
Individual Variabilities in Adipose Stem Cell Proliferation, Gene Expression and Responses to Lipopolysaccharide Stimulation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Preliminary Characterization of Adipose Stem Cells
2.2. Comparison of Cell Proliferative Capacity between ASCs
2.3. Comparison of Adipogenic Differentiation Potential between ASCs
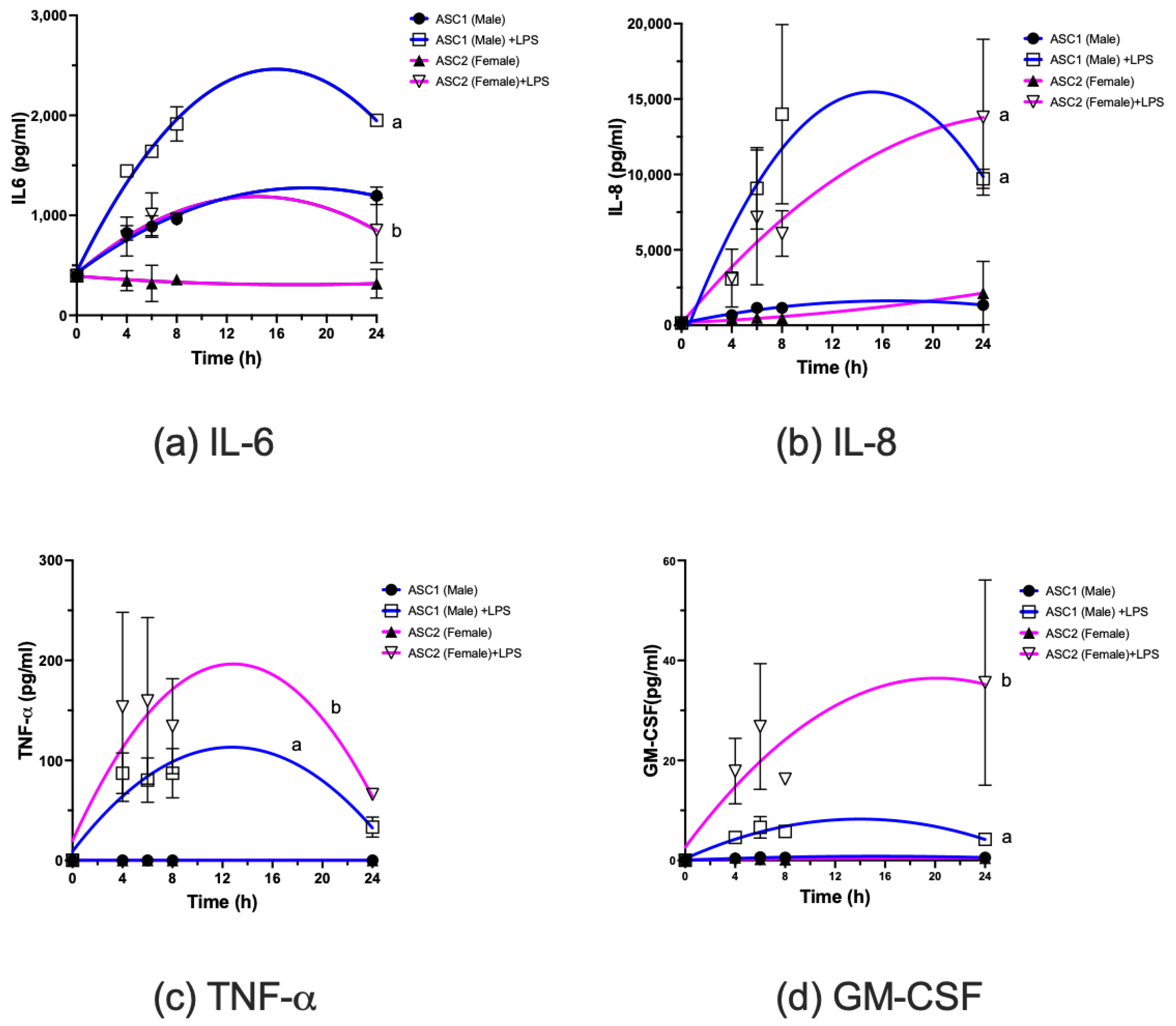
2.4. Comparison of LPS-Induced Changes in Cytokine mRNA and Protein Expression in ASCs
2.5. Effects of ASC-Derived Conditioned Media on Human THP-1 Cell Differentiation
3. Discussion
4. Materials and Methods
4.1. Chemical and Reagents
4.2. Cell Culture
4.3. Cytokines/Chemokines Analysis
4.4. Cell Proliferation Assay
4.5. Oil Red O Staining
4.6. Real-Time PCR Analysis of Gene Expression
4.7. Conditioned Media Treatment of Human THP-1 Cells
4.8. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ginsburg, G.S.; Phillips, K.A. Precision Medicine: From Science to Value. Health Aff. (Proj. Hope) 2018, 37, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Dugger, S.; Platt, A.; Goldstein, D. Drug development in the era of precision medicine. Nat. Rev. Drug Discov. 2018, 17, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Goetz, L.H.; Schork, N.J. Personalized medicine: Motivation, challenges, and progress. Fertil. Steril. 2018, 109, 952–963. [Google Scholar] [CrossRef] [PubMed]
- Tsimberidou, A.M.; Fountzilas, E.; Nikanjam, M.; Kurzrock, R. Review of precision cancer medicine: Evolution of the treatment paradigm. Cancer Treat. Rev. 2020, 86, 102019. [Google Scholar] [CrossRef]
- Bozdağ, S.C.; Yüksel, M.K.; Demirer, T. Adult Stem Cells and Medicine. Adv. Exp. Med. Biol. 2018, 1079, 17–36. [Google Scholar] [CrossRef]
- Venkei, Z.G.; Yamashita, Y.M. Emerging mechanisms of asymmetric stem cell division. J. Cell Biol. 2018, 217, 3785–3795. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Ye, J.; Yuan, F.Z.; Zhang, J.Y.; Chen, Y.R.; Fan, B.S.; Jiang, D.; Jiang, W.B.; Wang, X.; Yu, J.K. Advances of Stem Cell-Laden Hydrogels with Biomimetic Microenvironment for Osteochondral Repair. Front. Bioeng. Biotechnol. 2020, 8, 247. [Google Scholar] [CrossRef] [Green Version]
- Rana, D.; Zreiqat, H.; Benkirane-Jessel, N.; Ramakrishna, S.; Ramalingam, M. Development of decellularized scaffolds for stem cell-driven tissue engineering. J. Tissue Eng. Regen. Med. 2017, 11, 942–965. [Google Scholar] [CrossRef]
- Priya, N.; Sarcar, S.; Majumdar, A.S.; SundarRaj, S. Explant culture: A simple, reproducible, efficient and economic technique for isolation of mesenchymal stromal cells from human adipose tissue and lipoaspirate. J. Tissue Eng. Regen. Med. 2014, 8, 706–716. [Google Scholar] [CrossRef]
- Alstrup, T.; Eijken, M.; Bohn, A.B.; Møller, B.; Damsgaard, T.E. Isolation of Adipose Tissue-Derived Stem Cells: Enzymatic Digestion in Combination with Mechanical Distortion to Increase Adipose Tissue-Derived Stem Cell Yield from Human Aspirated Fat. Curr. Protoc. Stem Cell Biol. 2019, 48, e68. [Google Scholar] [CrossRef]
- Schneider, S.; Unger, M.; van Griensven, M.; Balmayor, E.R. Adipose-derived mesenchymal stem cells from liposuction and resected fat are feasible sources for regenerative medicine. Eur. J. Med. Res. 2017, 22, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estes, B.T.; Diekman, B.O.; Gimble, J.M.; Guilak, F. Isolation of adipose-derived stem cells and their induction to a chondrogenic phenotype. Nat. Protoc. 2010, 5, 1294–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chouchani, E.T.; Kajimura, S. Metabolic adaptation and maladaptation in adipose tissue. Nat. Metab. 2019, 1, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Liu, Y.; Sun, Y.; Wang, B.; Xiong, Y.; Lin, W.; Wei, Q.; Wang, H.; He, W.; Wang, B.; et al. Tissue source determines the differentiation potentials of mesenchymal stem cells: A comparative study of human mesenchymal stem cells from bone marrow and adipose tissue. Stem Cell Res. Ther. 2017, 8, 275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, E.M.; Esper, R.M.; Sen, A.; Simon, B.R.; Aslam, M.N.; Jiang, Y.; Dame, M.K.; McClintock, S.D.; Colacino, J.A.; Djuric, Z.; et al. Dietary polyunsaturated fatty acids modulate adipose secretome and is associated with changes in mammary epithelial stem cell self-renewal. J. Nutr. Biochem. 2019, 71, 45–53. [Google Scholar] [CrossRef]
- Meissburger, B.; Perdikari, A.; Moest, H.; Müller, S.; Geiger, M.; Wolfrum, C. Regulation of adipogenesis by paracrine factors from adipose stromal-vascular fraction—A link to fat depot-specific differences. Biochim. Biophys. Acta 2016, 1861 Pt A, 1121–1131. [Google Scholar] [CrossRef] [Green Version]
- Cho, D.S.; Lee, B.; Doles, J.D. Refining the adipose progenitor cell landscape in healthy and obese visceral adipose tissue using single-cell gene expression profiling. Life Sci. Alliance 2019, 2, e201900561. [Google Scholar] [CrossRef]
- Silva, K.R.; Côrtes, I.; Liechocki, S.; Carneiro, J.R.; Souza, A.A.; Borojevic, R.; Maya-Monteiro, C.M.; Baptista, L.S. Characterization of stromal vascular fraction and adipose stem cells from subcutaneous, preperitoneal and visceral morbidly obese human adipose tissue depots. PLoS ONE 2017, 12, e0174115. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.Y.; Ma, S.; Eirin, A.; Woollard, J.R.; Hickson, L.J.; Sun, D.; Lerman, A.; Lerman, L.O. Functional Plasticity of Adipose-Derived Stromal Cells during Development of Obesity. Stem Cells Trans. Med. 2016, 5, 893–900. [Google Scholar] [CrossRef]
- Pachón-Peña, G.; Serena, C.; Ejarque, M.; Petriz, J.; Duran, X.; Oliva-Olivera, W.; Simó, R.; Tinahones, F.J.; Fernández-Veledo, S.; Vendrell, J. Obesity Determines the Immunophenotypic Profile and Functional Characteristics of Human Mesenchymal Stem Cells from Adipose Tissue. Stem Cells Trans. Med. 2016, 5, 464–475. [Google Scholar] [CrossRef]
- Hofmann, E.; Soppert, J.; Ruhl, T.; Gousopoulos, E.; Gerra, S.; Storti, G.; Tian, Y.; Brandhofer, M.; Schweizer, R.; Song, S.Y.; et al. The Role of Macrophage Migration Inhibitory Factor in Adipose-Derived Stem Cells Under Hypoxia. Front. Physiol. 2021, 12, 638448. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Niu, P.; Zhao, Y.; Cheng, Y.; Chen, W.; Lin, L.; Lu, J.; Cheng, X.; Xu, Z. Impact of miR-223-3p and miR-2909 on inflammatory factors IL-6, IL-1ß, and TNF-α, and the TLR4/TLR2/NF-κB/STAT3 signaling pathway induced by lipopolysaccharide in human adipose stem cells. PLoS ONE 2019, 14, e0212063. [Google Scholar] [CrossRef] [Green Version]
- Ting, H.K.; Chen, C.L.; Meng, E.; Cherng, J.H.; Chang, S.J.; Kao, C.C.; Yang, M.H.; Leung, F.S.; Wu, S.T. Inflammatory Regulation by TNF-α-Activated Adipose-Derived Stem Cells in the Human Bladder Cancer Microenvironment. Int. J. Mol. Sci. 2021, 22, 3987. [Google Scholar] [CrossRef]
- D’Esposito, V.; Ambrosio, M.R.; Liguoro, D.; Perruolo, G.; Lecce, M.; Cabaro, S.; Aprile, M.; Marino, A.; Pilone, V.; Forestieri, P.; et al. In severe obesity, subcutaneous adipose tissue cell-derived cytokines are early markers of impaired glucose tolerance and are modulated by quercetin. Int. J. Obes. 2021, 45, 1811–1820. [Google Scholar] [CrossRef] [PubMed]
- Valverde, M.; Sánchez-Brito, A. Sustained Activation of TNFα-Induced DNA Damage Response in Newly Differentiated Adipocytes. Int. J. Mol. Sci. 2021, 22, 10548. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; El-Sabbagh, A.S.; Lukas, B.E.; Tanneberger, S.J.; Jiang, Y. Adipose stem cells in obesity: Challenges and opportunities. Biosci. Rep. 2020, 40, BSR20194076. [Google Scholar] [CrossRef] [PubMed]
- Ejarque, M.; Ceperuelo-Mallafré, V.; Serena, C.; Maymo-Masip, E.; Duran, X.; Díaz-Ramos, A.; Millan-Scheiding, M.; Núñez-Álvarez, Y.; Núñez-Roa, C.; Gama, P.; et al. Adipose tissue mitochondrial dysfunction in human obesity is linked to a specific DNA methylation signature in adipose-derived stem cells. Int. J. Obes. 2019, 43, 1256–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frazier, T.; Lee, S.; Bowles, A.; Semon, J.; Bunnell, B.; Wu, X.; Gimble, J. Gender and age-related cell compositional differences in C57BL/6 murine adipose tissue stromal vascular fraction. Adipocyte 2018, 7, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Lynch, K.; Pei, M. Age associated communication between cells and matrix: A potential impact on stem cell-based tissue regeneration strategies. Organogenesis 2014, 10, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Quail, D.F.; Dannenberg, A.J. The obese adipose tissue microenvironment in cancer development and progression. Nat. Rev. Endocrinol. 2019, 15, 139–154. [Google Scholar] [CrossRef]
- Lyes, M.A.; Payne, S.; Ferrell, P.; Pizzo, S.V.; Hollenbeck, S.T.; Bachelder, R.E. Adipose stem cell crosstalk with chemo-residual breast cancer cells: Implications for tumor recurrence. Breast Cancer Res. Treat. 2019, 174, 413–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muehlberg, F.L.; Song, Y.H.; Krohn, A.; Pinilla, S.P.; Droll, L.H.; Leng, X.; Seidensticker, M.; Ricke, J.; Altman, A.M.; Devarajan, E.; et al. Tissue-resident stem cells promote breast cancer growth and metastasis. Carcinogenesis 2009, 30, 589–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindstad, T.; Qu, S.; Sikkeland, J.; Jin, Y.; Kristian, A.; Mælandsmo, G.M.; Collas, P.; Saatcioglu, F. STAMP2 is required for human adipose-derived stem cell differentiation and adipocyte-facilitated prostate cancer growth in vivo. Oncotarget 2016, 8, 91817–91827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.H.; Wei, H.J.; Peng, B.Y.; Chou, H.H.; Chen, W.H.; Liu, H.Y.; Deng, W.P. Adipose-Derived Stem Cells Enhance Cancer Stem Cell Property and Tumor Formation Capacity in Lewis Lung Carcinoma Cells Through an Interleukin-6 Paracrine Circuit. Stem Cells Dev. 2016, 25, 1833–1842. [Google Scholar] [CrossRef]
- Zakaria, N.; Yahaya, B.H. Adipose-Derived Mesenchymal Stem Cells Promote Growth and Migration of Lung Adenocarcinoma Cancer Cells. Adv. Exp. Med. Biol. 2020, 1292, 83–95. [Google Scholar] [CrossRef]
- Chen, Y.; He, Y.; Wang, X.; Lu, F.; Gao, J. Adipose-derived mesenchymal stem cells exhibit tumor tropism and promote tumorsphere formation of breast cancer cells. Oncol. Rep. 2019, 41, 2126–2136. [Google Scholar] [CrossRef] [Green Version]
- Benaiges, E.; Ceperuelo-Mallafré, V.; Madeira, A.; Bosch, R.; Núñez-Roa, C.; Ejarque, M.; Maymó-Masip, E.; Huber-Ruano, I.; Lejeune, M.; Vendrell, J.; et al. Survivin drives tumor-associated macrophage reprogramming: A novel mechanism with potential impact for obesity. Cell. Oncol. 2021, 44, 777–792. [Google Scholar] [CrossRef]
- Scioli, M.G.; Storti, G.; D’Amico, F.; Gentile, P.; Kim, B.S.; Cervelli, V.; Orlandi, A. Adipose-Derived Stem Cells in Cancer Progression: New Perspectives and Opportunities. Int. J. Mol. Sci. 2019, 20, 3296. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.R.; Kim, E.K.; Kim, H.; Claycombe, K.J. Obesity-associated mouse adipose stem cell secretion of monocyte chemotactic protein-1. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E1153–E1158. [Google Scholar] [CrossRef] [Green Version]
- Rios, F.J.; Touyz, R.M.; Montezano, A.C. Isolation and Differentiation of Human Macrophages. Methods Mol. Biol. 2017, 1527, 311–320. [Google Scholar] [CrossRef]
- Jo, J.; Gavrilova, O.; Pack, S.; Jou, W.; Mullen, S.; Sumner, A.E.; Cushman, S.W.; Periwal, V. Hypertrophy and/or Hyperplasia: Dynamics of Adipose Tissue Growth. PLoS Comp. Biol. 2009, 5, e1000324. [Google Scholar] [CrossRef] [PubMed]
- Chomarat, P.; Banchereau, J.; Davoust, J.; Palucka, A.K. IL-6 switches the differentiation of monocytes from dendritic cells to macrophages. Nat. Immunol. 2000, 1, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Menetrier-Caux, C.; Montmain, G.; Dieu, M.C.; Bain, C.; Favrot, M.C.; Caux, C.; Blay, J.Y. Inhibition of the differentiation of dendritic cells from CD34+ progenitors by tumor cells: Role of interleukin-6 and macrophage colony-stimulating factor. Blood 1998, 92, 4778–4791. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, R.; Cheung, A.S.; Pang, K.; Saffery, R.; Novakovic, B. Sexual Dimorphism in Innate Immunity: The Role of Sex Hormones and Epigenetics. Front. Immunol. 2021, 11, 604000. [Google Scholar] [CrossRef] [PubMed]
- Skehan, P.; Storeng, R.; Scudiero, D.; Monks, A.; McMahon, J.; Vistica, D.; Warren, J.T.; Bokesch, H.; Kenney, S.; Boyd, M.R. New colorimetric cytotoxicity assay for anticancer-drug screening. J. Nat. Cancer Inst. 1990, 82, 1107–1112. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yasmeen, R.; Pham, Q.; Fukagawa, N.K.; Wang, T.T.Y. Individual Variabilities in Adipose Stem Cell Proliferation, Gene Expression and Responses to Lipopolysaccharide Stimulation. Int. J. Mol. Sci. 2022, 23, 12534. https://doi.org/10.3390/ijms232012534
Yasmeen R, Pham Q, Fukagawa NK, Wang TTY. Individual Variabilities in Adipose Stem Cell Proliferation, Gene Expression and Responses to Lipopolysaccharide Stimulation. International Journal of Molecular Sciences. 2022; 23(20):12534. https://doi.org/10.3390/ijms232012534
Chicago/Turabian StyleYasmeen, Rumana, Quynhchi Pham, Naomi K. Fukagawa, and Thomas T. Y. Wang. 2022. "Individual Variabilities in Adipose Stem Cell Proliferation, Gene Expression and Responses to Lipopolysaccharide Stimulation" International Journal of Molecular Sciences 23, no. 20: 12534. https://doi.org/10.3390/ijms232012534