
Sex Difference Leads to Differential Gene Expression Patterns and Therapeutic Efficacy in Mucopolysaccharidosis IVA Murine Model Receiving AAV8 Gene Therapy

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Sex, Promoter, and Mouse Model Comparison
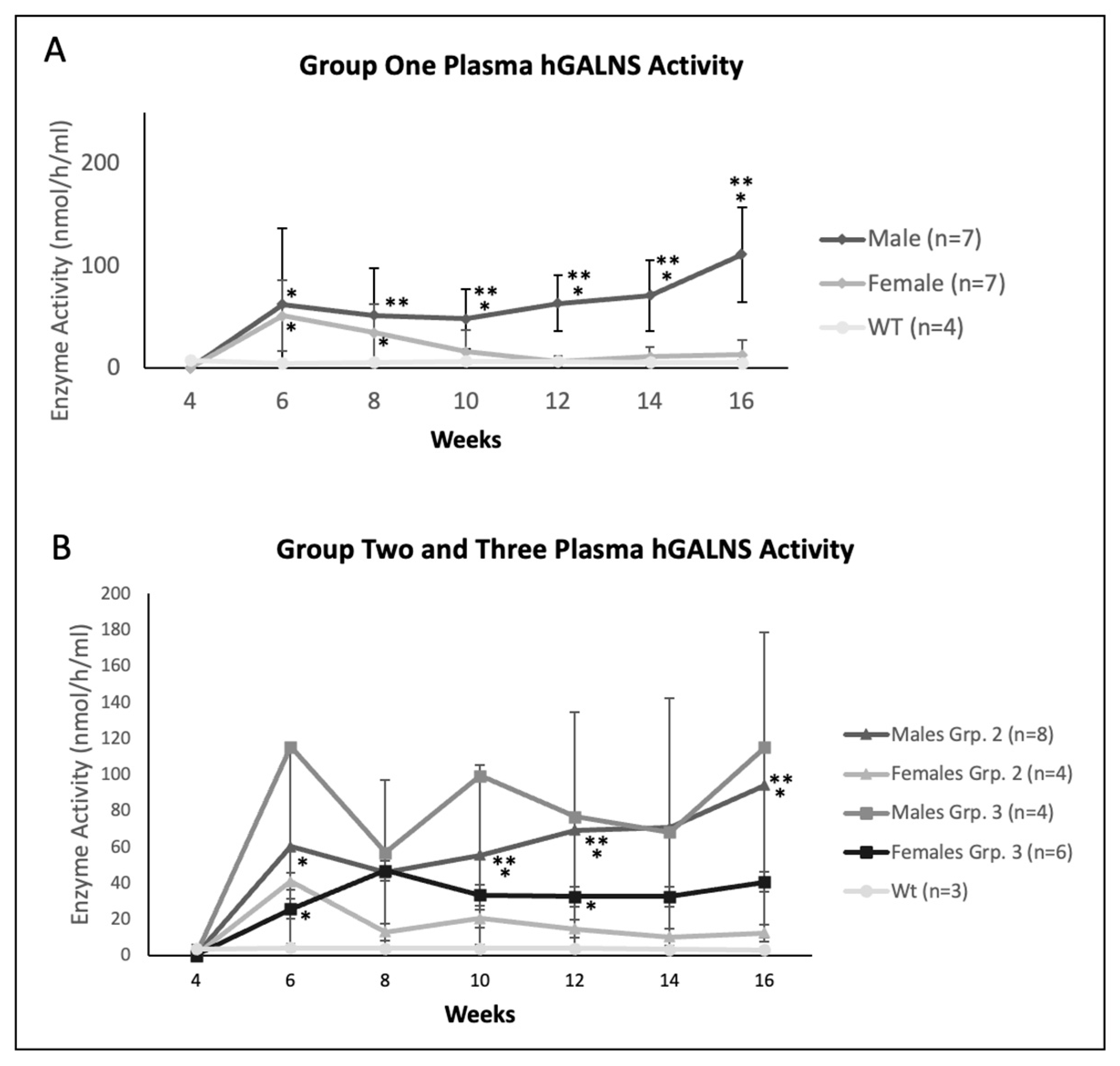
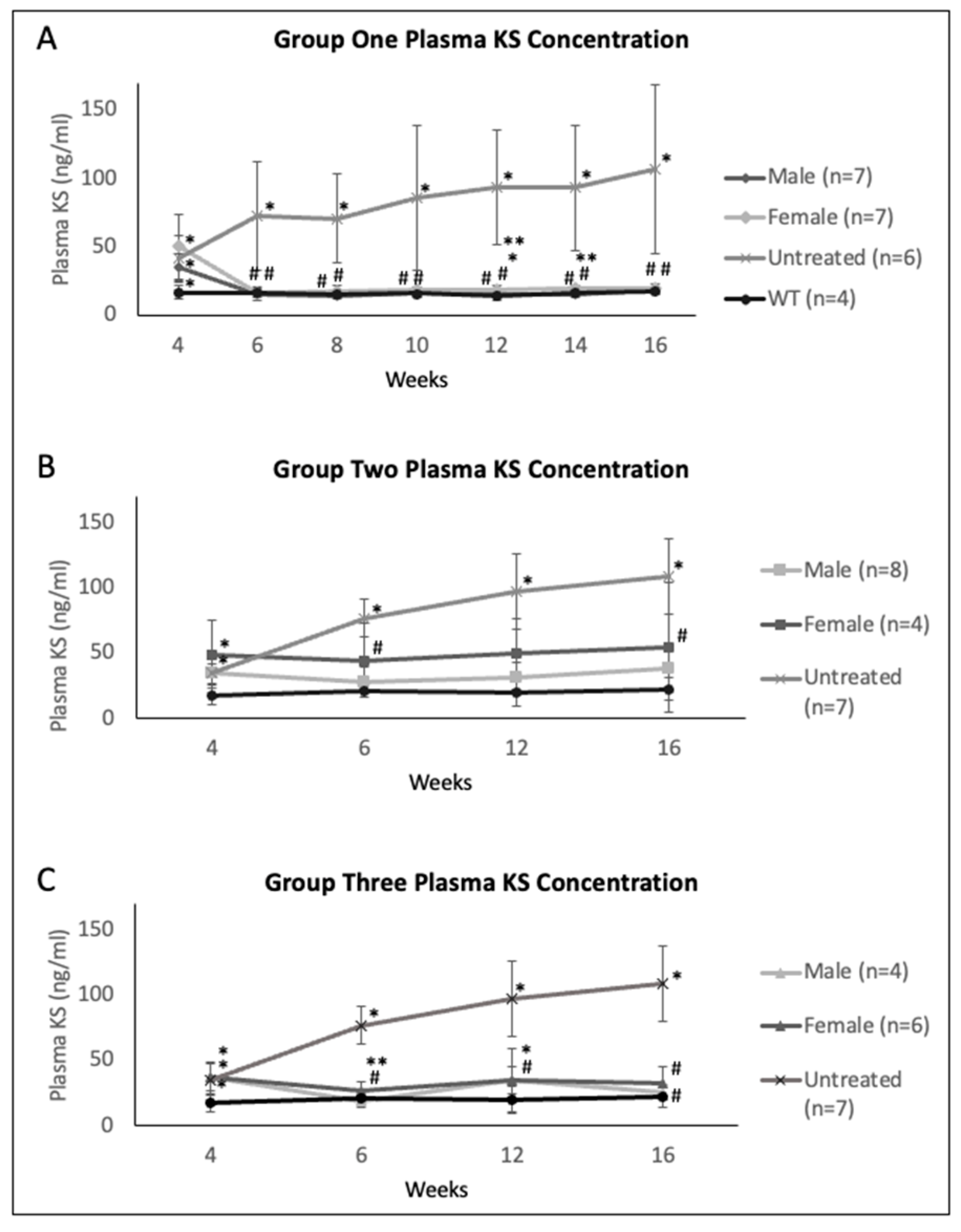
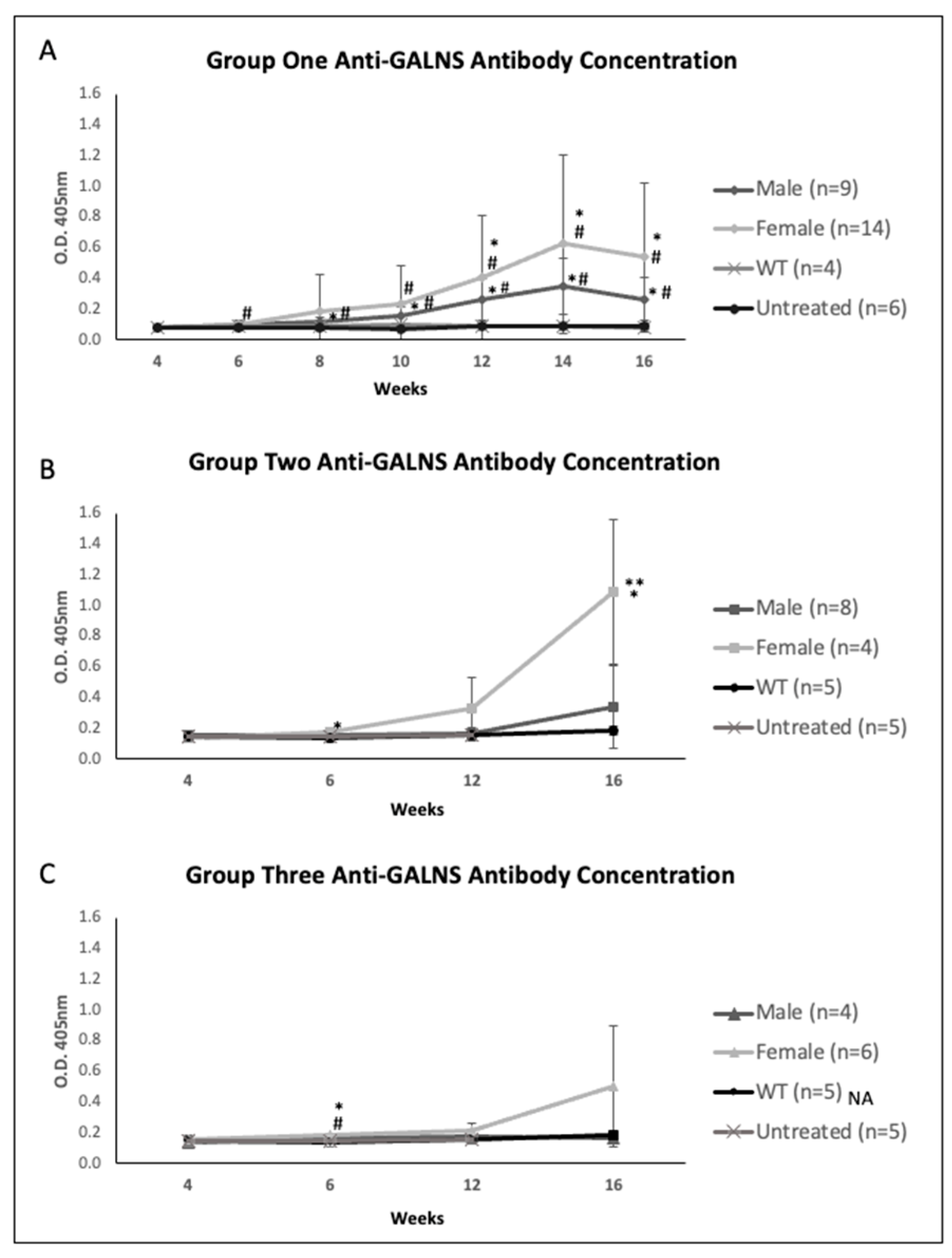
2.2. Plasma Enzyme Activity, KS, and Anti-GALNS Antibodies in Males versus Females
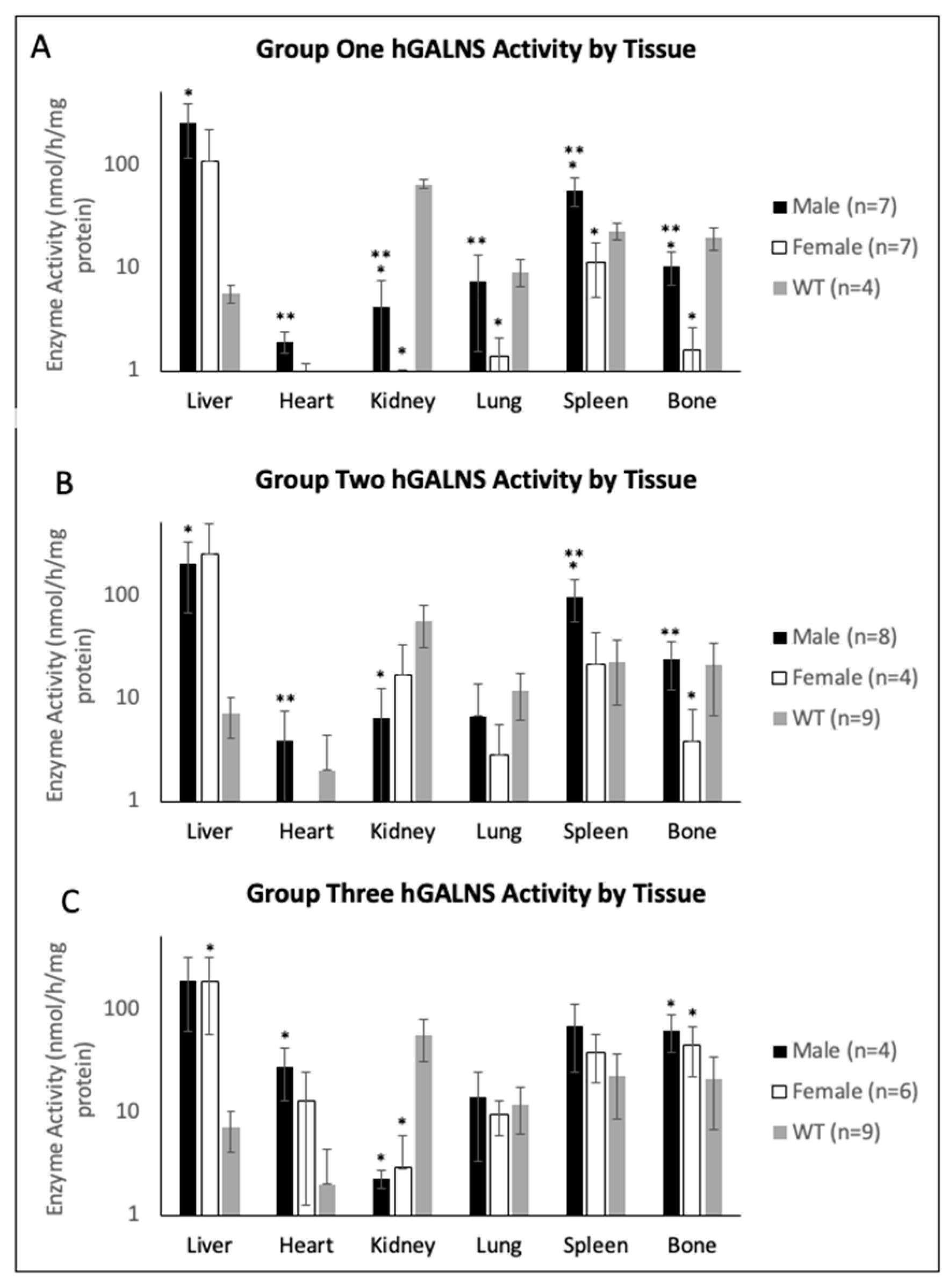
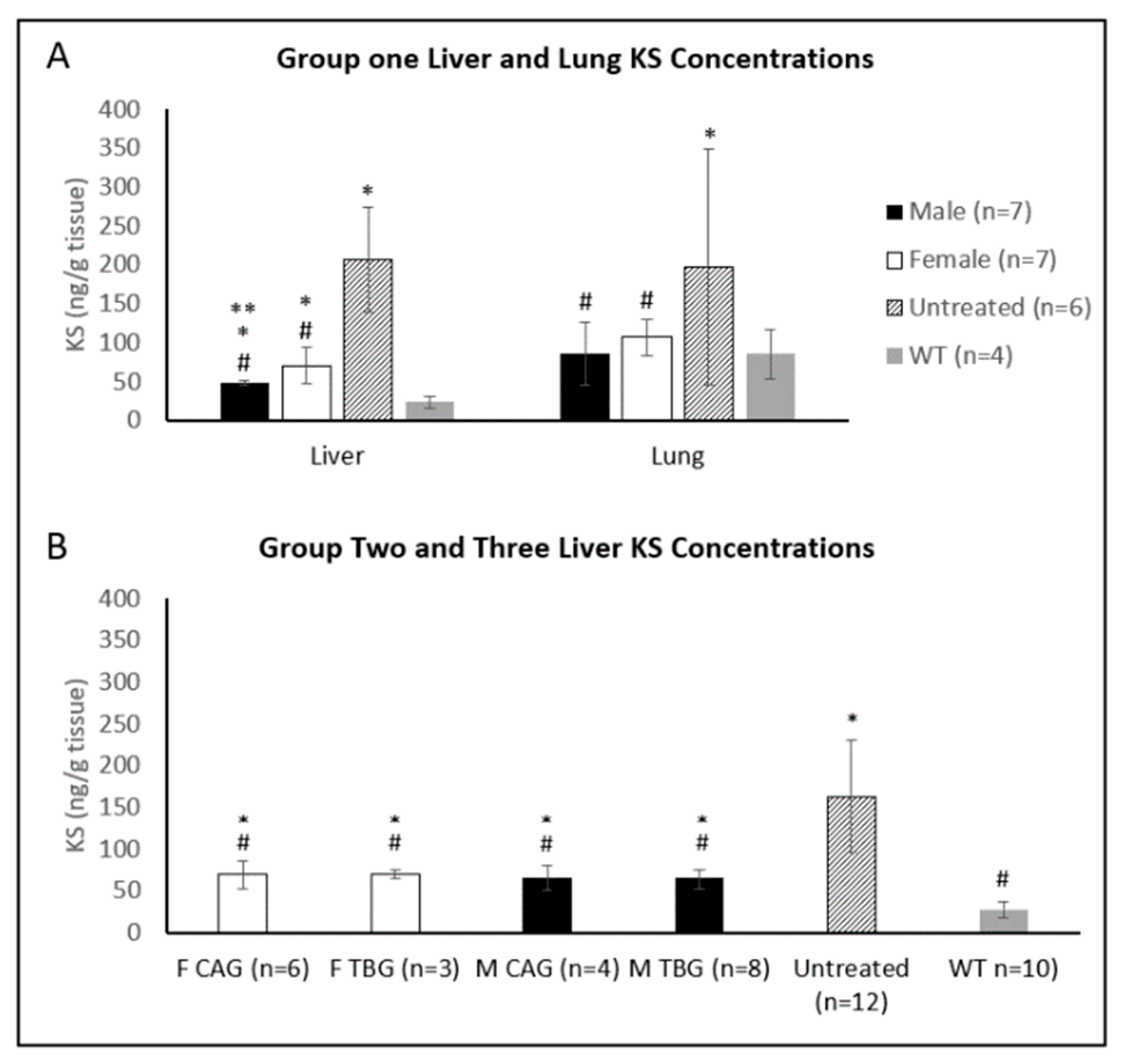
2.3. Tissue Enzyme Activity and KS Concentration
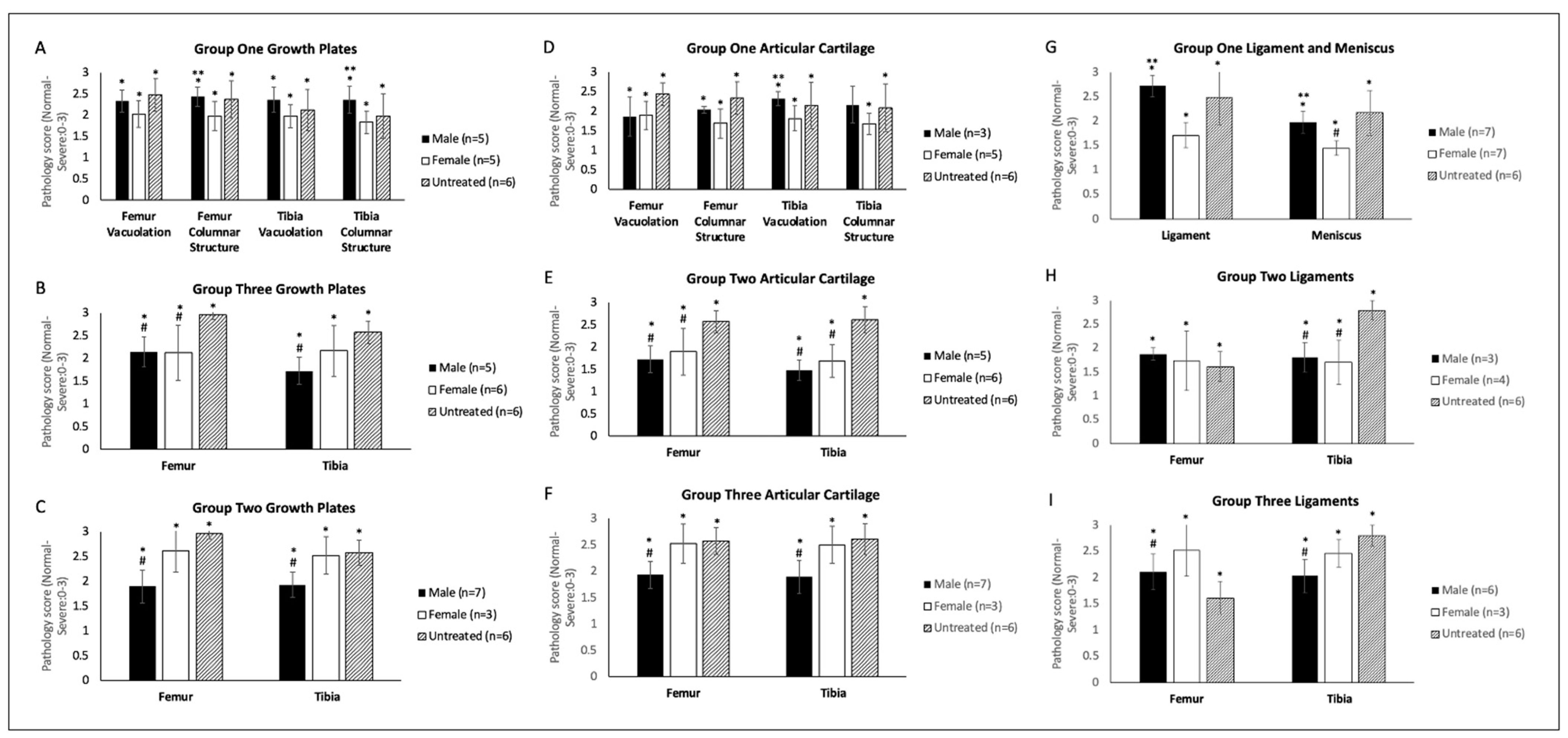
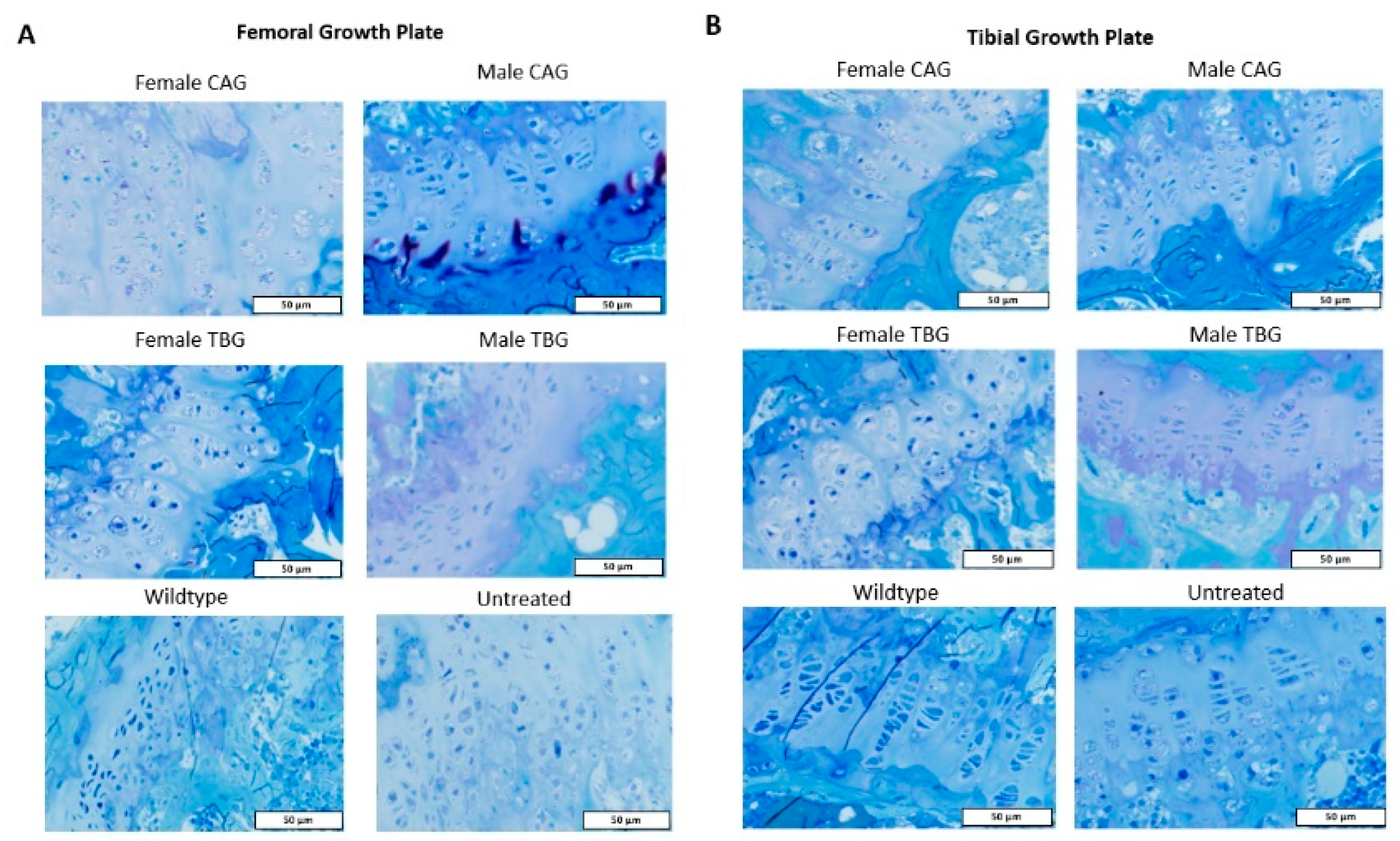
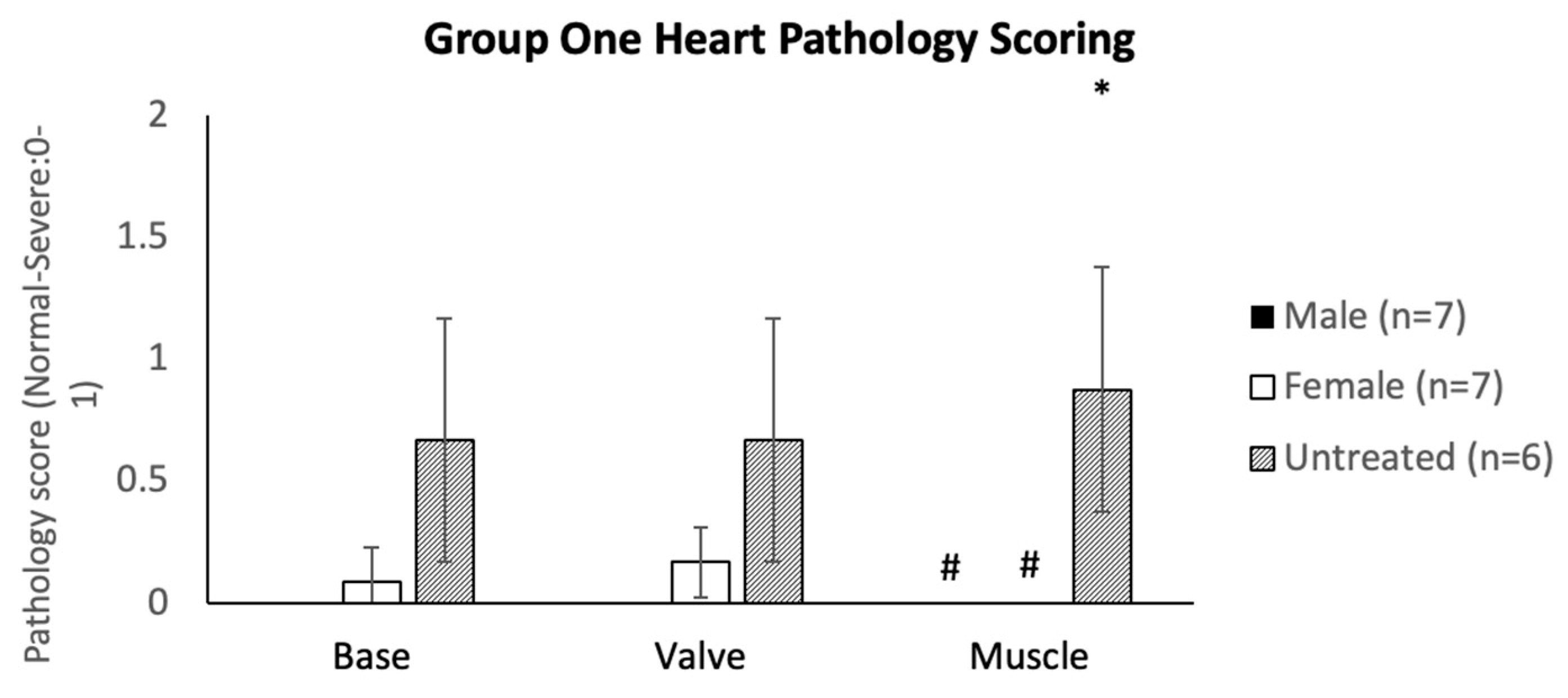
2.4. Bone and Heart Histopathology
3. Discussion
4. Materials and Methods
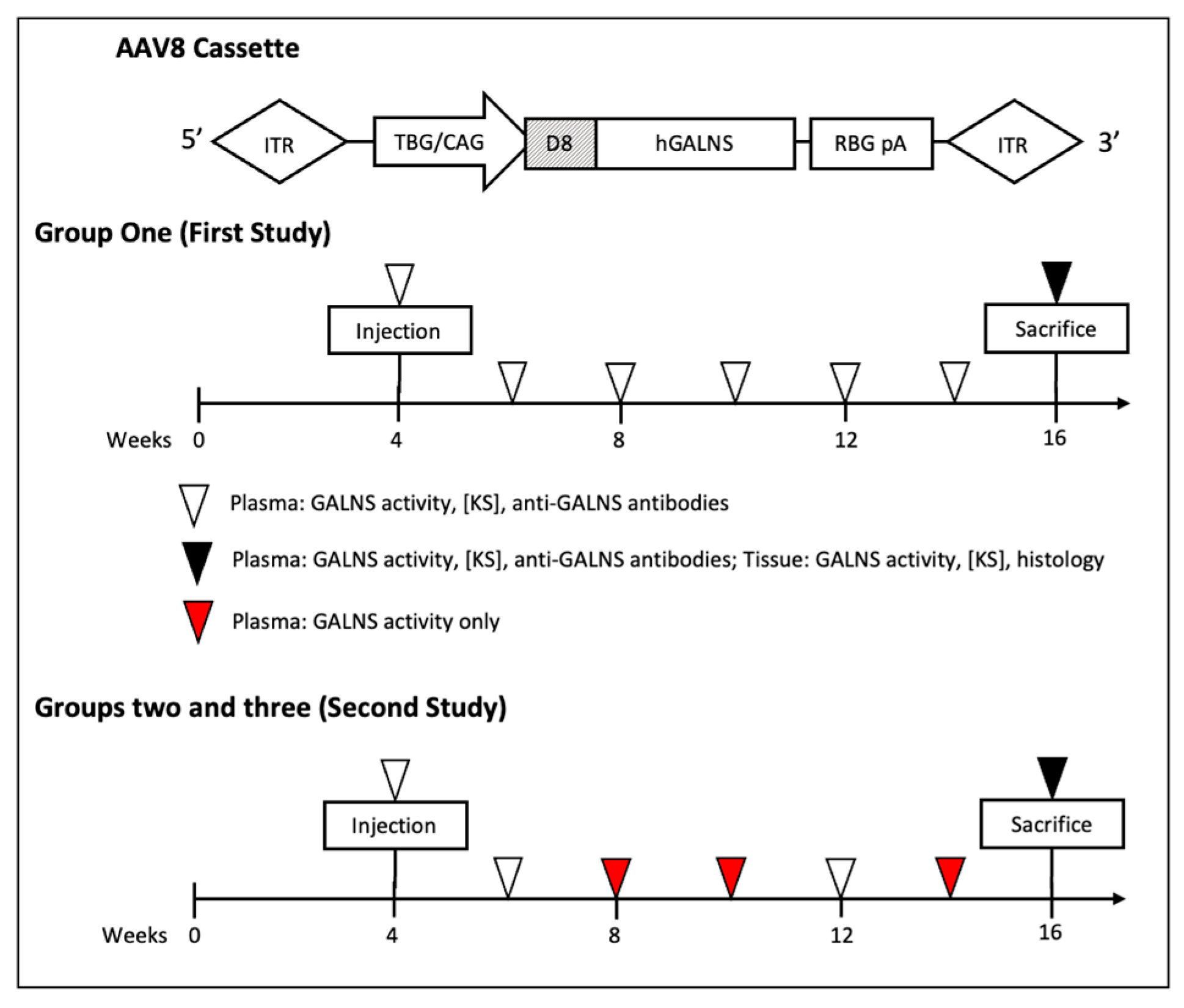
4.1. AAV Vector and Cassette Design
4.2. Murine Models
4.3. Blood and Tissue Collection
4.4. Plasma and Tissue GALNS Activity Assay
4.5. Tissue GAG Extraction and Analysis
4.6. Anti-GALNS Antibody Assay
4.7. Histopathological Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khan, S.; Alméciga-Díaz, C.J.; Sawamoto, K.; Mackenzie, W.G.; Theroux, M.C.; Pizarro, C.; Mason, R.W.; Orii, T.; Tomatsu, S. Mucopolysaccharidosis IVA and glycosaminoglycans. Mol. Genet. Metab. 2017, 120, 78–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomatsu, S.; Yasuda, E.; Patel, P.; Ruhnke, K.; Shimada, T.; MacKenzie, W.G.; Mason, R.; Thacker, M.M.; Theroux, M.; Montaño, A.M.; et al. Morquio A Syndrome: Diagnosis and Current and Future Therapies. Pediatr. Endocrinol. Rev. 2014, 12 (Suppl. 1), 141–151. [Google Scholar] [PubMed]
- Yasuda, E.; Suzuki, Y.; Shimada, T.; Sawamoto, K.; Mackenzie, W.G.; Theroux, M.C.; Pizarro, C.; Xie, L.; Miller, F.; Rahman, T.; et al. Activity of daily living for Morquio A syndrome. Mol. Genet. Metab. 2016, 118, 111–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavery, C.; Hendriksz, C. Mortality in Patients with Morquio Syndrome A. In JIMD Reports; Springer: Berlin/Heidelberg, Germany, 2014; Volume 15, pp. 59–66. [Google Scholar] [CrossRef]
- Sawamoto, K.; González, J.V.; Piechnik, M.; Otero, F.J.; Couce, M.L.; Suzuki, Y.; Tomatsu, S. Mucopolysaccharidosis IVA: Diagnosis, Treatment, and Management. Int. J. Mol. Sci. 2020, 21, 1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomatsu, S.; Sawamoto, K.; Shimada, T.; Bober, M.B.; Kubaski, F.; Yasuda, E.; Mason, R.W.; Khan, S.; Alméciga-Díaz, C.J.; A Barrera, L.; et al. Enzyme replacement therapy for treating mucopolysaccharidosis type IVA (Morquio A syndrome): Effect and limitations. Expert Opin. Orphan Drugs 2015, 3, 1279–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Y.; Musson, D.G.; Schweighardt, B.; Tompkins, T.; Jesaitis, L.; Shaywitz, A.J.; Yang, K.; O’Neill, C.A. Pharmacokinetic and pharmacodynamic evaluation of elosulfase alfa, an enzyme replacement therapy in patients with Morquio A syndrome. Clin. Pharmacokinet. 2014, 53, 1137–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doherty, C.; Stapleton, M.; Piechnik, M.; Mason, R.W.; MacKenzie, W.G.; Yamaguchi, S.; Kobayashi, H.; Suzuki, Y.; Tomatsu, S. Effect of enzyme replacement therapy on the growth of patients with Morquio A. J. Hum. Genet. 2019, 64, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Chinen, Y.; Higa, T.; Tomatsu, S.; Suzuki, Y.; Orii, T.; Hyakuna, N. Long-term therapeutic efficacy of allogenic bone marrow transplantation in a patient with mucopolysaccharidosis IVA. Mol. Genet. Metab. Rep. 2014, 1, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Luan, Z.; Jiang, H.; Fang, J.; Qin, M.; Lee, V.; Chen, J. Allogeneic Hematopoietic Stem Cell Transplantation in Thirty-Four Pediatric Cases of Mucopolysaccharidosis—A Ten-Year Report from the China Children Transplant Group. Biol. Blood Marrow Transpl. 2016, 22, 2104–2108. [Google Scholar] [CrossRef] [PubMed]
- Yabe, H.; Tanaka, A.; Chinen, Y.; Kato, S.; Sawamoto, K.; Yasuda, E.; Shintaku, H.; Suzuki, Y.; Orii, T.; Tomatsu, S. Hematopoietic stem cell transplantation for Morquio A syndrome. Mol. Genet. Metab. 2016, 117, 84–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraldi, A.; Serafini, M.; Sorrentino, N.C.; Gentner, B.; Aiuti, A.; Bernardo, M.E. Gene therapy for mucopolysaccharidoses: In vivo and ex vivo approaches. Ital. J. Pediatr. 2018, 44, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Naso, M.F.; Tomkowicz, B.; Perry, W.L., 3rd; Strohl, W.R. Adeno-Associated Virus (AAV) as a Vector for Gene Therapy. BioDrugs 2017, 31, 317–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Yang, H.; Colosi, P. Effect of Genome Size on AAV Vector Packaging. Mol. Ther. 2010, 18, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Grieger, J.C.; Samulski, R.J. Packaging Capacity of Adeno-Associated Virus Serotypes: Impact of Larger Genomes on Infectivity and Postentry Steps. J. Virol. 2005, 79, 9933–9944. [Google Scholar] [CrossRef] [Green Version]
- Bessis, N.; Garcia-Cozar, F.; Boissier, M.-C. Immune responses to gene therapy vectors: Influence on vector function and effector mechanisms. Gene Ther. 2004, 11, S10–S17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McIntosh, J.; Lenting, P.J.; Rosales, C.; Lee, D.; Rabbanian, S.; Raj, D.; Patel, N.; Tuddenham, E.G.D.; Christophe, O.D.; McVey, J.H.; et al. Therapeutic levels of FVIII following a single peripheral vein administration of rAAV vector encoding a novel human factor VIII variant. Blood 2013, 121, 3335–3344. [Google Scholar] [CrossRef] [PubMed]
- Nathwani, A.C.; Tuddenham, E.G.D.; Rangarajan, S.; Rosales, C.; McIntosh, J.; Linch, D.C.; Chowdary, P.; Riddell, A.; Pie, A.J.; Harrington, C.; et al. Adenovirus-Associated Virus Vector–Mediated Gene Transfer in Hemophilia B. N. Engl. J. Med. 2011, 365, 2357–2365. [Google Scholar] [CrossRef]
- Nathwani, A.C.; Reiss, U.M.; Tuddenham, E.G.; Rosales, C.; Chowdary, P.; McIntosh, J.; Della Peruta, M.; Lheriteau, E.; Patel, N.; Raj, D.; et al. Long-Term Safety and Efficacy of Factor IX Gene Therapy in Hemophilia B. N. Engl. J. Med. 2014, 371, 1994–2004. [Google Scholar] [CrossRef] [Green Version]
- Piechnik, M.; Sawamoto, K.; Ohnishi, H.; Kawamoto, N.; Ago, Y.; Tomatsu, S. Evading the AAV Immune Response in Mucopolysaccharidoses. Int. J. Mol. Sci. 2020, 21, 3433. [Google Scholar] [CrossRef] [PubMed]
- Keeler, G.D.; Markusic, D.M.; Hoffman, B.E. Liver induced transgene tolerance with AAV vectors. Cell. Immunol. 2017, 342, 103728. [Google Scholar] [CrossRef] [PubMed]
- Sands, M.S. AAV-Mediated Liver-Directed Gene Therapy. In Adeno-Associated Virus: Methods and Protocols; Snyder, R., Moullier, P., Eds.; Humana Press: Totowa, NJ, USA, 2011; Volume 807, pp. 141–157. [Google Scholar] [CrossRef] [Green Version]
- Sawamoto, K.; Karumuthil-Melethil, S.; Khan, S.; Stapleton, M.; Bruder, J.T.; Danos, O.; Tomatsu, S. Liver-Targeted AAV8 Gene Therapy Ameliorates Skeletal and Cardiovascular Pathology in a Mucopolysaccharidosis IVA Murine Model. Mol. Ther. Methods Clin. Dev. 2020, 18, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Davidoff, A.M.; Ng, C.Y.C.; Zhou, J.; Spence, Y.; Nathwani, A.C. Sex significantly influences transduction of murine liver by recombinant adeno-associated viral vectors through an androgen-dependent pathway. Blood 2003, 102, 480–488. [Google Scholar] [CrossRef] [Green Version]
- Maguire, C.A.; Crommentuijn, M.H.; Mu, D.; Hudry, E.; Serrano-Pozo, A.; Hyman, B.T.; A Tannous, B. Mouse Gender Influences Brain Transduction by Intravascularly Administered AAV9. Mol. Ther. 2013, 21, 1470–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Arizza, V.; Vazzana, M.; Schillaci, D.; Russo, D.; Giaramita, F.T.; Parrinello, N. Gender differences in the immune system activities of sea urchin Paracentrotus lividus. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2012, 164, 447–455. [Google Scholar] [CrossRef] [Green Version]
- Hill-Burns, E.M.; Clark, A.G. X-Linked Variation in Immune Response in Drosophila melanogaster. Genetics 2009, 183, 1477–1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fargallo, J.A.; Martínez-Padilla, J.; Toledano-Díaz, A.; Santiago-Moreno, J.; Dávila, J.A. Sex and testosterone effects on growth, immunity and melanin coloration of nestling Eurasian kestrels. J. Anim. Ecol. 2006, 76, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, J.; Wiesner, A.; Götz, P.; Sauer, K.P. Gender differences and individual variation in the immune system of the scorpionfly Panorpa vulgaris (Insecta: Mecoptera). Dev. Comp. Immunol. 2000, 24, 1–12. [Google Scholar] [CrossRef]
- Mondal, S.; Rai, U. Sexual Dimorphism in Phagocytic Activity of Wall Lizard’s Splenic Macrophages and Its Control by Sex Steroids. Gen. Comp. Endocrinol. 1999, 116, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Rettew, J.A.; Huet-Hudson, Y.M.; Marriott, I. Testosterone Reduces Macrophage Expression in the Mouse of Toll-Like Receptor 4, a Trigger for Inflammation and Innate Immunity. Biol. Reprod. 2008, 78, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, Y.; Ran, S.; Segal, S. Sex-associated differences in the regulation of immune responses controlled by the MHC of the mouse. J. Immunol. 1984, 132, 656–661. [Google Scholar] [PubMed]
- Zuk, M. The Sicker Sex. PLOS Pathog. 2009, 5, e1000267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, G.L.; Martino, A.T.; Aslanidi, G.V.; Jayandharan, G.R.; Srivastava, A.; Herzog, R.W. Innate Immune Responses to AAV Vectors. Front. Microbiol. 2011, 2, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nathwani, A.C.; Cochrane, M.; McIntosh, J.; Ng, C.Y.; Zhou, J.; Gray, J.T.; Davidoff, A.M. Enhancing transduction of the liver by adeno-associated viral vectors. Gene Ther. 2008, 16, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Mackus, W.J.M.; Frakking, F.N.J.; Grummels, A.; Gamadia, L.E.; de Bree, G.J.; Hamann, D.; van Lier, R.A.W.; van Oers, M.H.J. Expansion of CMV-specific CD8+CD45RA+CD27- T cells in B-cell chronic lymphocytic leukemia. Blood 2003, 102, 1057–1063. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.; Zak, R.; Luxton, G.G.; Ritchie, T.C.; Bantel-Schaal, U.; Engelhardt, J.F. Ubiquitination of both Adeno-Associated Virus Type 2 and 5 Capsid Proteins Affects the Transduction Efficiency of Recombinant Vectors. J. Virol. 2002, 76, 2043–2053. [Google Scholar] [CrossRef] [Green Version]
- Fisher, K.J.; Gao, G.P.; Weitzman, M.D.; DeMatteo, R.; Burda, J.F.; Wilson, J.M. Transduction with recombinant adeno-associated virus for gene therapy is limited by leading-strand synthesis. J. Virol. 1996, 70, 520–532. [Google Scholar] [CrossRef] [Green Version]
- Dodge, J.C.; Clarke, J.; Song, A.; Cheng, S.H.; Stewart, G.R.; Dodge, J.C.J.C.; Passini, M.A.; O’Riordan, C.R. Sex and Estrous Cycle Stage Influence the Efficiency of AAV-Mediated Gene Transfer in the Rodent Brain. Mol. Ther. 2005, 11, S192–S193. [Google Scholar] [CrossRef]
- Torcia, M.G.; Nencioni, L.; Clemente, A.M.; Civitelli, L.; Celestino, I.; Limongi, D.; Fadigati, G.; Perissi, E.; Cozzolino, F.; Garaci, E.; et al. Sex Differences in the Response to Viral Infections: TLR8 and TLR9 Ligand Stimulation Induce Higher IL10 Production in Males. PLoS ONE 2012, 7, e39853. [Google Scholar] [CrossRef]
- Klein, S.L. Sex influences immune responses to viruses, and efficacy of prophylaxis and treatments for viral diseases. BioEssays 2012, 34, 1050–1059. [Google Scholar] [CrossRef] [Green Version]
- Kovats, S.; Carreras, E.; Agrawal, H. Sex Steroid Receptors in Immune Cells. In Sex Hormones and Immunity to Infection; Klein, S., Roberts, C., Eds.; Springer: Berlin, Germany, 2010; pp. 53–91. [Google Scholar] [CrossRef]
- Vandamme, C.; Adjali, O.; Mingozzi, F. Unraveling the Complex Story of Immune Responses to AAV Vectors Trial After Tri-al. Hum. Gene Ther. 2017, 28, 161–1074. [Google Scholar] [CrossRef] [PubMed]
- Muenzer, J.; Prada, C.E.; Burton, B.; Lau, H.A.; Ficicioglu, C.; Foo, C.W.P.; Vaidya, S.A.; Whitley, C.B.; Harmatz, P. CHAMPI-ONS: A Phase 1/2 Clinical Trial with Dose Escalation of SB-913 ZFN-Mediated in Vivo Human Genome Editing for Treatment of MPS II (Hunter Syndrome). Mol. Genet. Metab. 2019, 126, S104. [Google Scholar]
- Office USGA. National Institutes of Health: Better Oversight Needed to Help Ensure Continued Progress Including Women in Health Research. 2021. Available online: https://www.gao.gov/products/gao-16-13 (accessed on 6 July 2022).
- Lee, H.; Pak, Y.; Yeo, E.-J.; Kim, Y.S.; Paik, H.Y.; Lee, S.K. It is time to integrate sex as a variable in preclinical and clinical studies. Exp. Mol. Med. 2018, 50, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakiniaeiz, Y.; Cosgrove, K.P.; Potenza, M.N.; Mazure, C.M. Balance of the Sexes: Addressing Sex Differences in Preclinical Re-search. Yale J. Biol. Med. 2016, 89, 255–259. [Google Scholar]
- Sugimoto, C.R.; Ahn, Y.-Y.; Smith, E.; Macaluso, B.; Larivière, V. Factors affecting sex-related reporting in medical research: A cross-disciplinary bibliometric analysis. Lancet 2019, 393, 550–559. [Google Scholar] [CrossRef] [Green Version]
- Tomatsu, S.; Orii, K.O.; Vogler, C.; Nakayama, J.; Levy, B.; Grubb, J.H.; Gutierrez, M.A.; Shim, S.; Yamaguchi, S.; Nishioka, T.; et al. Mouse model of N-acetylgalactosamine-6-sulfate sulfatase deficiency (Galns−/−) produced by targeted disruption of the gene defective in Morquio A disease. Hum. Mol. Genet. 2003, 12, 3349–3358. [Google Scholar] [CrossRef] [Green Version]
- Toietta, G.; Severini, G.M.; Traversari, C.; Tomatsu, S.; Sukegawa, K.; Fukuda, S.; Kondo, N.; Tortora, P.; Bordignon, C. Various Cells Retrovirally Transduced withN-Acetylgalactosoamine-6-Sulfate Sulfatase Correct Morquio Skin FibroblastsIn Vitro. Hum. Gene Ther. 2001, 12, 2007–2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mochizuki, H.; Yoshida, K.; Shibata, Y.; Kimata, K. Tetrasulfated Disaccharide Unit in Heparan Sulfate: Enzymatic formation and tissue distribution. J. Biol. Chem. 2008, 283, 31237–31245. [Google Scholar] [CrossRef] [Green Version]
- Oguma, T.; Tomatsu, S.; Okazaki, O. Analytical method for determination of disaccharides derived from keratan sulfates in human serum and plasma by high-performance liquid chromatography/turbo-ionspray ionization tandem mass spectrometry. Biomed. Chromatogr. 2007, 21, 356–362. [Google Scholar] [CrossRef]
- Oguma, T.; Tomatsu, S.; Montano, A.M.; Okazaki, O. Analytical method for the determination of disaccharides derived from keratan, heparan, and dermatan sulfates in human serum and plasma by high-performance liquid chromatography/turbo ionspray ionization tandem mass spectrometry. Anal. Biochem. 2007, 368, 79–86. [Google Scholar] [CrossRef]
- Kubaski, F.; Shimada, T.; Tomatsu, S.; Yasuda, E.; Mason, R.W.; MacKenzie, W.G.; Shibata, Y.; Giugliani, R.; Yamaguchi, S.; Suzuki, Y.; et al. Chondroitin 6-sulfate as a novel biomarker for mucopolysaccharidosis IVA and VII. Mol. Genet. Metab. 2015, 114, S66. [Google Scholar] [CrossRef] [Green Version]
- Shimada, T.; Tomatsu, S.; Mason, R.W.; Yasuda, E.; MacKenzie, W.G.; Hossain, J.; Shibata, Y.; Montaño, A.M.; Kubaski, F.; Giugliani, R.; et al. Di-sulfated Keratan Sulfate as a Novel Biomarker for Mucopolysaccharidosis II, IVA, and IVB. JIMD Rep. 2015, 21, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubaski, F.; Mason, R.W.; Nakatomi, A.; Shintaku, H.; Xie, L.; van Vlies, N.N.; Church, H.; Giugliani, R.; Kobayashi, H.; Yamaguchi, S.; et al. Newborn screening for mucopolysaccharidoses: A pilot study of measurement of glycosaminoglycans by tandem mass spectrometry. J. Inherit. Metab. Dis. 2016, 40, 151–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomatsu, S.; Orii, K.O.; Vogler, C.; Grubb, J.H.; Snella, E.; A Gutierrez, M.; Dieter, T.; Holden, C.C.; Sukegawa, K.; Orii, T.; et al. Production of MPS VII mouse (Gus(tm(hE540A x mE536A)Sly)) doubly tolerant to human and mouse beta-glucuronidase. Hum. Mol. Genet. 2003, 12, 961–973. [Google Scholar] [CrossRef] [Green Version]
- Tomatsu, S.; Gutierrez, M.; Nishioka, T.; Yamada, M.; Yamada, M.; Tosaka, Y.; Grubb, J.H.; Montaño, A.M.; Vieira, M.B.; Trandafirescu, G.G.; et al. Development of MPS IVA mouse (Galnstm(hC79S·mC76S)slu) tolerant to human N-acetylgalactosamine-6-sulfate sulfatase. Hum. Mol. Genet. 2005, 14, 3321–3335. [Google Scholar] [CrossRef]
- Tomatsu, S.; Montaño, A.M.; Dung, V.C.; Ohashi, A.; Oikawa, H.; Oguma, T.; Orii, T.; Barrera, L.; Sly, W.S. Enhancement of Drug Delivery: Enzyme-replacement Therapy for Murine Morquio A Syndrome. Mol. Ther. 2010, 18, 1094–1102. [Google Scholar] [CrossRef]
- Tomatsu, S.; Montaño, A.M.; Oikawa, H.; Dung, V.C.; Hashimoto, A.; Oguma, T.; Gutiérrez, M.L.; Takahashi, T.; Shimada, T.; Orii, T.; et al. Enzyme replacement therapy in newborn mucopolysaccharidosis IVA mice: Early treatment rescues bone lesions? Mol. Genet. Metab. 2015, 114, 195–202. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piechnik, M.; Amendum, P.C.; Sawamoto, K.; Stapleton, M.; Khan, S.; Fnu, N.; Álvarez, V.; Pachon, A.M.H.; Danos, O.; Bruder, J.T.; et al. Sex Difference Leads to Differential Gene Expression Patterns and Therapeutic Efficacy in Mucopolysaccharidosis IVA Murine Model Receiving AAV8 Gene Therapy. Int. J. Mol. Sci. 2022, 23, 12693. https://doi.org/10.3390/ijms232012693
Piechnik M, Amendum PC, Sawamoto K, Stapleton M, Khan S, Fnu N, Álvarez V, Pachon AMH, Danos O, Bruder JT, et al. Sex Difference Leads to Differential Gene Expression Patterns and Therapeutic Efficacy in Mucopolysaccharidosis IVA Murine Model Receiving AAV8 Gene Therapy. International Journal of Molecular Sciences. 2022; 23(20):12693. https://doi.org/10.3390/ijms232012693
Chicago/Turabian StylePiechnik, Matthew, Paige C. Amendum, Kazuki Sawamoto, Molly Stapleton, Shaukat Khan, Nidhi Fnu, Victor Álvarez, Angelica Maria Herreño Pachon, Olivier Danos, Joseph T. Bruder, and et al. 2022. "Sex Difference Leads to Differential Gene Expression Patterns and Therapeutic Efficacy in Mucopolysaccharidosis IVA Murine Model Receiving AAV8 Gene Therapy" International Journal of Molecular Sciences 23, no. 20: 12693. https://doi.org/10.3390/ijms232012693