Sociomicrobiology of Quorum Sensing in Gram-Negative Biofilm Development. Pathogens 2017, 6, 51. [Google Scholar] [CrossRef]
- Tang, K.; Zhang, X.-H. Quorum Quenching Agents: Resources for Antivirulence Therapy. Mar. Drugs 2014, 12, 3245–3282. [Google Scholar] [CrossRef] [PubMed]
- Brelles-Mario, G.; Bedmar, E.J. Detection, Purification and Characterisation of Quorum-Sensing Signal Molecules in Plant-Associated Bacteria. In Proceedings of the J. Biotechnol. 2001, 91, 197–209. [Google Scholar] [CrossRef]
- Pompilio, A.; Scribano, D.; Sarshar, M.; Di Bonaventura, G.; Palamara, A.T.; Ambrosi, C. Gram-Negative Bacteria Holding Together in a Biofilm: The Acinetobacter Baumannii Way. Microorganisms 2021, 9, 1353. [Google Scholar] [CrossRef] [PubMed]
- Kröger, C.; Kary, S.C.; Schauer, K.; Cameron, A.D.S. Genetic Regulation of Virulence and Antibiotic Resistance in Acinetobacter Baumannii. Genes 2016, 8, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayoub Moubareck, C.; Hammoudi Halat, D. Insights into Acinetobacter Baumannii: A Review of Microbiological, Virulence, and Resistance Traits in a Threatening Nosocomial Pathogen. Antibiotics 2020, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- Rao, R.; Karthika, R.; Singh, S.; Shashikala, P.; Kanungo, R.; Jayachandran, S.; Prashanth, K. Correlation between Biofilm Production and Multiple Drug Resistance in Imipenem Resistant Clinical Isolates of Acinetobacter Baumannii. Indian J. Med. Microbiol. 2008, 26, 333–337. [Google Scholar] [CrossRef]
- Poole, K. Efflux Pumps as Antimicrobial Resistance Mechanisms. Ann. Med. 2007, 39, 162–176. [Google Scholar] [CrossRef]
- Subhadra, B.; Kim, D.H.; Woo, K.; Surendran, S.; Choi, C.H. Control of Biofilm Formation in Healthcare: Recent Advances Exploiting Quorum-Sensing Interference Strategies and Multidrug Efflux Pump Inhibitors. Materials 2018, 11, 1676. [Google Scholar] [CrossRef] [Green Version]
- Poole, K.; Krebes, K.; McNally, C.; Neshat, S. Multiple Antibiotic Resistance in Pseudomonas Aeruginosa: Evidence for Involvement of an Efflux Operon. J. Bacteriol. 1993, 175, 7363–7372. [Google Scholar] [CrossRef] [Green Version]
- Vaillancourt, M.; Limsuwannarot, S.P.; Bresee, C.; Poopalarajah, R.; Jorth, P. Pseudomonas Aeruginosa MexR and MexEF Antibiotic Efflux Pump Variants Exhibit Increased Virulence. Antibiotics 2021, 10, 1164. [Google Scholar] [CrossRef]
- Impey, R.E.; Hawkins, D.A.; Sutton, J.M.; Soares da Costa, T.P. Overcoming Intrinsic and Acquired Resistance Mechanisms Associated with the Cell Wall of Gram-Negative Bacteria. Antibiotics 2020, 9, 623. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Shi, Q.-S.; Huang, X.-M.; Xie, X.-B. The Three Bacterial Lines of Defense against Antimicrobial Agents. Int. J. Mol. Sci. 2015, 16, 21711–21733. [Google Scholar] [CrossRef] [Green Version]
- Scoffone, V.C.; Trespidi, G.; Barbieri, G.; Irudal, S.; Perrin, E.; Buroni, S. Role of RND Efflux Pumps in Drug Resistance of Cystic Fibrosis Pathogens. Antibiotics 2021, 10, 863. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Lu, F.; Yuan, F.; Jiang, D.; Zhao, P.; Zhu, J.; Cheng, H.; Cao, J.; Lu, G. Biofilm Formation Caused by Clinical Acinetobacter Baumannii Isolates Is Associated with Overexpression of the AdeFGH Efflux Pump. Antimicrob. Agents Chemother. 2015, 59, 4817–4825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vrancianu, C.O.; Gheorghe, I.; Czobor, I.B.; Chifiriuc, M.C. Antibiotic Resistance Profiles, Molecular Mechanisms and Innovative Treatment Strategies of Acinetobacter Baumannii. Microorganisms 2020, 8, 935. [Google Scholar] [CrossRef]
- Aurilio, C.; Sansone, P.; Barbarisi, M.; Pota, V.; Giaccari, L.G.; Coppolino, F.; Barbarisi, A.; Passavanti, M.B.; Pace, M.C. Mechanisms of Action of Carbapenem Resistance. Antibiotics 2022, 11, 421. [Google Scholar] [CrossRef]
- Abel Zur Wiesch, P.; Abel, S.; Gkotzis, S.; Ocampo, P.; Engelstädter, J.; Hinkley, T.; Magnus, C.; Waldor, M.K.; Udekwu, K.; Cohen, T. Classic Reaction Kinetics Can Explain Complex Patterns of Antibiotic Action. Sci. Transl. Med. 2015, 7, 287ra73. [Google Scholar] [CrossRef] [Green Version]
- Pu, Y.; Ke, Y.; Bai, F. Active Efflux in Dormant Bacterial Cells–New Insights into Antibiotic Persistence. Drug Resist. Updat. 2017, 30, 7–14. [Google Scholar] [CrossRef]
- Pu, Y.; Zhao, Z.; Li, Y.; Zou, J.; Ma, Q.; Zhao, Y.; Ke, Y.; Zhu, Y.; Chen, H.; Baker, M.A.B.; et al. Enhanced Efflux Activity Facilitates Drug Tolerance in Dormant Bacterial Cells. Mol. Cell 2016, 62, 284–294. [Google Scholar] [CrossRef] [Green Version]
- Lewis, K. Riddle of Biofilm Resistance. Antimicrob. Agents Chemother. 2001, 45, 999–1007. [Google Scholar] [CrossRef] [Green Version]
- Alav, I.; Sutton, J.M.; Rahman, K.M. Role of Bacterial Efflux Pumps in Biofilm Formation. J. Antimicrob. Chemother. 2018, 73, 2003–2020. [Google Scholar] [CrossRef] [PubMed]
- Boll, J.M.; Tucker, A.T.; Klein, D.R.; Beltran, A.M.; Brodbelt, J.S.; Davies, B.W.; Trent, M.S. Reinforcing Lipid a Acylation on the Cell Surface of Acinetobacter Baumannii Promotes Cationic Antimicrobial Peptide Resistance and Desiccation Survival. MBio 2015, 6, e00478-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mengiste, B.; Lulie, S.; Getachew, B.; Gebrelibanos, M.; Mekuria, A.; Masresha, B. In-vitro Antibacterial Activity of Extracts from Aerial Parts of Verbena Officinalis. Adv. Biol. Res. 2015, 9, 53–57. [Google Scholar]
- Guha, S.; Udupa, S.; Ahmed, W.; Nagaraja, V. Rewired Downregulation of DNA Gyrase Impacts Cell Division, Expression of Topology Modulators, and Transcription in Mycobacterium Smegmatis. J. Mol. Biol. 2018, 430, 4986–5001. [Google Scholar] [CrossRef] [PubMed]
- Willmott, C.J.R.; Critchlow, S.E.; Eperon, I.C.; Maxwell, A. The Complex of DNA Gyrase and Quinolone Drugs with DNA Forms a Barrier to Transcription by RNA Polymerase. J. Mol. Biol. 1994, 242, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Rovinskiy, N.; Agbleke, A.A.; Chesnokova, O.; Pang, Z.; Higgins, N.P. Rates of Gyrase Supercoiling and Transcription Elongation Control Supercoil Density in a Bacterial Chromosome. PLOS Genet. 2012, 8, e1002845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiel, M.C.; Kaji, H.; Kaji, A. Ribosome Recycling: An Essential Process of Protein Synthesis. Biochem. Mol. Biol. Educ. 2007, 35, 40–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlov, M.Y.; Freistroffer, D.V.; MacDougall, J.; Buckingham, R.H.; Ehrenberg, M. Fast Recycling of Escherichia Coli Ribosomes Requires Both Ribosome Recycling Factor (RRF) and Release Factor RF3. EMBO J. 1997, 16, 4134–4141. [Google Scholar] [CrossRef]
- Nwugo, C.C.; Gaddy, J.A.; Zimbler, D.L.; Actis, L.A. Deciphering the Iron Response in Acinetobacter Baumannii: A Proteomics Approach. J. Proteomics 2011, 74, 44–58. [Google Scholar] [CrossRef] [Green Version]
- Storz, G.; Vogel, J.; Wassarman, K.M. Regulation by Small RNAs in Bacteria: Expanding Frontiers. Mol. Cell 2011, 43, 880–891. [Google Scholar] [CrossRef] [Green Version]
- Papenfort, K.; Vogel, J. Multiple Target Regulation by Small Noncoding RNAs Rewires Gene Expression at the Post-Transcriptional Level. Res. Microbiol. 2009, 160, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Roncarati, D.; Scarlato, V.; Vannini, A. Targeting of Regulators as a Promising Approach in the Search for Novel Antimicrobial Agents. Microorganisms 2022, 10, 185. [Google Scholar] [CrossRef] [PubMed]
- Bak, G.; Lee, J.Y.J.; Suk, S.; Kim, D.; Lee, J.Y.J.; Kim, K.S.; Choi, B.S.; Lee, Y. Identification of Novel sRNAs Involved in Biofilm Formation, Motility, and Fimbriae Formation in Escherichia Coli. Sci. Rep. 2015, 5, 15287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, R.P.; Mukherjee, R.; Chang, C.-M. Emerging Concern with Imminent Therapeutic Strategies for Treating Resistance in Biofilm. Antibiotics 2022, 11, 476. [Google Scholar] [CrossRef]
- Di Bonaventura, G.; Spedicato, I.; D’Antonio, D.; Robuffo, I.; Piccolomini, R. Biofilm Formation by Stenotrophomonas Maltophilia: Modulation by Quinolones, Trimethoprim-Sulfamethoxazole, and Ceftazidime. Antimicrob. Agents Chemother. 2004, 48, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript Assembly and Quantification by RNA-Seq Reveals Unannotated Transcripts and Isoform Switching during Cell Differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and Memory-Efficient Alignment of Short DNA Sequences to the Human Genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.H.; Yu, H.; Tu, K.; Li, C.; Li, Y.X.; Li, Y.Y. DCGL: An R Package for Identifying Differentially Coexpressed Genes and Links from Gene Expression Microarray Data. Bioinformatics 2010, 26, 2637–2638. [Google Scholar] [CrossRef] [Green Version]
- McClure, R.; Balasubramanian, D.; Sun, Y.; Bobrovskyy, M.; Sumby, P.; Genco, C.A.; Vanderpool, C.K.; Tjaden, B. Computational Analysis of Bacterial RNA-Seq Data. Nucleic Acids Res. 2013, 41, e140. [Google Scholar] [CrossRef] [Green Version]
- Bharti, R.; Siebert, D.; Blombach, B.; Grimm, D.G. Systematic Analysis of the Underlying Genomic Architecture for Transcriptional–Translational Coupling in Prokaryotes. NAR Genom. Bioinform. 2022, 4, lqac074. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Group | Transcribed Genes | Differentially Expressed | |||
|---|---|---|---|---|---|
| Total No. (%) | Novel Genes | Genes Encoding Hypothetical Proteins | Small RNA | ||
| Biofilm vs. planktonic cells | 3075 | 1592 (51.8) | 96 | 431 | 5 |
| Imipenem-treated biofilm vs. untreated biofilm cells | 2885 | 106 (3.7) | 7 | 11 | 1 |
| Colistin-treated biofilm vs. untreated biofilm cells | 2912 | 368 (12.6) | 33 | 74 | 0 |
| Gene_id | KEGG_ID | Fold Change in Gene Expression | Function/Product | ||
|---|---|---|---|---|---|
| Untreated Biofilm Relative to Untreated Planktonic | Imipenem-Treated Biofilm Relative to Untreated Biofilm | Colistin-Treated Biofilm Relative to Untreated Biofilm | |||
| Lipoprotein | |||||
| E5A72_RS12100 | acb: A1S_0938 | 5.0363 | 2.09 | 1.23 | Lipoprotein (biofilm matrix) |
| ATP-binding cassette (ABC) transporter | |||||
| E5A72_RS02170 | acb: A1S_2611 | 2.3183 | 2.14368 | 1.02 | Transport protein of outer membrane lipoproteins |
| E5A72_RS16335 | acb: A1S_1722 | −1.5054 | −1.639 | 0.095 | ABC transporter ATP-binding protein |
| E5A72_RS14510 | acb: A1S_1359 | −4.9058 | 1.7825 | 2.6702 | ABC-type Fe3+ transport system |
| E5A72_RS08785 | acb: A1S_0229 | 2.0395 | 1.5784 | −1.6349 | Lipopolysaccharide export system permease protein |
| E5A72_RS15155 | acb: A1S_1482 | −3.3293 | 1.0455 | 2.0257 | D-and L-methionine transport protein |
| β-lactam resistance | |||||
| E5A72_RS00940 | acb: A1S_2367 | −1.8362 | 2.92 | 1.16 * | ampC; β-lactamase |
| Novel00738 | acb: A1S_2736 | 3.5549 | 3.3532 | −1.013 | Resistance–nodulation–cell division (RND) family drug transporter |
| DNA replication and repair | |||||
| Novel00171 | acb: A1S_2626 | −4.9582 | −5.0584 | −3.8428 | DNA gyrase |
| E5A72_RS14680 | acb: A1S_1389 | −1.3449 | −0.0409 * | 1.1446 | DNA polymerase V component |
| E5A72_RS03815 | acb: A1S_2918 | −3.0414 | −0.08 * | 1.0746 | DNA repair protein |
| Fimbrial protein and csu operon | |||||
| E5A72_RS05130 | acb: A1S_3177 | 7.4033 | 2.0455 | 1.7527 | Fimbrial protein |
| E5A72_RS19200 | acb: A1S_2217 | 5.7238 | NA | 2.1677 | Protein CsuA |
| E5A72_RS19190 | acb: A1S_2215 | 1.3111 | −0.683 * | 1.0371 | Protein CsuC |
| E5A72_RS19180 | acb: A1S_2213 | 3.9517 | 0.2679 * | 4.1059 | Protein CsuE |
| Bacterial secretion system | |||||
| E5A72_RS14240 | acb: A1S_1310 | 6.0356 | 3.387 | 2.045 | Type VI secretion system protein |
| E5A72_RS15605 | acb: A1S_1564 | 4.2 | 1.3587 | 1.1006 | General secretion pathway protein J |
| E5A72_RS03500 | acb: A1S_2862 | 5.376 | 1.65783 | 2.349 | Preprotein translocase subunit SecA |
| Two-component regulatory system | |||||
| E5A72_RS09610 | acb: A1S_0399 | −4.1397 | −0.346 * | 1.8361 | LysR family transcriptional regulator |
| E5A72_RS12380 | acb: A1S_0992 | 1.617 | −0.132 * | −1.3517 | |
| E5A72_RS13515 | acb: A1S_1182 | 3.216 | 2.06 | 0.065 * | cAMP-activated global transcriptional regulator CRP |
| Novel00822 | acb: A1S_0685 | −1.4033 | 1.788 * | 2.4156 | Two-component response regulator |
| Quorum sensing or quenching | |||||
| E5A72_RS06870 | acb: A1S_0109 | 3.9355 | 1.43253 | 1.21081 | Acyl homoserine lactone (AHL) synthase (AbaI) |
| E5A72_RS16835 | acb: A1S_1809 | −3.9015 | 0.267 * | 1.6271 | Hydrolase transmembrane protein |
| Multidrug efflux pump | |||||
| E5A72_RS00640 | acb: A1S_2306 | 2.0527 | −0.083 * | 2.1244 | RND efflux transporter |
| E5A72_RS11940 | acb: A1S_0908 | 2.4322 | −0.4375 | 1.0937 | RND family multidrug resistance secretion protein |
| Transcription and translation | |||||
| E5A72_RS17735 | acb: A1S_1974 | −6.5036 | −3.9244 | −2.1244 | Ribosome releasing factor |
| Novel00584 | acb: A1S_1617 | −5.4437 | −3.7365 | −3.01 | 30S ribosomal protein S20 |
| Novel00490 | - | 7.0401 | 1.1587 | 0.415 | Transcription termination factor Rho OS |
| E5A72_RS04400 | acb: A1S_3029 | −3.4737 | 2.161 | 2.4156 | tRNA-Arg |
| Peptidoglycan biosynthesis | |||||
| E5A72_RS10790 | acb: A1S_1965 | 1.6823 | −0.1303 * | −1.6917 | UDP-N-acetylglucosamine 1-carboxyvinyltransferase |
| Novel00626 | acb: A1S_1965 | 3.5549 | NA | 2.1677 | UDP-N-acetylglucosamine acyltransferase |
| E5A72_RS05230 | acb: A1S_3203 | 1.0844 | −0217* | −1.032 | UDP-N-acetylmuramoylalanyl-D-glutamate-2, 6-diaminopimelate ligase |
| E5A72_RS17730 | acb: A1S_1973 | 2.513 | −0.0783 * | −1.2536 | Undecaprenyl pyrophosphate synthetase |
| E5A72_RS07480 | acb: A1S_2968 | −2.7334 | 0.4605 * | 1.2183 | Hypothetical protein |
| E5A72_RS05220 | acb: A1S_3200 | 1.7367 | 0.0003 * | −1.0371 | Phospho-N-acetylmuramoyl-pentapeptide transferase |
| Outer membrane protein | |||||
| E5A72_RS11985 | - | −4.8714 | −0.1244 * | 1.9751 | OprD family outer membrane porin |
| E5A72_RS18480 | acb: A1S_2076 | −2.4293 | 0.4473 * | 1.1074 | Outer membrane porin receptor for Fe (III)- coprogen |
| E5A72_RS01795 | acb: A1S_2538 | −3.1257 | 1.4605 * | 2.1677 | Outer membrane protein CarO precursor |
| Transcriptional regulators and others | |||||
| E5A72_RS13590 | acb: A1S_1197 | −2.5865 | −1.2664 | 0.240 * | Extracellular nuclease |
| E5A72_RS03015 | acb: A1S_2767 | −1.6789 | 0.7044 | 1.0387 | AraC family transcriptional regulator |
| E5A72_RS04330 | acb: A1S_3229 | 3.8412 | 0.25977 | −1.6917 | tRNA-i(6)A37 modification enzyme |
| E5A72_RS01785 | acb: A1S_2536 | 2.4782 | −1.1416 | −1.9789 | ATPase |
| E5A72_RS14655 | acb: A1S_1386 | −4.8062 | NA | 4.3282 | Catalase; K03781 catalase |
| E5A72_RS01830 | acb: A1S_2546 | −1.9701 | 0.9841 | 1.5253 | Secreted trypsin-like serine protease |
| E5A72_RS05375 | acb: A1S_3227 | 2.0827 | −0.02825 | −1.013 | RNA binding protein |
| E5A72_RS16050 | acb: A1S_1670 | −3.0262 | 0.598 * | −3.01 | Secretion protein HlyD |
| E5A72_RS02070 | acb: A1S_2592 | 1.5561 | 0.0036 | −1.0789 | Group A colicins tolerance protein |
| E5A72_RS02065 | acb: A1S_2591 | 2.0955 | −0.0461 | −1.4623 | Group A colicins tolerance protein |
| Non-coding RNA | |||||
| Gene encoding sRNA00203 | - | 5.3951 | 2.38935 | −0.389 * | |
| mRNA_ID | KEGG_ID | KEGG Annotation | Strand | Start | End | Length (bp) | Pathway |
|---|---|---|---|---|---|---|---|
| E5A72_RS12100 | acb: A1S_0938 | PgaB | + | 2520896 | 2522890 | 1995 | Biofilm matrix |
| E5A72_RS05130 | acb: A1S_3177 | Fimbrial protein | − | 1078917 | 1079381 | 2538 | Two-component system |
| E5A72_RS06870 | acb: A1S_0109 | Homoserine lactone synthase | + | 1427610 | 1428176 | 567 | Quorum sensing |
| E5A72_RS00640 | acb: A1S_2306 | RND2 efflux transporter | + | 116590 | 118041 | 555 | Multidrug efflux system |
| Novel00738 | acb: A1S_2736 | RND family drug transporter | + | 617454 | 623989 | 6536 | β-lactam resistance |
| Novel00626 | acb: A1S_1965 | UDP-N acetylglucosamine acyltransferase | − | 3676375 | 3677683 | 1309 | Cationic antimicrobial peptide resistance |
| Novel00171 | acb: A1S_2626 | DNA gyrase | − | 483250 | 484821 | 1572 | DNA replication |
| E5A72_RS17735 | acb: A1S_1974 | Ribosome releasing factor | − | 3685962 | 3686516 | 555 | Translation |
| Novel00584 | acb: A1S_1617 | 30S ribosomal protein S20 | − | 3284582 | 3284881 | 300 | |
| Novel00490 | - | Transcription termination factor Rho OS | − | 2214580 | 2215974 | 1395 | Transcription |
| E5A72_RS14510 | acb: A1S_1359 | ABC3-type Fe3+ transport system | + | 3025424 | 3026461 | 1038 | ABC transporters |
| E5A72_RS14240 | acb: A1S_1310 | K11892 type VI secretion system protein ImpK | + | 2971603 | 2972409 | 807 | Bacterial secretion system |
| E5A72_RS03500 | acb: A1S_2862 | Preprotein translocase subunit SecA | − | 756066 | 758789 | 2724 | |
| E5A72_RS09610 | acb: A1S_0399 | LysR family transcriptional regulator | + | 1993774 | 1994670 | 897 | Two-component system |
| E5A72_RS13515 | acb: A1S_1182 | CRP4 transcriptional regulator | − | 2818333 | 2819040 | 708 | |
| sRNA00203 | - | - | − | 1245743 | 1245795 | 53 | Non-coding RNA |
| Gene ID | Amplicon Size (bp) | Primer/Probe | Sequence (5′ to 3′) |
|---|---|---|---|
| E5A72_RS12100 (pgaB) | 105 | E5A72_RS12100-F | CGGATGCGAATGGTTCTGC |
| E5A72_RS12100-R | GCGTACGGGTTTGAATTTGC | ||
| E5A72_RS05130 | 217 | E5A72_RS05130_4_F | CCGAAGGTACAGCTAACAGTG |
| E5A72_RS05130_4_R | CCACCCACATTTGCATTTACT | ||
| E5A72_RS06870 | 121 | E5A72_RS06870_F | GCCAGACTACTACCCACCAC |
| E5A72_RS06870_R | CTACGGCTGAAAACCTTGAT | ||
| E5A72_RS00640 | 108 | E5A72_RS00640_F | TCAGGCTTCACGTGCACTAC |
| E5A72_RS00640_R | AAACCGAGTGAAGCTGGAGA | ||
| Novel00738 | 79 | Novel00738_F | GCTGCCATTACTCGTTTACCT |
| Novel00738_R | CAGGACGGCTCTCAACAAC | ||
| Novel00738_IN | FAM-GGCAAGCTGTAGCGATGCTTGTTAAT-TAMRA | ||
| Novel00626 | 110 | Novel00626_F | CGCATCGTTACCCATTCTT |
| Novel00626_R | GAAATGCCCTTGTAGGAACTCT | ||
| Novel00626_IN | FAM-TTGGTTGATCGTGTGACTGAAGTTACTGA-TAMRA | ||
| Novel00171 | 109 | Novel00171_F | CATTGCCGGATGTGAGAG |
| Novel00171_R | ACACGAGCAGATTTCTTGTAGG | ||
| E5A72_RS17735 (Frr) | 98 | E5A72_RS17735_F | GCGAAAGTTGCTATCCGTAA |
| E5A72_RS17735_R | GCACGACGCTCATCATCT | ||
| Novel00584 | 114 | Novel00584_F | TGCGTTCTATGGTTCGTACTT |
| Novel00584_R | GCACGACGCTCATCATCT | ||
| Novel00490 | 109 | Novel00490_F | TTAGCCCGTGCATACAACAC |
| Novel00490_R | TAGCCCGTGCATACAACAC | ||
| Novel00490_IN | FAM-TGGTGTGGATGCACATGCTTTAGAAC-TAMRA | ||
| E5A72_RS14510 | 103 | E5A72_RS14510_F | AGGTTTAGGCTGGGAAATGG |
| E5A72_RS14510_R | ATTTGCTGCTTTGCTTACCG | ||
| E5A72_RS14240 | 110 | E5A72_RS14240_1_F | GCACGAGTAGGCGATGAA |
| E5A72_RS14240_1_R | AAAGGTAGCTCACGATGGATAA | ||
| E5A72_RS03500 (secA) | 107 | E5A72_RS03500_F | GACATTATTGCTCAGGCAGGT |
| E5A72_RS03500_R | GCAAGTTTCGCTTTCCAGTT | ||
| E5A72_RS09610 | 86 | E5A72_RS09610_3_F | AAGGTGGAACTGTGATGATGG |
| E5A72_RS09610_3_R | AATTCCCAAACCTGCACAAG | ||
| E5A72_RS13515 | 118 | E5A72_RS13515_3_F | ATCGACCTATCTTCACAACCAG |
| E5A72_RS13515_3_R | ATACACGGCCAACCATTTC | ||
| sRNA00203 | 76 | sRNA00203_4_F | GCATAAAAACCTCTTGAAACTGTTC |
| sRNA00203_4_R | AGCGTTCATTTCAACCGATA | ||
| sRNA00203_4_IN | TCAAGTTCCTTATGATCTCTTCCTTGA | ||
| gyrB | 93 | gyrB-F | ACGATTTACCGTGCTGGTC |
| gyrB-R | GGTATTATCCGTTTCACCAATC | ||
| gyrB_IN | FAM-TATCATCATGGTGATCCGCAATATCC-TAMRA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shenkutie, A.M.; Zhang, J.; Yao, M.; Asrat, D.; Chow, F.W.N.; Leung, P.H.M. Effects of Sub-Minimum Inhibitory Concentrations of Imipenem and Colistin on Expression of Biofilm-Specific Antibiotic Resistance and Virulence Genes in Acinetobacter baumannii Sequence Type 1894. Int. J. Mol. Sci. 2022, 23, 12705. https://doi.org/10.3390/ijms232012705
Shenkutie AM, Zhang J, Yao M, Asrat D, Chow FWN, Leung PHM. Effects of Sub-Minimum Inhibitory Concentrations of Imipenem and Colistin on Expression of Biofilm-Specific Antibiotic Resistance and Virulence Genes in Acinetobacter baumannii Sequence Type 1894. International Journal of Molecular Sciences. 2022; 23(20):12705. https://doi.org/10.3390/ijms232012705
Chicago/Turabian StyleShenkutie, Abebe Mekuria, Jiaying Zhang, Mianzhi Yao, Daniel Asrat, Franklin W. N. Chow, and Polly H. M. Leung. 2022. "Effects of Sub-Minimum Inhibitory Concentrations of Imipenem and Colistin on Expression of Biofilm-Specific Antibiotic Resistance and Virulence Genes in Acinetobacter baumannii Sequence Type 1894" International Journal of Molecular Sciences 23, no. 20: 12705. https://doi.org/10.3390/ijms232012705