
The Transcriptomic Landscape of Pediatric Astrocytoma
 , , , , , ,
, , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
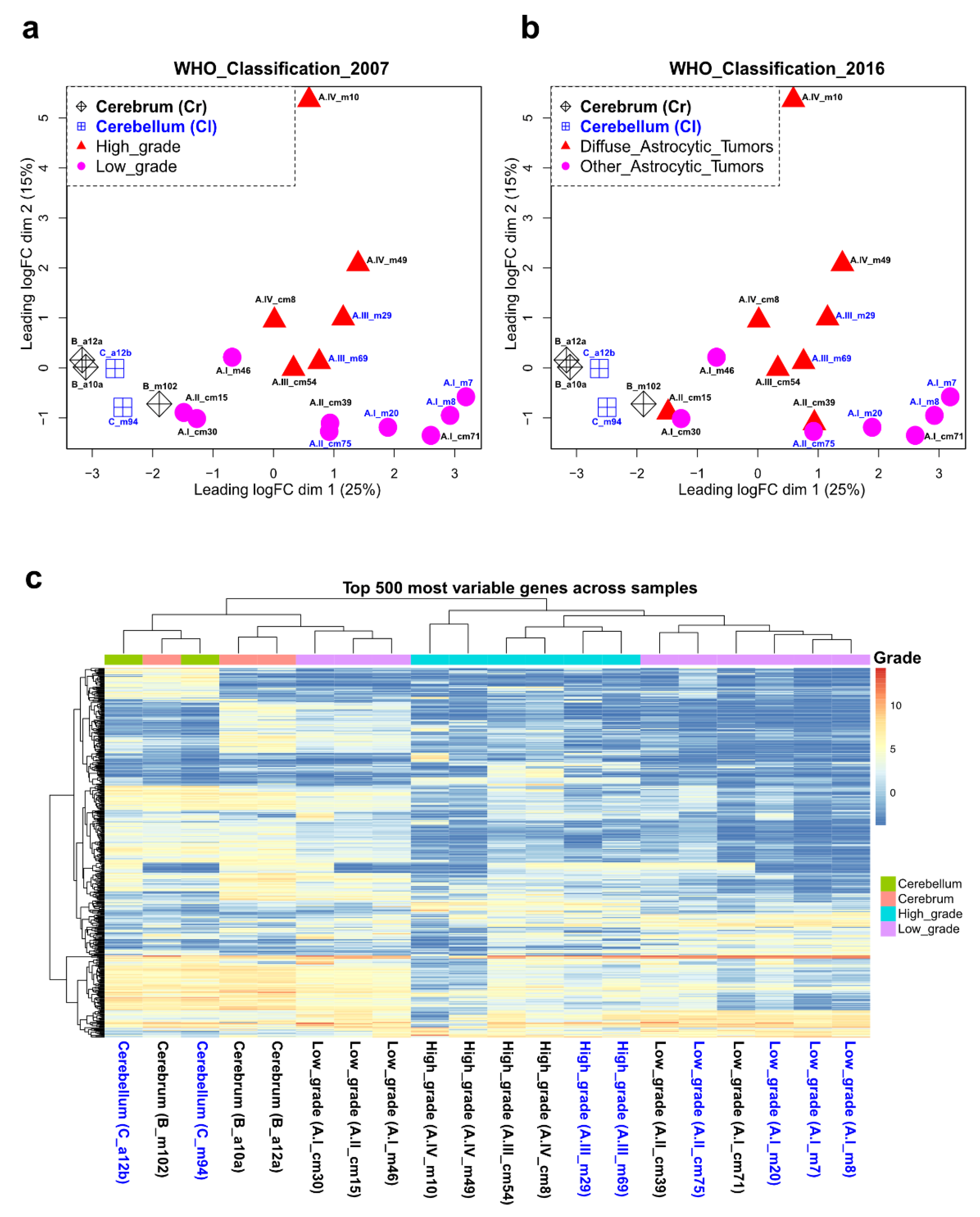
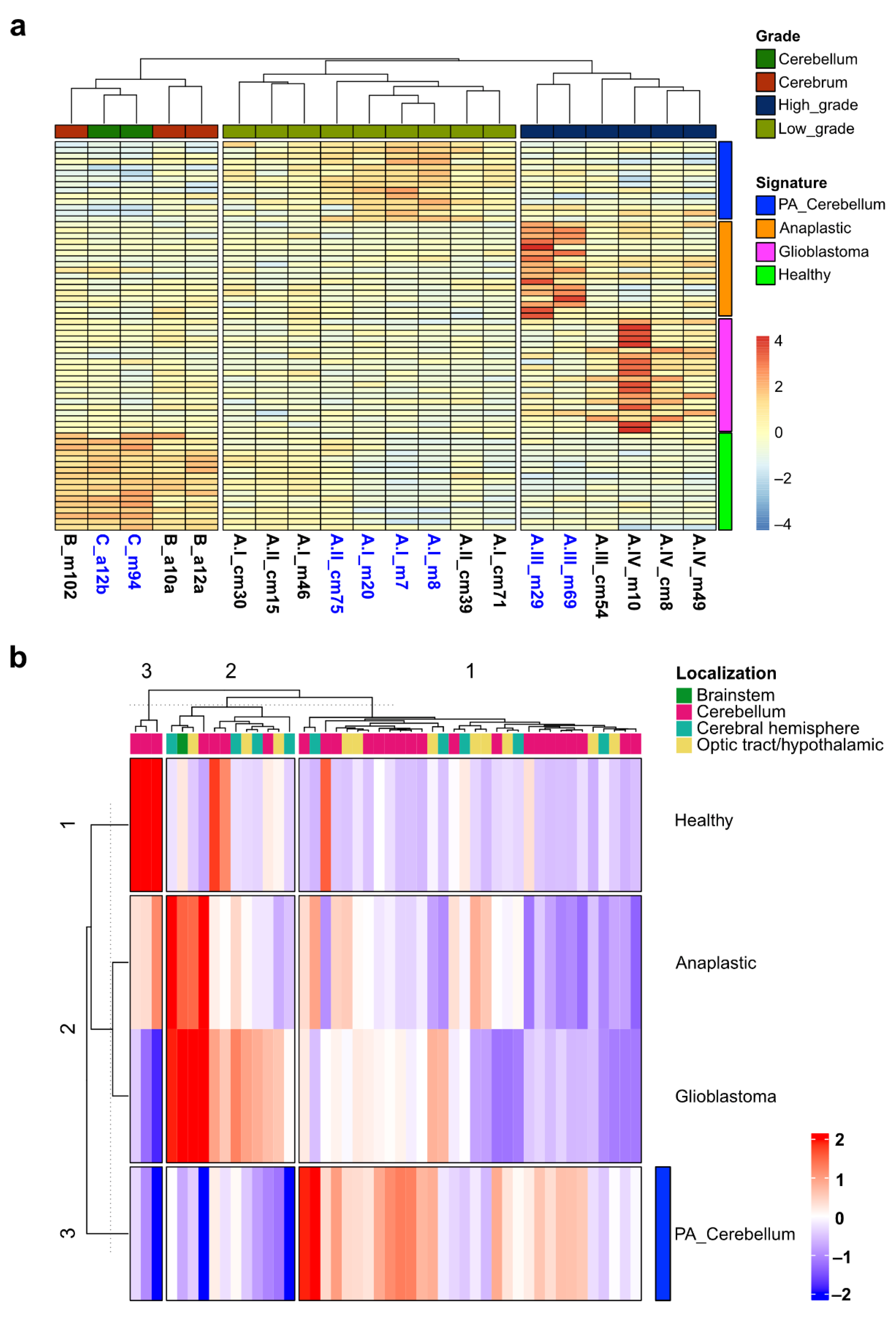
2.1. Clustering of Astrocytoma Transcriptomic Profiles according to Their Grade of Malignancy
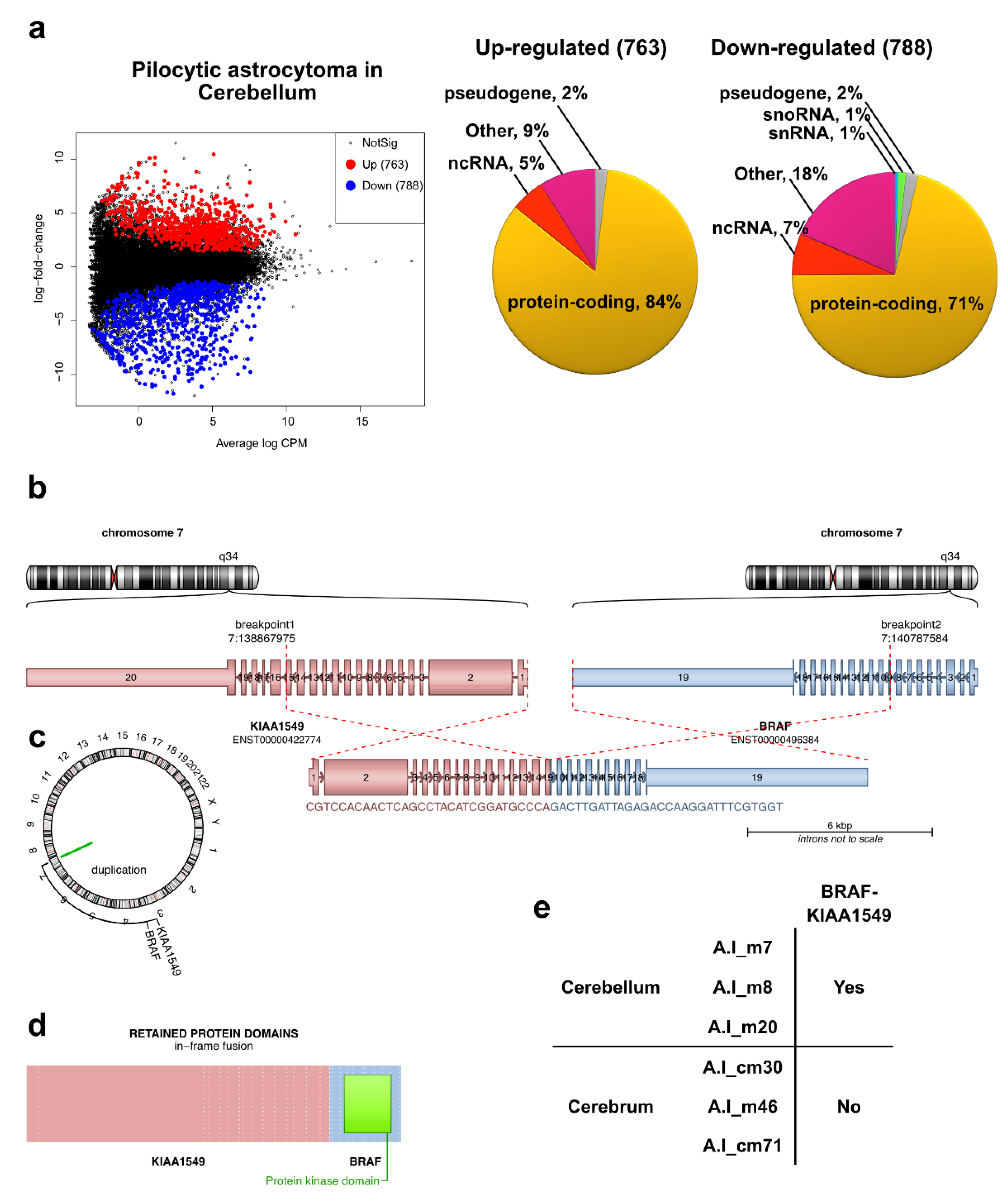
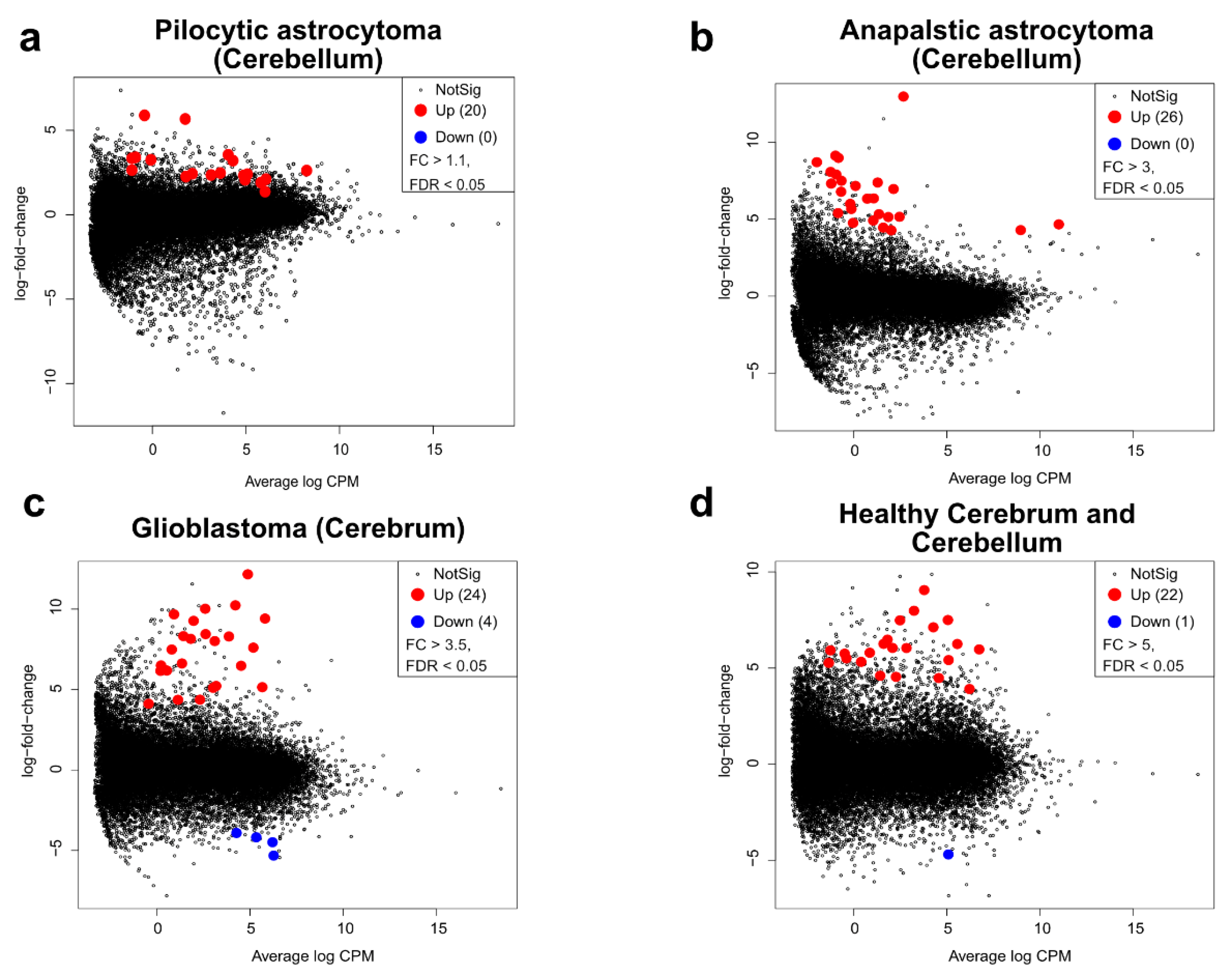
2.2. Transcriptomic Profile and Altered Biological Functions in Pilocytic Astrocytoma
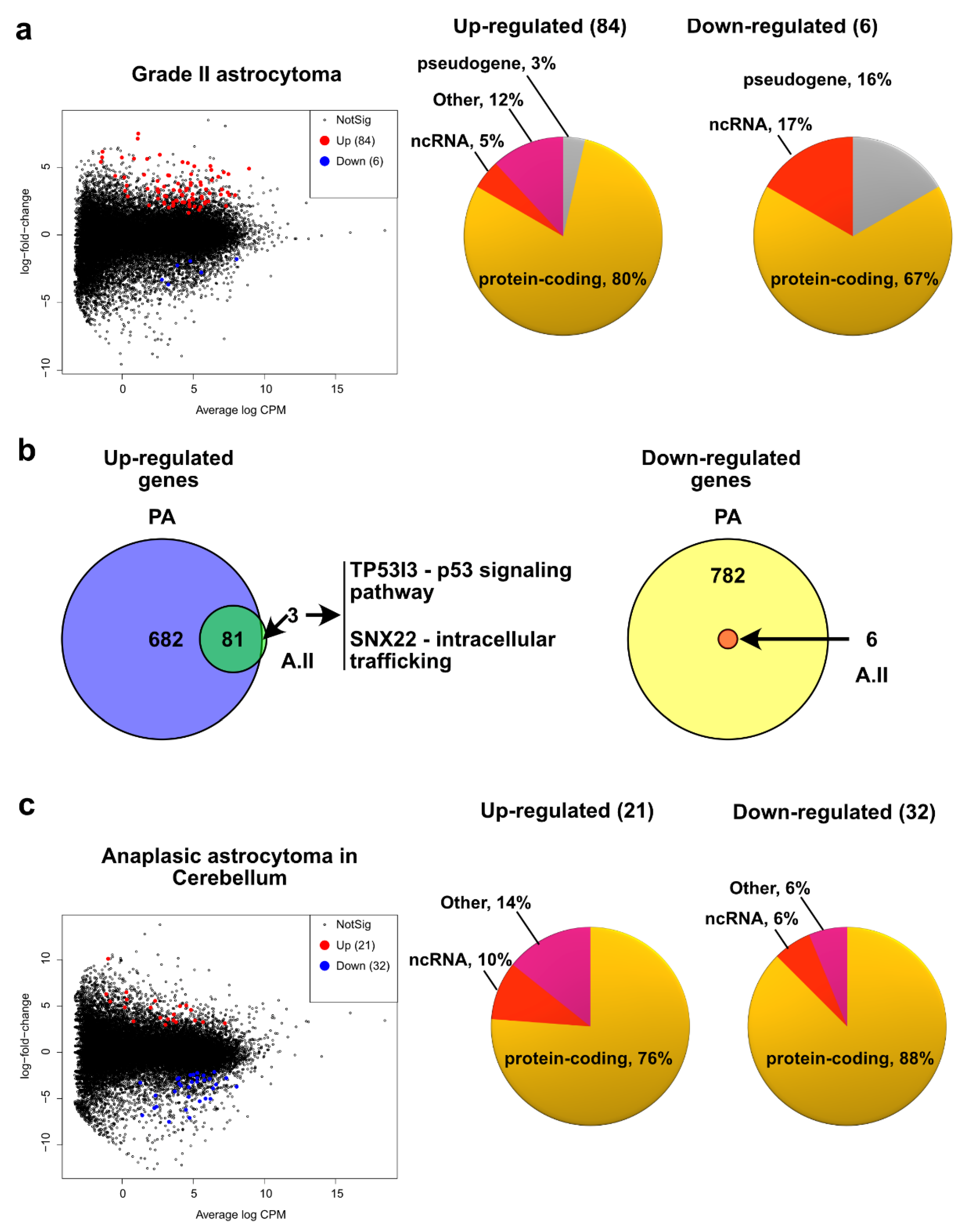
2.3. Grade II Astrocytoma
2.4. Pediatric High Grade Astrocytoma—Anaplastic Astrocytoma
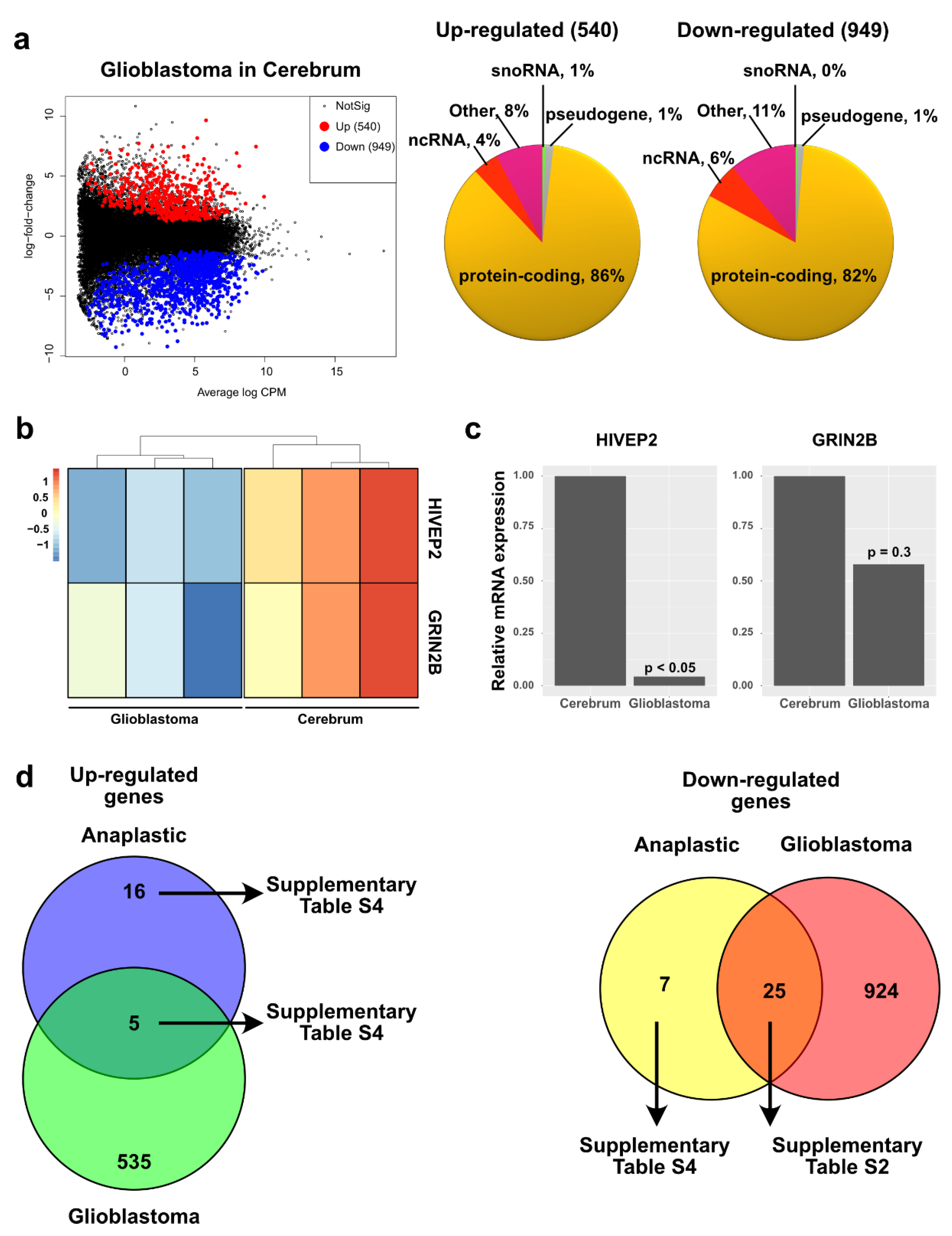
2.5. Pediatric High Grade Astrocytoma—Glioblastoma
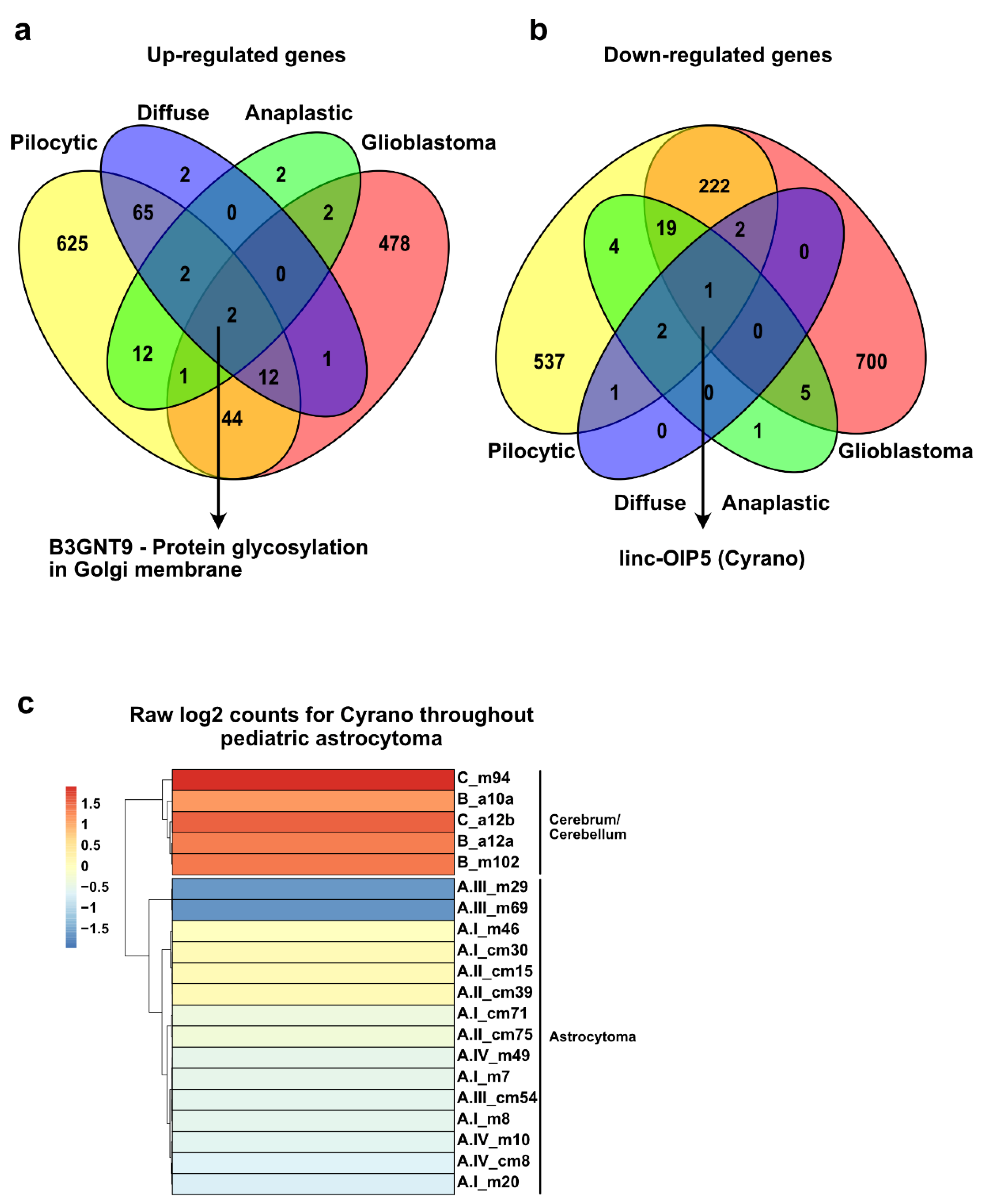
2.6. Similarities among All Grades of Astrocytoma Reveal a Putative General Tumorigenesis Factor
2.7. Unique Upregulated Genes in each Grade of Astrocytoma May Function as a Transcriptional Signature to Classify These Tumors
3. Discussion
3.1. Altered Pathways in Pediatric Astrocytoma
3.2. Low-Grade Astrocytoma
3.3. High-Grade Astrocytoma
3.4. Cyrano, a lncRNA That May Be Involved in the Tumorigenesis of Pediatric Astrocytoma
3.5. A Transcriptional Signature to Classify Pediatric Astrocytoma
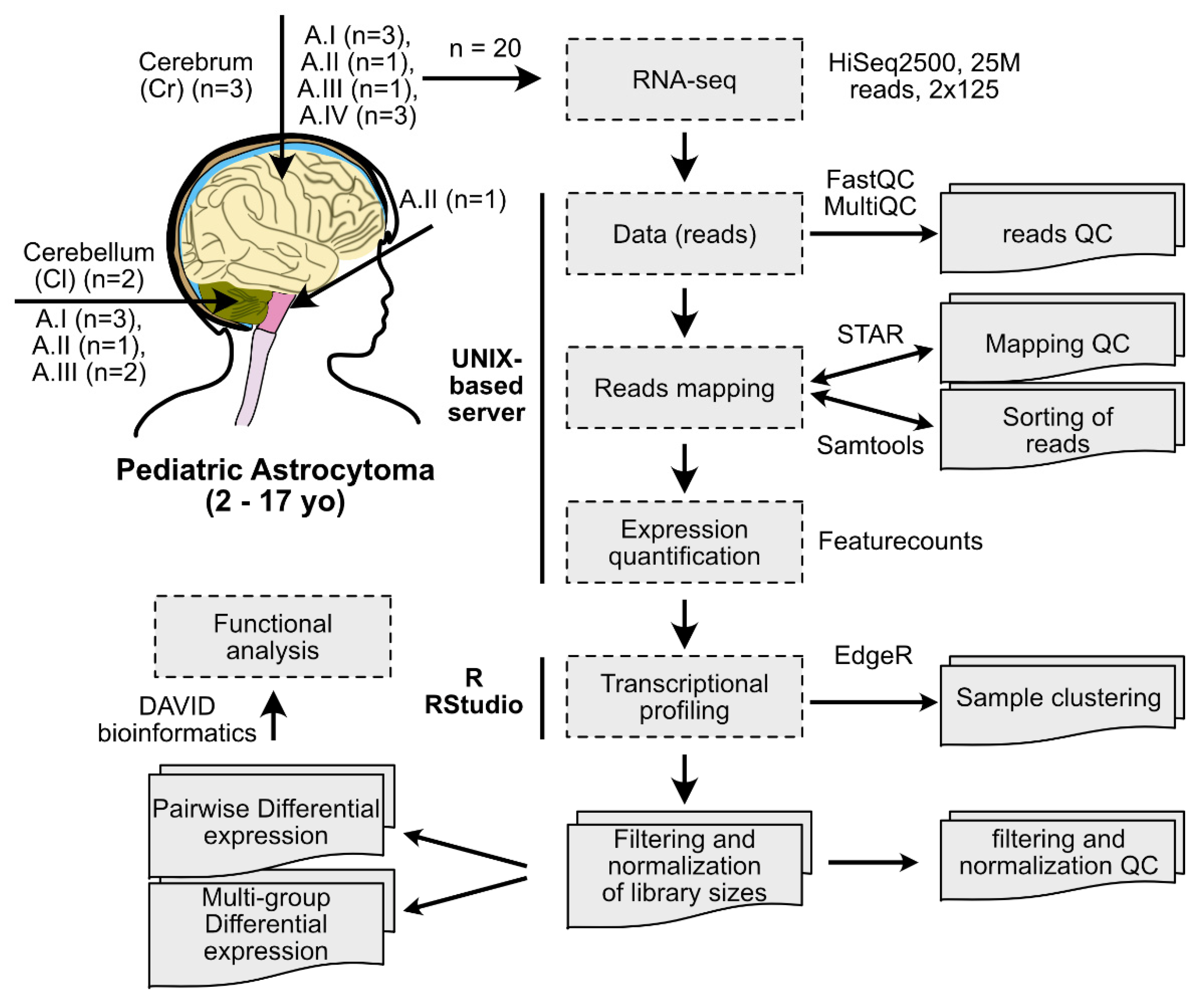
4. Materials and Methods
4.1. Human Samples
4.2. RNA Extractions and NGS
4.3. Data Analysis
4.4. Genotyping
4.5. RT-qPCR
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chico-Ponce de Leon, F.; Castro-Sierra, E.; Perezpena-Diazconti, M.; Gordillo-Dominguez, L.F.; Santana-Montero, B.L.; Rocha-Rivero, L.E.; Vaca-Ruiz, M.E.; Ríos-Alanís, M.; Sánchez-Herrera, F.; Valdés-Ordoño, R. Pediatric Intracranial Tumors. Bol. Med. Hosp. Infant. Mex. 2016, 63, 367–381. [Google Scholar]
- Bhatia, A.; Pruthi, S. Pediatric Brain Tumors: A Different Ball Game. Semin. Roentgenol. 2018, 53, 77–100. [Google Scholar] [CrossRef] [PubMed]
- Pollack, I.F.; Agnihotri, S.; Broniscer, A. Childhood Brain Tumors: Current Management, Biological Insights, and Future Directions. J. Neurosurg. Pediatr. 2019, 23, 261–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A Summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- PDQ Pediatric Treatment Editorial Board. Childhood Astrocytomas Treatment (PDQ(R)): Health Professional Version. In PDQ Cancer Information Summaries; National Cancer Institute (US): Bethesda, MD, USA, 2002. [Google Scholar]
- Louis, D.N.; Ohgaki, H.; Wiestler, O.D.; Cavenee, W.K.; Burger, P.C.; Jouvet, A.; Scheithauer, B.W.; Kleihues, P. The 2007 WHO Classification of Tumours of the Central Nervous System. Acta Neuropathol. 2007, 114, 97–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, N.L.; Hiroko, O.; Otmar, D.W.; Webster, K.C. (Eds.) WHO Classification of Tumours of the Central Nervous System; Revised 4t.; International Agency for Research on Cancer (IARC): Lyon, France, 2007. [Google Scholar]
- Ghafouri-Fard, S.; Dashti, S.; Farsi, M.; Hussen, B.M.; Taheri, M. A Review on the Role of Oncogenic LncRNA OIP5-AS1 in Human Malignancies. Biomed. Pharmacother. 2021, 137, 111366. [Google Scholar] [CrossRef]
- Conesa, A.; Madrigal, P.; Tarazona, S.; Gomez-Cabrero, D.; Cervera, A.; McPherson, A.; Szcześniak, M.W.; Gaffney, D.J.; Elo, L.L.; Zhang, X.; et al. A Survey of Best Practices for RNA-Seq Data Analysis. Genome Biol. 2016, 17, 13. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; He, X.; Hoadley, K.A.; Parker, J.S.; Hayes, D.N.; Perou, C.M. Comparison of RNA-Seq by Poly (A) Capture, Ribosomal RNA Depletion, and DNA Microarray for Expression Profiling. BMC Genomics 2014, 15, 419. [Google Scholar] [CrossRef] [Green Version]
- Salles, D.; Santino, S.F.; Ribeiro, D.A.; Malinverni, A.C.M.; Stávale, J.N. The Involvement of the MAPK Pathway in Pilocytic Astrocytomas. Pathol. Res. Pract. 2022, 232, 153821. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Tan, Q.; Collins, J.R.; Alvord, W.G.; Roayaei, J.; Stephens, R.; Baseler, M.W.; Lane, H.C.; Lempicki, R.A. The DAVID Gene Functional Classification Tool: A Novel Biological Module-Centric Algorithm to Functionally Analyze Large Gene Lists. Genome Biol. 2007, 8, R183. [Google Scholar] [CrossRef] [Green Version]
- Ryall, S.; Zapotocky, M.; Fukuoka, K.; Nobre, L.; Guerreiro Stucklin, A.; Bennett, J.; Siddaway, R.; Li, C.; Pajovic, S.; Arnoldo, A.; et al. Integrated Molecular and Clinical Analysis of 1000 Pediatric Low-Grade Gliomas. Cancer Cell 2020, 37, 569–583. [Google Scholar] [CrossRef] [PubMed]
- Ryall, S.; Tabori, U.; Hawkins, C. Pediatric Low-Grade Glioma in the Era of Molecular Diagnostics. Acta Neuropathol. Commun. 2020, 8, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, D.T.W.; Gronych, J.; Lichter, P.; Witt, O.; Pfister, S.M. MAPK Pathway Activation in Pilocytic Astrocytoma. Cell. Mol. Life Sci. 2012, 69, 1799–1811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rorive, S.; Maris, C.; Debeir, O.; Sandras, F.; Vidaud, M.; Bièche, I.; Salmon, I.; Decaestecker, C. Exploring the Distinctive Biological Characteristics of Pilocytic and Low-Grade Diffuse Astrocytomas Using Microarray Gene Expression Profiles. J. Neuropathol. Exp. Neurol. 2006, 65, 794–807. [Google Scholar] [CrossRef] [Green Version]
- Seifert, M.; Garbe, M.; Friedrich, B.; Mittelbronn, M.; Klink, B. Comparative Transcriptomics Reveals Similarities and Differences between Astrocytoma Grades. BMC Cancer 2015, 15, 952. [Google Scholar] [CrossRef]
- Ryall, S.; Tabori, U.; Hawkins, C. A Comprehensive Review of Paediatric Low-Grade Diffuse Glioma: Pathology, Molecular Genetics and Treatment. Brain Tumor Pathol. 2017, 34, 51–61. [Google Scholar] [CrossRef]
- Nishikawa, R. Pediatric and Adult Gliomas: How Different Are They? Neuro. Oncol. 2010, 12, 1203–1204. [Google Scholar]
- Torres-Caballero, A.; Serrato, A.; López-Santaella, T.; Ortiz, R.; de León, F.C.-P.; González-Carranza, V.; Torres-García, S.; Arenas-Huertero, F.; Hernández-Hernández, A. Evaluation of the Melting Temperature of TaqMan Probes as a Genotyping Method for IDH1, IDH2, and H3F3A in Pediatric Astrocytomas. Bol. Med. Hosp. Infant. Mex. 2020, 77, 303–311. [Google Scholar] [CrossRef]
- Sturm, D.; Pfister, S.M.; Jones, D.T.W. Pediatric Gliomas: Current Concepts on Diagnosis, Biology, and Clinical Management. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2017, 35, 2370–2377. [Google Scholar] [CrossRef]
- Raj-Kumar, P.-K.; Liu, J.; Hooke, J.A.; Kovatich, A.J.; Kvecher, L.; Shriver, C.D.; Hu, H. PCA-PAM50 Improves Consistency between Breast Cancer Intrinsic and Clinical Subtyping Reclassifying a Subset of Luminal A Tumors as Luminal B. Sci. Rep. 2019, 9, 7956. [Google Scholar] [CrossRef] [Green Version]
- Schettini, F.; Chic, N.; Brasó-Maristany, F.; Paré, L.; Pascual, T.; Conte, B.; Martínez-Sáez, O.; Adamo, B.; Vidal, M.; Barnadas, E.; et al. Clinical, Pathological, and PAM50 Gene Expression Features of HER2-Low Breast Cancer. NPJ Breast Cancer 2021, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Prat, A.; Guarneri, V.; Paré, L.; Griguolo, G.; Pascual, T.; Dieci, M.V.; Chic, N.; González-Farré, B.; Frassoldati, A.; Sanfeliu, E.; et al. A Multivariable Prognostic Score to Guide Systemic Therapy in Early-Stage HER2-Positive Breast Cancer: A Retrospective Study with an External Evaluation. Lancet. Oncol. 2020, 21, 1455–1464. [Google Scholar] [CrossRef]
- Pernas, S.; Petit, A.; Climent, F.; Paré, L.; Perez-Martin, J.; Ventura, L.; Bergamino, M.; Galván, P.; Falo, C.; Morilla, I.; et al. PAM50 Subtypes in Baseline and Residual Tumors Following Neoadjuvant Trastuzumab-Based Chemotherapy in HER2-Positive Breast Cancer: A Consecutive-Series from a Single Institution. Front. Oncol. 2019, 9, 707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schettini, F.; Prat, A. Dissecting the Biological Heterogeneity of HER2-Positive Breast Cancer. Breast 2021, 59, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.S.; Mullins, M.; Cheang, M.C.U.; Leung, S.; Voduc, D.; Vickery, T.; Davies, S.; Fauron, C.; He, X.; Hu, Z.; et al. Supervised Risk Predictor of Breast Cancer Based on Intrinsic Subtypes. J. Clin. Oncol. 2009, 27, 1160–1167. [Google Scholar] [CrossRef]
- Davey, M.G.; Ryan, É.J.; Abd Elwahab, S.; Elliott, J.A.; McAnena, P.F.; Sweeney, K.J.; Malone, C.M.; McLaughlin, R.; Barry, M.K.; Keane, M.M.; et al. Clinicopathological Correlates, Oncological Impact, and Validation of Oncotype DXTM in a European Tertiary Referral Centre. Breast J. 2021, 27, 521–528. [Google Scholar] [CrossRef]
- Davey, M.G.; Ryan, É.J.; Boland, M.R.; Barry, M.K.; Lowery, A.J.; Kerin, M.J. Clinical Utility of the 21-Gene Assay in Predicting Response to Neoadjuvant Endocrine Therapy in Breast Cancer: A Systematic Review and Meta-Analysis. Breast 2021, 58, 113–120. [Google Scholar] [CrossRef]
- Perou, C.M.; Sørlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular Portraits of Human Breast Tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef] [Green Version]
- Sorlie, T.; Tibshirani, R.; Parker, J.; Hastie, T.; Marron, J.S.; Nobel, A.; Deng, S.; Johnsen, H.; Pesich, R.; Geisler, S.; et al. Repeated Observation of Breast Tumor Subtypes in Independent Gene Expression Data Sets. Proc. Natl. Acad. Sci. USA 2003, 100, 8418–8423. [Google Scholar] [CrossRef] [Green Version]
- Sørlie, T.; Perou, C.M.; Tibshirani, R.; Aas, T.; Geisler, S.; Johnsen, H.; Hastie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; et al. Gene Expression Patterns of Breast Carcinomas Distinguish Tumor Subclasses with Clinical Implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef] [Green Version]
- Yi, M.; Nissley, D.V.; McCormick, F.; Stephens, R.M. SsGSEA Score-Based Ras Dependency Indexes Derived from Gene Expression Data Reveal Potential Ras Addiction Mechanisms with Possible Clinical Implications. Sci. Rep. 2020, 10, 10258. [Google Scholar] [CrossRef] [PubMed]
- Barbie, D.A.; Tamayo, P.; Boehm, J.S.; Kim, S.Y.; Moody, S.E.; Dunn, I.F.; Schinzel, A.C.; Sandy, P.; Meylan, E.; Scholl, C.; et al. Systematic RNA Interference Reveals That Oncogenic KRAS-Driven Cancers Require TBK1. Nature 2009, 462, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewski, K.; Jarzab, M.; Pfeifer, A.; Oczko-Wojciechowska, M.; Jarzab, B.; Liberski, P.P.; Zakrzewska, M. Transcriptional Profiles of Pilocytic Astrocytoma Are Related to Their Three Different Locations, but Not to Radiological Tumor Features. BMC Cancer 2015, 15, 778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, M.K.; Mansur, D.B.; Reifenberger, G.; Perry, A.; Leonard, J.R.; Aldape, K.D.; Albin, M.G.; Emnett, R.J.; Loeser, S.; Watson, M.A.; et al. Distinct Genetic Signatures among Pilocytic Astrocytomas Relate to Their Brain Region Origin. Cancer Res. 2007, 67, 890–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, S.R.; Witt, H.; Hovestadt, V.; Zucknick, M.; Kool, M.; Pearson, D.M.; Korshunov, A.; Ryzhova, M.; Ichimura, K.; Jabado, N.; et al. Differential Expression and Methylation of Brain Developmental Genes Define Location-Specific Subsets of Pilocytic Astrocytoma. Acta Neuropathol. 2013, 126, 291–301. [Google Scholar] [CrossRef]
- Aichmüller, C.F.; Iskar, M.; Jones, D.T.W.; Korshunov, A.; Radlwimmer, B.; Kool, M.; Ernst, A.; Pfister, S.M.; Lichter, P.; Zapatka, M. Pilocytic Astrocytoma Demethylation and Transcriptional Landscapes Link BZIP Transcription Factors to Immune Response. Neuro. Oncol. 2020, 22, 1327–1338. [Google Scholar] [CrossRef]
- Zhou, R.; Man, Y. Integrated Analysis of DNA Methylation Profiles and Gene Expression Profiles to Identify Genes Associated with Pilocytic Astrocytomas. Mol. Med. Rep. 2016, 13, 3491–3497. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Hara, A.; Homma, T.; Yonekawa, Y.; Ohgaki, H. Altered Expression of Immune Defense Genes in Pilocytic Astrocytomas. J. Neuropathol. Exp. Neurol. 2005, 64, 891–901. [Google Scholar] [CrossRef]
- Wong, K.-K.; Chang, Y.-M.; Tsang, Y.T.M.; Perlaky, L.; Su, J.; Adesina, A.; Armstrong, D.L.; Bhattacharjee, M.; Dauser, R.; Blaney, S.M.; et al. Expression Analysis of Juvenile Pilocytic Astrocytomas by Oligonucleotide Microarray Reveals Two Potential Subgroups. Cancer Res. 2005, 65, 76–84. [Google Scholar] [CrossRef]
- Jones, C.; Karajannis, M.A.; Jones, D.T.W.; Kieran, M.W.; Monje, M.; Baker, S.J.; Becher, O.J.; Cho, Y.-J.; Gupta, N.; Hawkins, C.; et al. Pediatric High-Grade Glioma: Biologically and Clinically in Need of New Thinking. Neuro. Oncol. 2017, 19, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Gerges, N.; Fontebasso, A.M.; Albrecht, S.; Faury, D.; Jabado, N. Pediatric High-Grade Astrocytomas: A Distinct Neuro-Oncological Paradigm. Genome Med. 2013, 5, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauser, P. Classification and Treatment of Pediatric Gliomas in the Molecular Era. Children 2021, 8, 739. [Google Scholar] [CrossRef] [PubMed]
- Ulitsky, I.; Shkumatava, A.; Jan, C.H.; Sive, H.; Bartel, D.P. Conserved Function of LincRNAs in Vertebrate Embryonic Development despite Rapid Sequence Evolution. Cell 2011, 147, 1537–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, G.-W.; Wu, L.; Kuang, W.; Chen, Y.; Zhu, X.-G.; Guo, H.; Lang, H.-L. Knockdown of Linc-OIP5 Inhibits Proliferation and Migration of Glioma Cells through down-Regulation of YAP-NOTCH Signaling Pathway. Gene 2017, 610, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.A.M.; Srinivasan, S.; Patric, I.R.P.; Hegde, A.S.; Chandramouli, B.A.; Arimappamagan, A.; Santosh, V.; Kondaiah, P.; Rao, M.R.S.; Somasundaram, K. A 16-Gene Signature Distinguishes Anaplastic Astrocytoma from Glioblastoma. PLoS ONE 2014, 9, e85200. [Google Scholar] [CrossRef]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize Analysis Results for Multiple Tools and Samples in a Single Report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve Years of SAMtools and BCFtools. Gigascience 2021, 10, giab008. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. The R Package Rsubread Is Easier, Faster, Cheaper and Better for Alignment and Quantification of RNA Sequencing Reads. Nucleic Acids Res. 2019, 47, e47. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential Expression Analysis of Multifactor RNA-Seq Experiments with Respect to Biological Variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Lun, A.T.L.; Smyth, G.K. From Reads to Genes to Pathways: Differential Expression Analysis of RNA-Seq Experiments Using Rsubread and the EdgeR Quasi-Likelihood Pipeline. F1000Research 2016, 5, 1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics Enrichment Tools: Paths toward the Comprehensive Functional Analysis of Large Gene Lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Uhrig, S.; Ellermann, J.; Walther, T.; Burkhardt, P.; Fröhlich, M.; Hutter, B.; Toprak, U.H.; Neumann, O.; Stenzinger, A.; Scholl, C.; et al. Accurate and Efficient Detection of Gene Fusions from RNA Sequencing Data. Genome Res. 2021, 31, 448–460. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene Set Enrichment Analysis: A Knowledge-Based Approach for Interpreting Genome-Wide Expression Profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Bengtsson, H.; Corrada Bravo, H.; Gentleman, R.; Hossjer, O.; Jaffee, H.; Jiang, D. Langfelder MatrixStats: Functions That Apply to Rows and Columns of Matrices (and to Vectors); R Package Version 0.52.2. 2017. Available online: https://github.com/HenrikBengtsson/matrixStats (accessed on 16 November 2021).
- Gu, Z.; Gu, L.; Eils, R.; Schlesner, M.; Brors, B. Circlize Implements and Enhances Circular Visualization in R. Bioinformatics 2014, 30, 2811–2812. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Eils, R.; Schlesner, M. Complex Heatmaps Reveal Patterns and Correlations in Multidimensional Genomic Data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [Green Version]
- Eguía-Aguilar, P.; Gutiérrez-Castillo, L.; Pérezpeña-Díazconti, M.; García-Chéquer, J.; García-Quintana, J.; Chico-Ponce de León, F.; Gordillo-Domínguez, L.; Torres-García, S.; Arenas-Huertero, F. Expression of MicroRNAs in Tumors of the Central Nervous System in Pediatric Patients in México. Child’s Nerv. Syst. 2017, 33, 2117–2128. [Google Scholar] [CrossRef]
- Ahmed, M.; Kim, D.R. Pcr: An R Package for Quality Assessment, Analysis and Testing of QPCR Data. PeerJ 2018, 6, e4473. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández-Hernández, A.; López-Santaella, T.; Torres-Caballero, A.; Serrato, A.; Torres-Flores, U.; Montesinos-Valencia, D.; Chico-Ponce de León, F.; González-Carranza, V.; Torres-García, S.; Rebollar-Vega, R.; et al. The Transcriptomic Landscape of Pediatric Astrocytoma. Int. J. Mol. Sci. 2022, 23, 12696. https://doi.org/10.3390/ijms232012696
Hernández-Hernández A, López-Santaella T, Torres-Caballero A, Serrato A, Torres-Flores U, Montesinos-Valencia D, Chico-Ponce de León F, González-Carranza V, Torres-García S, Rebollar-Vega R, et al. The Transcriptomic Landscape of Pediatric Astrocytoma. International Journal of Molecular Sciences. 2022; 23(20):12696. https://doi.org/10.3390/ijms232012696
Chicago/Turabian StyleHernández-Hernández, Abrahan, Tayde López-Santaella, Aranxa Torres-Caballero, Amarantha Serrato, Ulises Torres-Flores, Diego Montesinos-Valencia, Fernando Chico-Ponce de León, Vicente González-Carranza, Samuel Torres-García, Rosa Rebollar-Vega, and et al. 2022. "The Transcriptomic Landscape of Pediatric Astrocytoma" International Journal of Molecular Sciences 23, no. 20: 12696. https://doi.org/10.3390/ijms232012696