
Identification of the NAC Transcription Factors and Their Function in ABA and Salinity Response in Nelumbo nucifera
Abstract
:1. Introduction
2. Results
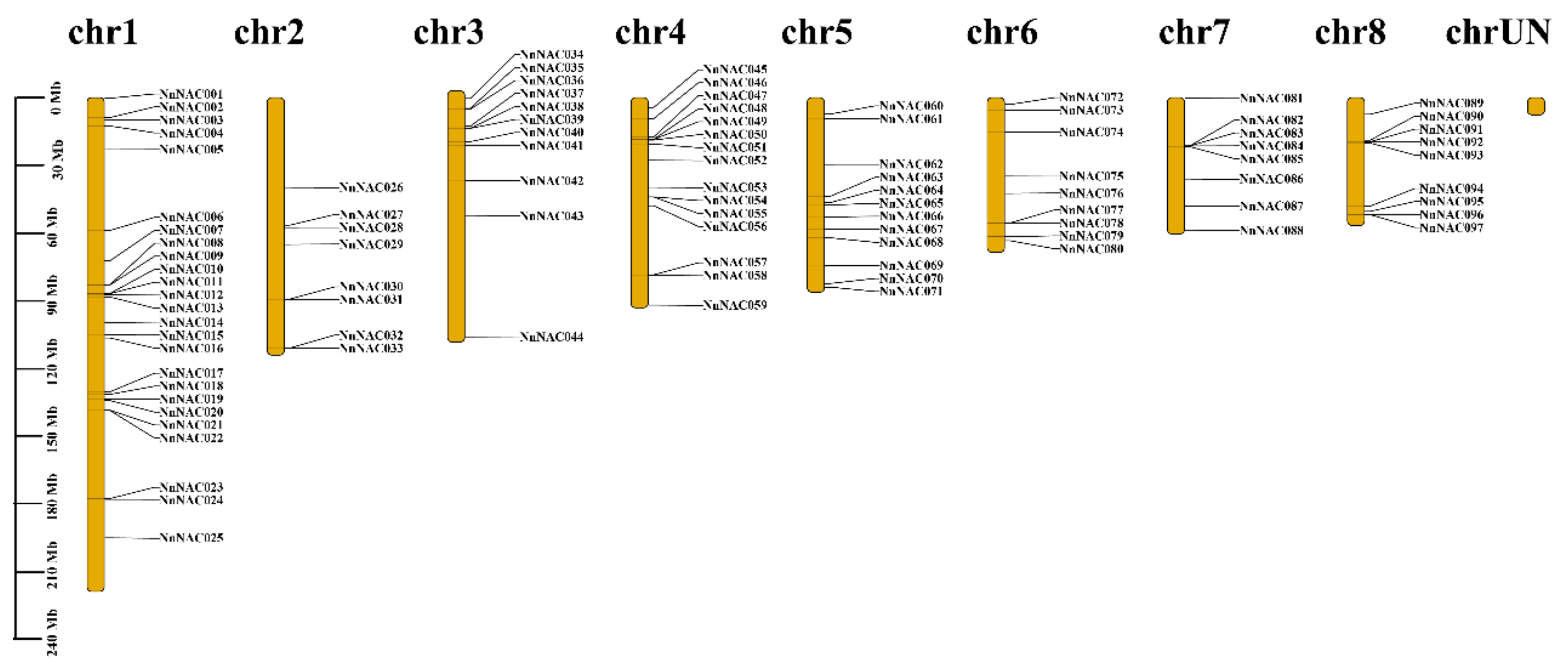
2.1. Identification of NAC Transcription Factors in Nelumbo nucifera
2.2. Phylogenetic Analysis and Classification of NAC Genes
2.3. Gene Structure and Motif Composition of the NAC Gene Family
2.4. Cis-Acting Elements and Synteny Analysis of NAC Gene Promotors
2.5. Expression Patterns of NAC Genes in Different Plant Tissues
2.6. qRT-PCR Analysis of the NAC Genes
2.7. Subcellular Localization of NAC Proteins
2.8. The Transcriptional Activation Capabilities Analysis of NAC Proteins
3. Discussion
4. Materials and Methods
4.1. The Identification of NAC Transcription Factor Members
4.2. Basic Information and Phylogenetic Analyses
4.3. Synteny and Gene Structure Analyses
4.4. Microarray Analysis of NAC Gene Expression Patterns
4.5. Subcellular Localization Analysis
4.6. Plant Materials and Treatments
4.7. Quantitative Real-Time PCR (qRT-PCR)
4.8. The Transcriptional Activation Assays
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, W.; Stewart, C.N. Plant synthetic promoters and transcription factors. Curr. Opin. Biotechnol. 2016, 37, 36–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, A.N.; Ernst, H.A.; Leggio, L.L.; Skriver, K. NAC transcription factors: Structurally distinct, functionally diverse. Trends Plant Sci. 2005, 10, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Munir, N.; Yukun, C.; Xiaohui, C.; Nawaz, M.A.; Iftikhar, J.; Rizwan, H.M.; Xu, S.; Yuling, L.; Xuhan, X.; Zhongxiong, L. Genome-wide identification and comprehensive analyses of NAC transcription factor gene family and expression patterns during somatic embryogenesis in Dimocarpus longan Lour. Plant Physiol. Biochem. 2020, 157, 169–184. [Google Scholar] [CrossRef]
- Ernst, H.A.; Olsen, A.N.; Larsen, S.; Lo Leggio, L. Structure of the conserved domain of ANAC, a member of the NAC family of transcription factors. EMBO Rep. 2004, 5, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Kjaersgaard, T.; Jensen, M.K.; Christiansen, M.W.; Gregersen, P.; Kragelund, B.B.; Skriver, K. Senescence-associated barley NAC (NAM, ATAF1,2, CUC) transcription factor interacts with radical-induced cell death 1 through a disordered regulatory domain. J. Biol. Chem. 2011, 286, 35418–35429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bengoa Luoni, S.A.; Cenci, A.; Moschen, S.; Nicosia, S.; Radonic, L.M.; Sabio, Y.G.J.V.; Langlade, N.B.; Vile, D.; Rovere, C.V.; Fernandez, P. Genome-wide and comparative phylogenetic analysis of senescence-associated NAC transcription factors in sunflower (Helianthus annuus). BMC Genom. 2021, 22, 893. [Google Scholar] [CrossRef] [PubMed]
- Li, P.X.; Peng, Z.Y.; Xu, P.L.; Tang, G.Y.; Ma, C.L.; Zhu, J.Q.; Shan, L.; Wan, S.B. Genome-Wide Identification of NAC Transcription Factors and Their Functional Prediction of Abiotic Stress Response in Peanut. Front. Genet. 2021, 12, 630292. [Google Scholar] [CrossRef] [PubMed]
- Nie, G.; Yang, Z.; He, J.; Liu, A.; Chen, J.; Wang, S.; Wang, X.; Feng, G.; Li, D.; Peng, Y.; et al. Genome-Wide Investigation of the NAC Transcription Factor Family in Miscanthus sinensis and Expression Analysis Under Various Abiotic Stress. Front. Plant Sci. 2021, 12, 766550. [Google Scholar] [CrossRef] [PubMed]
- Duan, A.Q.; Yang, X.L.; Feng, K.; Liu, J.X.; Xu, Z.S.; Xiong, A.S. Genome-wide analysis of NAC transcription factors and their response to abiotic stress in celery (Apium graveolens L.). Comput. Biol. Chem. 2020, 84, 107186. [Google Scholar] [CrossRef]
- Diao, W.; Snyder, J.C.; Wang, S.; Liu, J.; Pan, B.; Guo, G.; Ge, W.; Dawood, M. Genome-Wide Analyses of the NAC Transcription Factor Gene Family in Pepper (Capsicum annuum L.): Chromosome Location, Phylogeny, Structure, Expression Patterns, Cis-Elements in the Promoter, and Interaction Network. Int. J. Mol. Sci. 2018, 19, 1028. [Google Scholar] [CrossRef]
- Singh, A.K.; Sharma, V.; Pal, A.K.; Acharya, V.; Ahuja, P.S. Genome-wide organization and expression profiling of the NAC transcription factor family in potato (Solanum tuberosum L.). DNA Res. 2013, 20, 403–423. [Google Scholar] [CrossRef] [Green Version]
- Le, D.T.; Nishiyama, R.; Watanabe, Y.; Mochida, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.S. Genome-wide survey and expression analysis of the plant-specific NAC transcription factor family in soybean during development and dehydration stress. DNA Res. 2011, 18, 263–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, C.; Wu, M.; Zhou, Y.; Gong, Z.; Yu, W.; Zhang, Y.; Yang, Z. NAC-mediated membrane lipid remodeling negatively regulates fruit cold tolerance. Hortic. Res. 2022, 9, uhac039. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Kang, X.; Liu, P.; She, K.; Zhang, Y.; Lin, X.; Li, B.; Chen, Z. The NAC-like transcription factor CsNAC7 positively regulates the caffeine biosynthesis-related gene yhNMT1 in Camellia sinensis. Hortic. Res. 2022, 9, uhab046. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Liu, Y.; Xie, Z.; Yu, B.; Sun, Y.; Huang, J. OsNAC016 regulates plant architecture and drought tolerance by interacting with the kinases GSK2 and SAPK8. Plant Physiol. 2022, 189, 1296–1313. [Google Scholar] [CrossRef]
- Liu, G.S.; Li, H.L.; Grierson, D.; Fu, D.Q. NAC Transcription Factor Family Regulation of Fruit Ripening and Quality: A Review. Cells 2022, 11, 525. [Google Scholar] [CrossRef]
- Xu, Y.; Zou, S.; Zeng, H.; Wang, W.; Wang, B.; Wang, H.; Tang, D. A NAC Transcription Factor TuNAC69 Contributes to ANK-NLR-WRKY NLR-Mediated Stripe Rust Resistance in the Diploid Wheat Triticum urartu. Int. J. Mol. Sci. 2022, 23, 564. [Google Scholar] [CrossRef]
- Wang, J.; Tian, S.; Yu, Y.; Ren, Y.; Guo, S.; Zhang, J.; Li, M.; Zhang, H.; Gong, G.; Wang, M.; et al. Natural variation in the NAC transcription factor NONRIPENING contributes to melon fruit ripening. J. Integr. Plant Biol. 2022, 64, 1448–1461. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Lv, B.; Li, J.; Luo, L.; Lu, S.; Zhang, X.; Ma, H.; Ming, F. The NAC family transcription factor OsNAP confers abiotic stress response through the ABA pathway. Plant Cell Physiol. 2014, 55, 604–619. [Google Scholar] [CrossRef] [Green Version]
- Rahman, H.; Ramanathan, V.; Nallathambi, J.; Duraialagaraja, S.; Muthurajan, R. Over-expression of a NAC 67 transcription factor from finger millet (Eleusine coracana L.) confers tolerance against salinity and drought stress in rice. BMC Biotechnol. 2016, 16 (Suppl. S1), 35. [Google Scholar] [CrossRef]
- Wang, B.; Zhong, Z.; Wang, X.; Han, X.; Yu, D.; Wang, C.; Song, W.; Zheng, X.; Chen, C.; Zhang, Y. Knockout of the OsNAC006 Transcription Factor Causes Drought and Heat Sensitivity in Rice. Int. J. Mol. Sci. 2020, 21, 2288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, J.; Lv, B.; Luo, L.; He, J.; Mao, C.; Xi, D.; Ming, F. The NAC-type transcription factor OsNAC2 regulates ABA-dependent genes and abiotic stress tolerance in rice. Sci. Rep. 2017, 7, 40641. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Hong, Y.; Zhang, H.; Li, D.; Song, F. Rice NAC transcription factor ONAC095 plays opposite roles in drought and cold stress tolerance. BMC Plant Biol. 2016, 16, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, X.; Wang, H.; Cai, J.; Bi, Y.; Li, D.; Song, F. Rice NAC transcription factor ONAC066 functions as a positive regulator of drought and oxidative stress response. BMC Plant Biol. 2019, 19, 278. [Google Scholar] [CrossRef]
- Ma, J.; Wang, L.Y.; Dai, J.X.; Wang, Y.; Lin, D. The NAC-type transcription factor CaNAC46 regulates the salt and drought tolerance of transgenic Arabidopsis thaliana. BMC Plant Biol. 2021, 21, 11. [Google Scholar] [CrossRef]
- Wang, Q.; Guo, C.; Li, Z.; Sun, J.; Deng, Z.; Wen, L.; Li, X.; Guo, Y. Potato NAC Transcription Factor StNAC053 Enhances Salt and Drought Tolerance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2021, 22, 2568. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, S.; Ma, X.; Wang, Y.; Kong, F.; Meng, Q. A stress-associated NAC transcription factor (SlNAC35) from tomato plays a positive role in biotic and abiotic stresses. Physiol. Plant. 2016, 158, 45–64. [Google Scholar] [CrossRef]
- Chong, L.; Xu, R.; Huang, P.; Guo, P.; Zhu, M.; Du, H.; Sun, X.; Ku, L.; Zhu, J.K.; Zhu, Y. The tomato OST1-VOZ1 module regulates drought-mediated flowering. Plant Cell 2022, 34, 2001–2018. [Google Scholar] [CrossRef]
- Thirumalaikumar, V.P.; Devkar, V.; Mehterov, N.; Ali, S.; Ozgur, R.; Turkan, I.; Mueller-Roeber, B.; Balazadeh, S. NAC transcription factor JUNGBRUNNEN1 enhances drought tolerance in tomato. Plant Biotechnol. J. 2018, 16, 354–366. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.; Zhang, H.; Huang, L.; Li, D.; Song, F. Overexpression of a Stress-Responsive NAC Transcription Factor Gene ONAC022 Improves Drought and Salt Tolerance in Rice. Front. Plant Sci. 2016, 7, 4. [Google Scholar] [CrossRef]
- Ju, Y.L.; Yue, X.F.; Min, Z.; Wang, X.H.; Fang, Y.L.; Zhang, J.X. VvNAC17, a novel stress-responsive grapevine (Vitis vinifera L.) NAC transcription factor, increases sensitivity to abscisic acid and enhances salinity, freezing, and drought tolerance in transgenic Arabidopsis. Plant Physiol. Biochem. 2020, 146, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Wang, Y.; Li, B.; Chang, J.; Chen, M.; Li, K.; Yang, G.; He, G. TaNAC29, a NAC transcription factor from wheat, enhances salt and drought tolerance in transgenic Arabidopsis. BMC Plant Biol. 2015, 15, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Rashotte, A.M.; Dane, F. Citrullus colocynthis NAC transcription factors CcNAC1 and CcNAC2 are involved in light and auxin signaling. Plant Cell Rep. 2014, 33, 1673–1686. [Google Scholar] [CrossRef] [PubMed]
- Diao, P.; Chen, C.; Zhang, Y.; Meng, Q.; Lv, W.; Ma, N. The role of NAC transcription factor in plant cold response. Plant Signal. Behav. 2020, 15, 1785668. [Google Scholar] [CrossRef]
- Wu, J.; Wang, L.; Wang, S. Comprehensive analysis and discovery of drought-related NAC transcription factors in common bean. BMC Plant Biol. 2016, 16, 193. [Google Scholar] [CrossRef] [Green Version]
- Kou, X.; Zhou, J.; Wu, C.E.; Yang, S.; Liu, Y.; Chai, L.; Xue, Z. The interplay between ABA/ethylene and NAC TFs in tomato fruit ripening: A review. Plant Mol. Biol. 2021, 106, 223–238. [Google Scholar] [CrossRef]
- Zhang, X.; Long, Y.; Chen, X.; Zhang, B.; Xin, Y.; Li, L.; Cao, S.; Liu, F.; Wang, Z.; Huang, H.; et al. A NAC transcription factor OsNAC3 positively regulates ABA response and salt tolerance in rice. BMC Plant Biol. 2021, 21, 546. [Google Scholar] [CrossRef]
- Zhang, X.; Long, Y.; Huang, J.; Xia, J. OsNAC45 is Involved in ABA Response and Salt Tolerance in Rice. Rice 2020, 13, 79. [Google Scholar] [CrossRef]
- Zhu, F.; Luo, T.; Liu, C.; Wang, Y.; Zheng, L.; Xiao, X.; Zhang, M.; Yang, H.; Yang, W.; Xu, R.; et al. A NAC transcription factor and its interaction protein hinder abscisic acid biosynthesis by synergistically repressing NCED5 in Citrus reticulata. J. Exp. Bot. 2020, 71, 3613–3625. [Google Scholar] [CrossRef]
- Lu, X.; Dun, H.; Lian, C.; Zhang, X.; Yin, W.; Xia, X. The role of peu-miR164 and its target PeNAC genes in response to abiotic stress in Populus euphratica. Plant Physiol. Biochem. 2017, 115, 418–438. [Google Scholar] [CrossRef]
- Wang, J.; Bao, J.; Zhou, B.; Li, M.; Li, X.; Jin, J. The osa-miR164 target OsCUC1 functions redundantly with OsCUC3 in controlling rice meristem/organ boundary specification. New Phytol. 2021, 229, 1566–1581. [Google Scholar] [CrossRef] [PubMed]
- Sosa-Valencia, G.; Palomar, M.; Covarrubias, A.A.; Reyes, J.L. The legume miR1514a modulates a NAC transcription factor transcript to trigger phasiRNA formation in response to drought. J. Exp. Bot. 2017, 68, 2013–2026. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Zhang, C.; Cao, D.; Damaris, R.N.; Yang, P. The Latest Studies on Lotus (Nelumbo nucifera)—An Emerging Horticultural Model Plant. Int. J. Mol. Sci. 2019, 20, 3680. [Google Scholar] [CrossRef] [Green Version]
- Cao, D.; Lin, Z.; Huang, L.; Damaris, R.N.; Yang, P. Genome-wide analysis of AP2/ERF superfamily in lotus (Nelumbo nucifera) and the association between NnADAP and rhizome morphology. BMC Genom. 2021, 22, 171. [Google Scholar] [CrossRef]
- Li, J.; Xiong, Y.; Li, Y.; Ye, S.; Yin, Q.; Gao, S.; Yang, D.; Yang, M.; Palva, E.T.; Deng, X. Comprehensive Analysis and Functional Studies of WRKY Transcription Factors in Nelumbo nucifera. Int. J. Mol. Sci. 2019, 20, 5006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, T.Y.; Liu, Y.Y.; Zhu, H.H.; Zhang, J.; Yang, J.X.; Fu, Q.; Wang, N.; Wang, Z. Genome-wide analyses of the bHLH gene family reveals structural and functional characteristics in the aquatic plant Nelumbo nucifera. PeerJ 2019, 7, e7153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Z.; Cao, D.; Damaris, R.N.; Yang, P. Genome-wide identification of MADS-box gene family in sacred lotus (Nelumbo nucifera) identifies a SEPALLATA homolog gene involved in floral development. BMC Plant Biol. 2020, 20, 497. [Google Scholar] [CrossRef] [PubMed]
- Ming, R.; VanBuren, R.; Liu, Y.; Yang, M.; Han, Y.; Li, L.T.; Zhang, Q.; Kim, M.J.; Schatz, M.C.; Campbell, M.; et al. Genome of the long-living sacred lotus (Nelumbo nucifera Gaertn.). Genome Biol. 2013, 14, R41. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Fan, G.; Liu, Y.; Sun, F.; Shi, C.; Liu, X.; Peng, J.; Chen, W.; Huang, X.; Cheng, S.; et al. The sacred lotus genome provides insights into the evolution of flowering plants. Plant J. 2013, 76, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Wang, T.; Cheng, T.; Zhao, L.; Zheng, X.; Zhu, F.; Dong, C.; Xu, J.; Xie, K.; Hu, Z.; et al. Genomic variation reveals demographic history and biological adaptation of the ancient relictual, lotus (Nelumbo Adans). Hortic. Res. 2022, 9, uhac029. [Google Scholar] [CrossRef]
- Ooka, H.; Satoh, K.; Doi, K.; Nagata, T.; Otomo, Y.; Murakami, K.; Matsubara, K.; Osato, N.; Kawai, J.; Carninci, P.; et al. Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana. DNA Res. 2003, 10, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, Y.; Shi, S.; Zhou, Y.; Zhou, Y.; Yang, J.; Tang, X. Genome-wide identification and characterization of GRAS transcription factors in sacred lotus (Nelumbo nucifera). PeerJ 2016, 4, e2388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas-Hernandez, B.Y.; Nunez-Munoz, L.; Calderon-Perez, B.; Xoconostle-Cazares, B.; Ruiz-Medrano, R. The NAC Transcription Factor ANAC087 Induces Aerial Rosette Development and Leaf Senescence in Arabidopsis. Front. Plant Sci. 2022, 13, 818107. [Google Scholar] [CrossRef]
- Tao, Y.; Wan, J.X.; Liu, Y.S.; Yang, X.Z.; Shen, R.F.; Zhu, X.F. The NAC transcription factor ANAC017 regulates aluminum tolerance by regulating the cell wall-modifying genes. Plant Physiol. 2022, 189, 2517–2534. [Google Scholar] [CrossRef]
- Song, S.; Willems, L.A.J.; Jiao, A.; Zhao, T.; Eric Schranz, M.; Bentsink, L. The membrane associated NAC transcription factors ANAC060 and ANAC040 are functionally redundant in the inhibition of seed dormancy in Arabidopsis thaliana. J. Exp. Bot. 2022, 73, 5514–5528. [Google Scholar] [CrossRef] [PubMed]
- Mao, H.; Li, S.; Chen, B.; Jian, C.; Mei, F.; Zhang, Y.; Li, F.; Chen, N.; Li, T.; Du, L.; et al. Variation in cis-regulation of a NAC transcription factor contributes to drought tolerance in wheat. Mol. Plant 2022, 15, 276–292. [Google Scholar] [CrossRef]
- Luo, P.; Chen, Y.; Rong, K.; Lu, Y.; Wang, N.; Xu, Z.; Pang, B.; Zhou, D.; Weng, J.; Li, M.; et al. ZmSNAC13, a maize NAC transcription factor conferring enhanced resistance to multiple abiotic stresses in transgenic Arabidopsis. Plant Physiol. Biochem. 2022, 170, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Lin, Y.; Xu, M.; Bian, H.; Zhang, C.; Wang, J.; Wang, H.; Xu, Y.; Niu, Q.; Zuo, J.; et al. The role and interaction between transcription factor NAC-NOR and DNA demethylase SlDML2 in the biosynthesis of tomato fruit flavor volatiles. New Phytol. 2022, 235, 1913–1926. [Google Scholar] [CrossRef]
- Du, Q.; Lv, W.; Guo, Y.; Yang, J.; Wang, S.; Li, W.X. MIR164b represses iron uptake by regulating the NAC domain transcription factor5-Nuclear Factor Y, Subunit A8 module in Arabidopsis. Plant Physiol. 2022, 189, 1095–1109. [Google Scholar] [CrossRef]
- Song, H.; Liu, Y.; Dong, G.; Zhang, M.; Wang, Y.; Xin, J.; Su, Y.; Sun, H.; Yang, M. Genome-Wide Characterization and Comprehensive Analysis of NAC Transcription Factor Family in Nelumbo nucifera. Front. Genet. 2022, 13, 901838. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Zhang, Q.; Shi, Y.; Sun, Z.; Zhang, Q.; Hou, S.; Wu, R.; Jiang, L.; Zhao, X.; Guo, Y. Genome-Wide Analysis of the NAC Domain Transcription Factor Gene Family in Theobroma cacao. Genes 2019, 11, 35. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Ma, Z.; Sun, W.; Huang, L.; Wu, Q.; Tang, Z.; Bu, T.; Li, C.; Chen, H. Genome-wide analysis of the NAC transcription factor family in Tartary buckwheat (Fagopyrum tataricum). BMC Genom. 2019, 20, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.; Liu, J.; Du, B.; Zhang, M.; Wang, A.; Zhang, L. NAC Transcription Factor PwNAC11 Activates ERD1 by Interaction with ABF3 and DREB2A to Enhance Drought Tolerance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2021, 22, 6952. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.Y.; Liu, X.F.; Fu, B.L.; Zhang, Q.Y.; Tong, Y.; Wang, J.; Wang, W.Q.; Grierson, D.; Yin, X.R. Methyl Jasmonate Enhances Ethylene Synthesis in Kiwifruit by Inducing NAC Genes That Activate ACS1. J. Agric. Food Chem. 2020, 68, 3267–3276. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, M.; Wang, S. Processing characteristics and flavour of full lotus root powder beverage. J. Sci. Food Agric. 2010, 90, 2482–2489. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Li, S.; Hussain, J.; Xu, X.; Yin, J.; Zhang, Y.; Chen, X.; Li, L. Isolation and functional characterization of a salt responsive transcriptional factor, LrbZIP from lotus root (Nelumbo nucifera Gaertn). Mol. Biol. Rep. 2013, 40, 4033–4045. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Rombauts, S.; Dehais, P.; Van Montagu, M.; Rouze, P. PlantCARE, a plant cis-acting regulatory element database. Nucleic Acids Res. 1999, 27, 295–296. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.P.; Xu, Z.S.; Zheng, W.J.; Zhao, W.; Wang, Y.X.; Yu, T.F.; Chen, M.; Zhou, Y.B.; Min, D.H.; Ma, Y.Z.; et al. Genome-Wide Analysis of the RAV Family in Soybean and Functional Identification of GmRAV-03 Involvement in Salt and Drought Stresses and Exogenous ABA Treatment. Front. Plant Sci. 2017, 8, 905. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Genomic (bp) | Transcript (bp) | CDS (bp) | Amino Acid Residues (aa) | pI | MW (KDa) |
|---|---|---|---|---|---|---|---|
| NnAC001 | Nn1g00014 | 808 | 571 | 571 | 190 | 4.48 | 21.73 |
| NnNAC002 | Nn1g00424 | 1478 | 1219 | 1219 | 406 | 6.78 | 38.7 |
| NnNAC003 | Nn1g00471 | 3868 | 1768 | 1768 | 589 | 9.59 | 68.3 |
| NnNAC004 | Nn1g00574 | 2293 | 1147 | 1147 | 382 | 5.16 | 43.21 |
| NnNAC005 | Nn1g01090 | 3088 | 1138 | 1138 | 379 | 6.35 | 41.38 |
| NnNAC006 | Nn1g02798 | 4088 | 847 | 847 | 282 | 9.38 | 19 |
| NnNAC007 | Nn1g03472 | 825 | 628 | 628 | 209 | 9.44 | 24.45 |
| NnNAC008 | Nn1g03986 | 2380 | 1171 | 1171 | 390 | 9.29 | 43.13 |
| NnNAC009 | Nn1g03987 | 549 | 286 | 286 | 95 | 7.01 | 41.53 |
| NnNAC010 | Nn1g04183 | 2056 | 1279 | 1279 | 426 | 6.25 | 48.01 |
| NnNAC011 | Nn1g04185 | 1898 | 1150 | 1150 | 383 | 6.57 | 43.76 |
| NnNAC012 | Nn1g04206 | 1652 | 1069 | 1069 | 356 | 8.36 | 39.32 |
| NnNAC013 | Nn1g04246 | 1929 | 973 | 973 | 324 | 7.79 | 41.69 |
| NnNAC014 | Nn1g04798 | 38,082 | 772 | 772 | 257 | 7.66 | 38.71 |
| NnNAC015 | Nn1g05100 | 4004 | 430 | 430 | 143 | 4.76 | 30.56 |
| NnNAC016 | Nn1g05179 | 2747 | 1675 | 1675 | 559 | 4.89 | 63.62 |
| NnNAC017 | Nn1g06246 | 8935 | 1390 | 1390 | 463 | 4.93 | 51.29 |
| NnNAC018 | Nn1g06289 | 811 | 571 | 571 | 190 | 4.48 | 21.82 |
| NnNAC019 | Nn1g06403 | 2271 | 964 | 964 | 321 | 6.25 | 26.5 |
| NnNAC020 | Nn1g06441 | 2864 | 1033 | 1033 | 344 | 8.01 | 37.92 |
| NnNAC021 | Nn1g06666 | 2070 | 841 | 841 | 280 | 9.16 | 44.2 |
| NnNAC022 | Nn1g06671 | 1365 | 1063 | 1063 | 354 | 8.89 | 40.32 |
| NnNAC023 | Nn1g08315 | 1708 | 1279 | 1279 | 426 | 6.13 | 48.21 |
| NnNAC024 | Nn1g08339 | 1770 | 1282 | 1282 | 427 | 6.13 | 48.28 |
| NnNAC025 | Nn1g08874 | 907 | 907 | 907 | 302 | 8.39 | 34.63 |
| NnNAC026 | Nn2g12755 | 7485 | 949 | 949 | 316 | 8.77 | 35.3 |
| NnNAC027 | Nn2g13806 | 1016 | 649 | 649 | 216 | 4.93 | 26.4 |
| NnNAC028 | Nn2g13861 | 1328 | 820 | 820 | 273 | 4.64 | 62.23 |
| NnNAC029 | Nn2g14216 | 1499 | 622 | 622 | 207 | 9.34 | 23.41 |
| NnNAC030 | Nn2g14870 | 1290 | 532 | 532 | 177 | 9.15 | 21.13 |
| NnNAC031 | Nn2g14874 | 1991 | 1012 | 1012 | 337 | 6.07 | 38.87 |
| NnNAC032 | Nn2g15917 | 1522 | 1246 | 1246 | 415 | 9.46 | 23.88 |
| NnNAC033 | Nn2g15921 | 3819 | 1153 | 1153 | 384 | 8.48 | 43.41 |
| NnNAC034 | Nn3g16348 | 922 | 229 | 229 | 76 | 9.86 | 18.71 |
| NnNAC035 | Nn3g16672 | 348 | 349 | 349 | 116 | 8.91 | 13.64 |
| NnNAC036 | Nn3g16695 | 9610 | 1183 | 1183 | 394 | 5.6 | 43.89 |
| NnNAC037 | Nn3g17225 | 2541 | 1255 | 1255 | 418 | 6.66 | 46.29 |
| NnNAC038 | Nn3g17296 | 1617 | 1069 | 1069 | 256 | 8.99 | 33.49 |
| NnNAC039 | Nn3g17297 | 1123 | 847 | 847 | 282 | 6.26 | 32.37 |
| NnNAC040 | Nn3g17722 | 14,012 | 1480 | 1480 | 493 | 5.09 | 55.55 |
| NnNAC041 | Nn3g17854 | 1580 | 1009 | 1009 | 336 | 7.54 | 37.46q |
| NnNAC042 | Nn3g18857 | 1031 | 1222 | 1222 | 407 | 6.89 | 46.14 |
| NnNAC043 | Nn3g19685 | 4659 | 850 | 850 | 283 | 5.58 | 31.98 |
| NnNAC044 | Nn3g21809 | 1945 | 349 | 349 | 116 | 9.49 | 13.55 |
| NnNAC045 | Nn4g22208 | 1379 | 775 | 775 | 258 | 9.38 | 29.22 |
| NnNAC046 | Nn4g22597 | 7186 | 1354 | 1354 | 451 | 5.5 | 50.44 |
| NnNAC047 | Nn4g23151 | 1916 | 1234 | 1234 | 411 | 8.25 | 45.91 |
| NnNAC048 | Nn4g23226 | 1304 | 1072 | 1072 | 357 | 7.05 | 40.27 |
| NnNAC049 | Nn4g23228 | 816 | 478 | 478 | 159 | 9.41 | 18.61 |
| NnNAC050 | Nn4g23242 | 1085 | 847 | 847 | 282 | 6.67 | 32.54 |
| NnNAC051 | Nn4g23388 | 701 | 514 | 514 | 171 | 6.66 | 20.03 |
| NnNAC052 | Nn4g23853 | 1406 | 904 | 904 | 301 | 7 | 34.07 |
| NnNAC053 | Nn4g24534 | 372 | 373 | 373 | 124 | 9.98 | 14.2 |
| NnNAC054 | Nn4g24728 | 4125 | 2053 | 2053 | 684 | 5.31 | 77.94 |
| NnNAC055 | Nn4g24729 | 2437 | 715 | 715 | 238 | 5.06 | 27.26 |
| NnNAC056 | Nn4g24873 | 2218 | 1054 | 1054 | □ | 8.84 | 40.27 |
| NnNAC057 | Nn4g25922 | 1642 | 460 | 460 | 153 | 9.92 | 18.13 |
| NnNAC058 | Nn4g25937 | 1644 | 460 | 460 | 153 | 9.7 | 18.1 |
| NnNAC059 | Nn4g26681 | 2233 | 964 | 964 | 321 | 7.69 | 37.02 |
| NnNAC060 | Nn5g27114 | 2326 | 481 | 481 | 160 | 9.04 | 18.97 |
| NnNAC061 | Nn5g27174 | 4421 | 1081 | 1081 | 360 | 8.13 | 40.74 |
| NnNAC062 | Nn5g27709 | 1144 | 943 | 943 | 314 | 8.97 | 19.65 |
| NnNAC063 | Nn5g28365 | 394 | 490 | 490 | 163 | 5.84 | 34.06 |
| NnNAC064 | Nn5g28509 | 1373 | 592 | 592 | 197 | 7.7 | 21.99 |
| NnNAC065 | Nn5g28579 | 9445 | 1387 | 1387 | 462 | 5.03 | 46.52 |
| NnNAC066 | Nn5g28895 | 3086 | 1705 | 1705 | 568 | 9.53 | 61.43 |
| NnNAC067 | Nn5g29253 | 4770 | 1027 | 1027 | 342 | 5.37 | 38.58 |
| NnNAC068 | Nn5g29499 | 3786 | 397 | 397 | 132 | 5.81 | 45.6 |
| NnNAC069 | Nn5g30346 | 8202 | 658 | 658 | 219 | 9.53 | 22.18 |
| NnNAC070 | Nn5g30910 | 2903 | 895 | 895 | 298 | 8.55 | 29.31 |
| NnNAC071 | Nn5g31036 | 2104 | 1111 | 1111 | 370 | 5.74 | 41.69 |
| NnNAC072 | Nn6g31660 | 2301 | 1156 | 1156 | 385 | 6.36 | 42.44 |
| NnNAC073 | Nn6g31768 | 1749 | 772 | 772 | 257 | 8.59 | 35.64 |
| NnNAC074 | Nn6g32265 | 2062 | 1000 | 1000 | 333 | 5.77 | 38.39 |
| NnNAC075 | Nn6g33629 | 3148 | 271 | 271 | 90 | 9.44 | 29.17 |
| NnNAC076 | Nn6g34050 | 2482 | 1099 | 1099 | 366 | 6.81 | 42.52 |
| NnNAC077 | Nn6g34819 | 1205 | 760 | 760 | 253 | 6 | 28.76 |
| NnNAC078 | Nn6g34820 | 1464 | 793 | 793 | 264 | 6.8 | 30.26 |
| NnNAC079 | Nn6g35218 | 16,416 | 1771 | 1771 | 590 | 4.66 | 54.24 |
| NnNAC080 | Nn6g35359 | 3153 | 874 | 874 | 291 | 6.6 | 47.95 |
| NnNAC081 | Nn7g35779 | 1087 | 913 | 913 | 304 | 5.38 | 34.73 |
| NnNAC082 | Nn7g37118 | 1575 | 1015 | 1015 | 338 | 5.46 | 38.78 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, S.; Jiang, T.; Zhang, Y.; Zhang, K.; Feng, K.; Wu, P.; Li, L. Identification of the NAC Transcription Factors and Their Function in ABA and Salinity Response in Nelumbo nucifera. Int. J. Mol. Sci. 2022, 23, 12394. https://doi.org/10.3390/ijms232012394
Zhao S, Jiang T, Zhang Y, Zhang K, Feng K, Wu P, Li L. Identification of the NAC Transcription Factors and Their Function in ABA and Salinity Response in Nelumbo nucifera. International Journal of Molecular Sciences. 2022; 23(20):12394. https://doi.org/10.3390/ijms232012394
Chicago/Turabian StyleZhao, Shuping, Tao Jiang, Yao Zhang, Kailing Zhang, Kai Feng, Peng Wu, and Liangjun Li. 2022. "Identification of the NAC Transcription Factors and Their Function in ABA and Salinity Response in Nelumbo nucifera" International Journal of Molecular Sciences 23, no. 20: 12394. https://doi.org/10.3390/ijms232012394