


A Role of DNA Methylation within the CYP17A1 Gene in the Association of Genetic and Environmental Risk Factors with Stress-Related Manifestations of Schizophrenia
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Sample Characteristics
2.2. DNAm in Patients and Controls
2.3. Genetic and Environmental Influences on DNAm at VMS
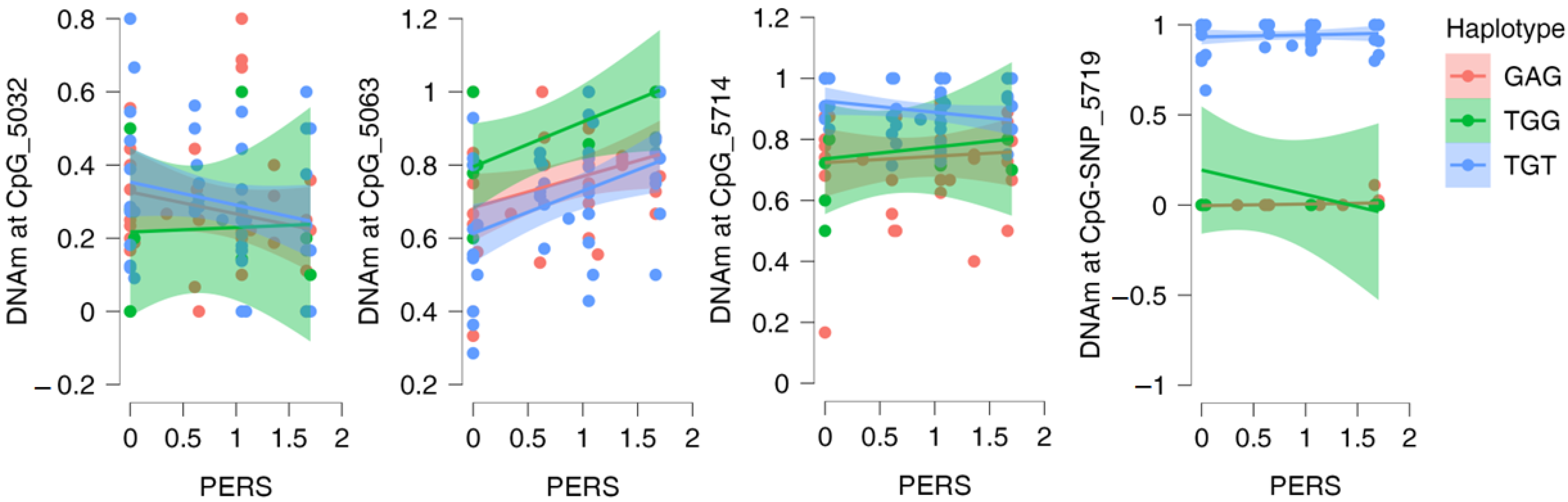
2.4. DNAm and Stress-Related Phenotypes
2.5. AS3MT VNTR and Stress-Related Phenotypes
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. DNAm Analysis
4.3. AS3MT VNTR Genotyping
4.4. Assessment of Environmental Risk Factors
4.5. Assessment of Stress-Related Phenotypes
4.6. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Trubetskoy, V.; Pardiñas, A.F.; Qi, T.; Panagiotaropoulou, G.; Awasthi, S.; Bigdeli, T.B.; Bryois, J.; Chen, C.Y.; Dennison, C.A.; Hall, L.S.; et al. Mapping genomic loci implicates genes and synaptic biology in schizophrenia. Nature 2022, 604, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Pries, L.K.; Lage-Castellanos, A.; Delespaul, P.; Kenis, G.; Luykx, J.J.; Lin, B.D.; Richards, A.L.; Akdede, B.; Binbay, T.; Altinyazar, V.; et al. Estimating Exposome Score for Schizophrenia Using Predictive Modeling Approach in Two Independent Samples: The Results From the EUGEI Study. Schizoph. Bull. 2019, 45, 960–965. [Google Scholar] [CrossRef] [PubMed]
- Green, I.W.; Glausier, J.R. Different Paths to Core Pathology: The Equifinal Model of the Schizophrenia Syndrome. Schizophr. Bull. 2016, 42, 542–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dion, A.; Muñoz, P.T.; Franklin, T.B. Epigenetic mechanisms impacted by chronic stress across the rodent lifespan. Neurobiol. Stress 2022, 17, 100434. [Google Scholar] [CrossRef] [PubMed]
- Perzel Mandell, K.A.; Eagles, N.J.; Wilton, R.; Price, A.J.; Semick, S.A.; Collado-Torres, L.; Ulrich, W.S.; Tao, R.; Han, S.; Szalay, A.S.; et al. Genome-wide sequencing-based identification of methylation quantitative trait loci and their role in schizophrenia risk. Nat. Commun. 2021, 12, 5251. [Google Scholar] [CrossRef] [PubMed]
- Czamara, D.; Eraslan, G.; Page, C.M.; Lahti, J.; Lahti-Pulkkinen, M.; Hämäläinen, E.; Kajantie, E.; Laivuori, H.; Villa, P.M.; Reynolds, R.M.; et al. Integrated analysis of environmental and genetic influences on cord blood DNA methylation in new-borns. Nat. Commun. 2019, 10, 2548. [Google Scholar] [CrossRef] [Green Version]
- Czamara, D.; Tissink, E.; Tuhkanen, J.; Martins, J.; Awaloff, Y.; Drake, A.J.; Khulan, B.; Palotie, A.; Winter, S.M.; Nemeroff, C.B.; et al. Combined effects of genotype and childhood adversity shape variability of DNA methylation across age. Transl. Psychiatry 2021, 11, 88. [Google Scholar] [CrossRef]
- Schizophrenia Working Group of the Psychiatric Genomics Consortium. Biological insights from 108 schizophrenia-associated genetic loci. Nature 2014, 511, 421–427. [Google Scholar] [CrossRef] [Green Version]
- Lotan, A.; Fenckova, M.; Bralten, J.; Alttoa, A.; Dixson, L.; Williams, R.W.; van der Voet, M. Neuroinformatic analyses of common and distinct genetic components associated with major neuropsychiatric disorders. Front. Neurosci. 2014, 8, 331. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Chen, Y.; Lu, J.; Li, W.; Li, X.; Guo, H.; Chen, Q.; Yang, Y.; Xia, H. AS3MT Polymorphism: A Risk Factor for Epilepsy Susceptibility and Adverse Drug Reactions to Valproic Acid and Oxcarbazepine Treatment in Children From South China. Front. Neurosci. 2021, 15, 705297. [Google Scholar] [CrossRef]
- Gerritsen, L.; Milaneschi, Y.; Vinkers, C.H.; van Hemert, A.M.; van Velzen, L.; Schmaal, L.; Penninx, B.W. HPA Axis Genes, and Their Interaction with Childhood Maltreatment, are Related to Cortisol Levels and Stress-Related Phenotypes. Neuropsychopharmacology 2017, 42, 2446–2455. [Google Scholar] [CrossRef] [PubMed]
- Parade, S.H.; Huffhines, L.; Daniels, T.E.; Stroud, L.R.; Nugent, N.R.; Tyrka, A.R. A systematic review of childhood maltreatment and DNA methylation: Candidate gene and epigenome-wide approaches. Transl. Psychiatry 2021, 11, 134. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.T.; Barber, G.P.; Benet-Pagès, A.; Casper, J.; Clawson, H.; Diekhans, M.; Fischer, C.; Gonzalez, J.N.; Hinrichs, A.S.; Lee, C.M.; et al. The UCSC Genome Browser database: 2022 update. Nucleic Acids Res. 2022, 50, D1115–D1122. [Google Scholar] [CrossRef] [PubMed]
- GTEx Consortium Laboratory; Data Analysis &Coordinating Center (LDACC)—Analysis Working Group; Statistical Methods groups—Analysis Working Group; Enhancing GTEx (eGTEx) Groups; NIH Common Fund; NIH/NCI; NIH/NHGRI; NIH/NIMH; NIH/NIDA; Biospecimen Collection Source Site—NDRI. Genetic effects on gene expression across human tissues. Nature 2017, 550, 204–213. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.S.; Huntley, M.H.; Durand, N.C.; Stamenova, E.K.; Bochkov, I.D.; Robinson, J.T.; Sanborn, A.L.; Machol, I.; Omer, A.D.; Lander, E.S.; et al. A 3D map of the human genome at kilobase resolution reveals principles of chromatin looping. Cell 2014, 159, 1665–1680. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Sebra, R.; Pullman, B.S.; Qiao, W.; Peter, I.; Desnick, R.J.; Geyer, C.R.; DeCoteau, J.F.; Scott, S.A. Quantitative and multiplexed DNA methylation analysis using long-read single-molecule real-time bisulfite sequencing (SMRT-BS). BMC Genom. 2015, 16, 350. [Google Scholar] [CrossRef] [Green Version]
- Pechtel, P.; Pizzagalli, D.A. Effects of early life stress on cognitive and affective function: An integrated review of human literature. Psychopharmacology 2011, 214, 55–70. [Google Scholar] [CrossRef] [Green Version]
- Slavich, G.M.; Auerbach, R.P. Stress and its sequelae: Depression, suicide, inflammation, and physical illness. In APA Handbook of Psychopathology: Psychopathology, 1st ed.; Butcher, J.N., Hooley, J.M., Eds.; American Psychological Association: Washington, DC, USA, 2018; Volume 1, pp. 375–402. [Google Scholar]
- Wood, T.C.; Salavagionne, O.E.; Mukherjee, B.; Wang, L.; Klumpp, A.F.; Thomae, B.A.; Eckloff, B.W.; Schaid, D.J.; Wieben, E.D.; Weinshilboum, R.M. Human arsenic methyltransferase (AS3MT) pharmacogenetics: Gene resequencing and functional genomics studies. J. Biol. Chem. 2006, 281, 7364–7373. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Rubio, P.; Meza-Montenegro, M.M.; Cantu-Soto, E.; Klimecki, W.T. Genetic association between intronic variants in AS3MT and arsenic methylation efficiency is focused on a large linkage disequilibrium cluster in chromosome 10. J. Appl. Toxicol. 2010, 30, 260–270. [Google Scholar] [CrossRef] [Green Version]
- Engström, K.S.; Hossain, M.B.; Lauss, M.; Ahmed, S.; Raqib, R.; Vahter, M.; Broberg, K. Efficient arsenic metabolism--the AS3MT haplotype is associated with DNA methylation and expression of multiple genes around AS3MT. PLoS ONE 2013, 8, 53732. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Jaffe, A.E.; Straub, R.E.; Tao, R.; Shin, J.H.; Wang, Y.; Chen, Q.; Li, C.; Jia, Y.; Ohi, K.; et al. A human-specific AS3MT isoform and BORCS7 are molecular risk factors in the 10q24.32 schizophrenia-associated locus. Nat. Med. 2016, 22, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Yang, Z.H.; Li, H.J.; Xiao, X.; Li, M.; Chang, H. A Human-Specific Schizophrenia Risk Tandem Repeat Affects Alternative Splicing of a Human-Unique Isoform AS3MTd2d3 and Mushroom Dendritic Spine Density. Schizophr. Bull. 2021, 47, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Zhang, Q.; Chen, X.; Li, Y.; Li, X.; Du, B.; Deng, X.; Ji, F.; Wang, C.; Xiang, Y.T.; et al. The VNTR of the AS3MT gene is associated with brain activations during a memory span task and their training-induced plasticity. Psychol. Med. 2021, 51, 1927–1932. [Google Scholar] [CrossRef] [Green Version]
- Korovaitseva, G.I.; Gabaeva, M.V.; Yunilainen, O.A.; Golimbet, V.E. Effect of VNTR Polymorphism of the AS3MT Gene and Obstetrical Complications on the Severity of Schizophrenia. Bull. Exp. Biol. Med. 2019, 168, 84–86. [Google Scholar] [CrossRef]
- Machiela, M.J.; Chanock, S.J. LDlink: A web-based application for exploring population-specific haplotype structure and linking correlated alleles of possible functional variants. Bioinformatics 2015, 31, 3555–3557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fong, M.C.; Law, T.S.; Ma, M.K.; Hui, N.Y.; Wang, W.S. Can inhibition deficit hypothesis account for age-related differences in semantic fluency? Converging evidence from Stroop color and word test and an ERP flanker task. Brain Lang. 2021, 218, 104952. [Google Scholar] [CrossRef] [PubMed]
- Huan, T.; Joehanes, R.; Song, C.; Peng, F.; Guo, Y.; Mendelson, M.; Yao, C.; Liu, C.; Ma, J.; Richard, M.; et al. Genome-wide identification of DNA methylation QTLs in whole blood highlights pathways for cardiovascular disease. Nat. Commun. 2019, 10, 4267. [Google Scholar] [CrossRef] [Green Version]
- Savage, J.E.; Jansen, P.R.; Stringer, S.; Watanabe, K.; Bryois, J.; de Leeuw, C.A.; Nagel, M.; Awasthi, S.; Barr, P.B.; Coleman, J.; et al. Genome-wide association meta-analysis in 269,867 individuals identifies new genetic and functional links to intelligence. Nat. Genet. 2018, 50, 912–919. [Google Scholar] [CrossRef] [Green Version]
- Donohoe, G.; Morris, D.W.; Corvin, A. The psychosis susceptibility gene ZNF804A: Associations, functions, and phenotypes. Schizophr. Bull. 2010, 36, 904–909. [Google Scholar] [CrossRef] [Green Version]
- Van den Oord, E.J.; Clark, S.L.; Xie, L.Y.; Shabalin, A.A.; Dozmorov, M.G.; Kumar, G.; Swedish Schizophrenia Consortium; Vladimirov, V.I.; Magnusson, P.K.; Aberg, K.A. A Whole Methylome CpG-SNP Association Study of Psychosis in Blood and Brain Tissue. Schizophr. Bull. 2016, 42, 1018–1026. [Google Scholar] [CrossRef]
- Alfimova, M.; Kondratyev, N.; Golov, A.; Golimbet, V. Relationship between Alzheimer’s disease-associated SNPs within the CLU gene, local DNA methylation and episodic verbal memory in healthy and schizophrenia subjects. Psychiatry Res. 2019, 272, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Kondratyev, N.; Golov, A.; Alfimova, M.; Lezheiko, T.; Golimbet, V. Prediction of smoking by multiplex bisulfite PCR with long amplicons considering allele-specific effects on DNA methylation. Clin. Epigenetics 2018, 10, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfimova, M.; Kondratyev, N.; Golov, A.; Golimbet, V. Profiling haplotype specific CpG and CpH methylation within a schizophrenia GWAS locus on chromosome 14 in schizophrenia and healthy subjects. Sci. Rep. 2020, 10, 4704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, P.; Zhang, X.; Huang, C.C.; Jafari, N.; Kibbe, W.A.; Hou, L.; Lin, S.M. Comparison of Beta-value and M-value methods for quantifying methylation levels by microarray analysis. BMC Bioinform. 2010, 11, 587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afifi, T.O.; Salmon, S.; Garcés, I.; Struck, S.; Fortier, J.; Taillieu, T.; Stewart-Tufescu, A.; Asmundson, G.; Sareen, J.; MacMillan, H.L. Confirmatory factor analysis of adverse childhood experiences (ACEs) among a community-based sample of parents and adolescents. BMC Pediatr. 2020, 20, 178. [Google Scholar] [CrossRef] [Green Version]
- Radua, J.; Ramella-Cravaro, V.; Ioannidis, J.; Reichenberg, A.; Phiphopthatsanee, N.; Amir, T.; Yenn Thoo, H.; Oliver, D.; Davies, C.; Morgan, C.; et al. What causes psychosis? An umbrella review of risk and protective factors. World Psychiatry 2018, 17, 49–66. [Google Scholar] [CrossRef] [Green Version]
- Golden, C.J. Stroop Color and Word Test; Stoelting Company: Wood Dale, IL, USA, 1978. [Google Scholar]
- Kay, S.R.; Fiszbein, A.; Opler, L.A. The positive and negative syndrome scale (PANSS) for schizophrenia. Schizophr. Bull. 1987, 13, 261–276. [Google Scholar] [CrossRef]
- JASP Team. JASP (Version 0.16) (Computer Software). 2021. Available online: https://jasp-stats.org/ (accessed on 1 April 2022).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | DNAm Sample | VNTR Sample | ||
|---|---|---|---|---|
| Patients | Controls | Patients | Controls | |
| n | 66 | 63 | 304 | 466 |
| Sex (% women) | 50 | 52 | 50 | 59 * |
| Age (years) | 27.49 (6.76) | 26.77 (6.42) | 28.76 (8.80) | 28.45 (8.94) |
| Age range | 18–45 | 18–45 | 16–63 | 17–60 |
| Smokers (%) | 30 | 52 * | - | - |
| Education (% tertiary) | 71 | 89 * | 63 | 83 ** |
| WM (T-scores) | 39.16 (11.91) | 50.39 (9.50) ** | 38.21 (11.91) | 49.90 (11.82) ** |
| VF (T-scores) | 39.46 (10.21) | 50.17 (9.02) ** | 38.06 (10.91) | 50.07 (10.21) ** |
| EVM (T-scores) | 37.03 (12.61) | 50.64 (9.34) ** | 35.22 (12.30) | 47.86 (11.19) ** |
| Cognitive flexibility (T-scores) | 37.55 (11.00) | 49.99 (9.13) ** | 35.55 (11.73) | 49.41 (9.53) ** |
| Cognitive Inhibition (T-scores) | 40.45(9.55) | 50.14 (9.73) ** | 39.82 (10.51) | 47.71 (11.09) ** |
| Diagnosis (% F20) | 82 | - | 80 | - |
| Illness duration | 5.95 (6.23) | 6.19 (6.89) | - | |
| PANSS P | 27.38 (8.66) | 25.85(8.94) | - | |
| PANSS N | 20.36 (5.89) | 19.99 (7.40) | - | |
| PANSS G | 31.52 (12.06) | 30.36 (12.57) | - | |
| PANSS G6 ≥ 4 (n) | 3 | - | 12 | - |
| SIB (%) | 22 | - | 26 | - |
| SOB (% winter birth) | 23 | - | 25 | - |
| OC (%) | 36 | - | 35 | - |
| ACE (%) | 55 | - | 46 | - |
| PERS | 0.81 (0.63) | - | 0.71 (0.59) | - |
| Group | CpG_5032 | CpG_5063 | CpG_5714 | CpG-SNP_5719 |
|---|---|---|---|---|
| Patients | 0.28 (0.19) | 0.74 (0.17) | 0.82 (0.16) | 0.49 (0.48) |
| Controls | 0.25 (0.18) | 0.76 (0.16) | 0.76 (0.21) | 0.53 (0.48) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alfimova, M.; Kondratyev, N.; Korovaitseva, G.; Lezheiko, T.; Plakunova, V.; Gabaeva, M.; Golimbet, V. A Role of DNA Methylation within the CYP17A1 Gene in the Association of Genetic and Environmental Risk Factors with Stress-Related Manifestations of Schizophrenia. Int. J. Mol. Sci. 2022, 23, 12629. https://doi.org/10.3390/ijms232012629
Alfimova M, Kondratyev N, Korovaitseva G, Lezheiko T, Plakunova V, Gabaeva M, Golimbet V. A Role of DNA Methylation within the CYP17A1 Gene in the Association of Genetic and Environmental Risk Factors with Stress-Related Manifestations of Schizophrenia. International Journal of Molecular Sciences. 2022; 23(20):12629. https://doi.org/10.3390/ijms232012629
Chicago/Turabian StyleAlfimova, Margarita, Nikolay Kondratyev, Galina Korovaitseva, Tatyana Lezheiko, Victoria Plakunova, Marina Gabaeva, and Vera Golimbet. 2022. "A Role of DNA Methylation within the CYP17A1 Gene in the Association of Genetic and Environmental Risk Factors with Stress-Related Manifestations of Schizophrenia" International Journal of Molecular Sciences 23, no. 20: 12629. https://doi.org/10.3390/ijms232012629