

cd1 Mutation in Drosophila Affects Phenoxazinone Synthase Catalytic Site and Impairs Long-Term Memory
 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
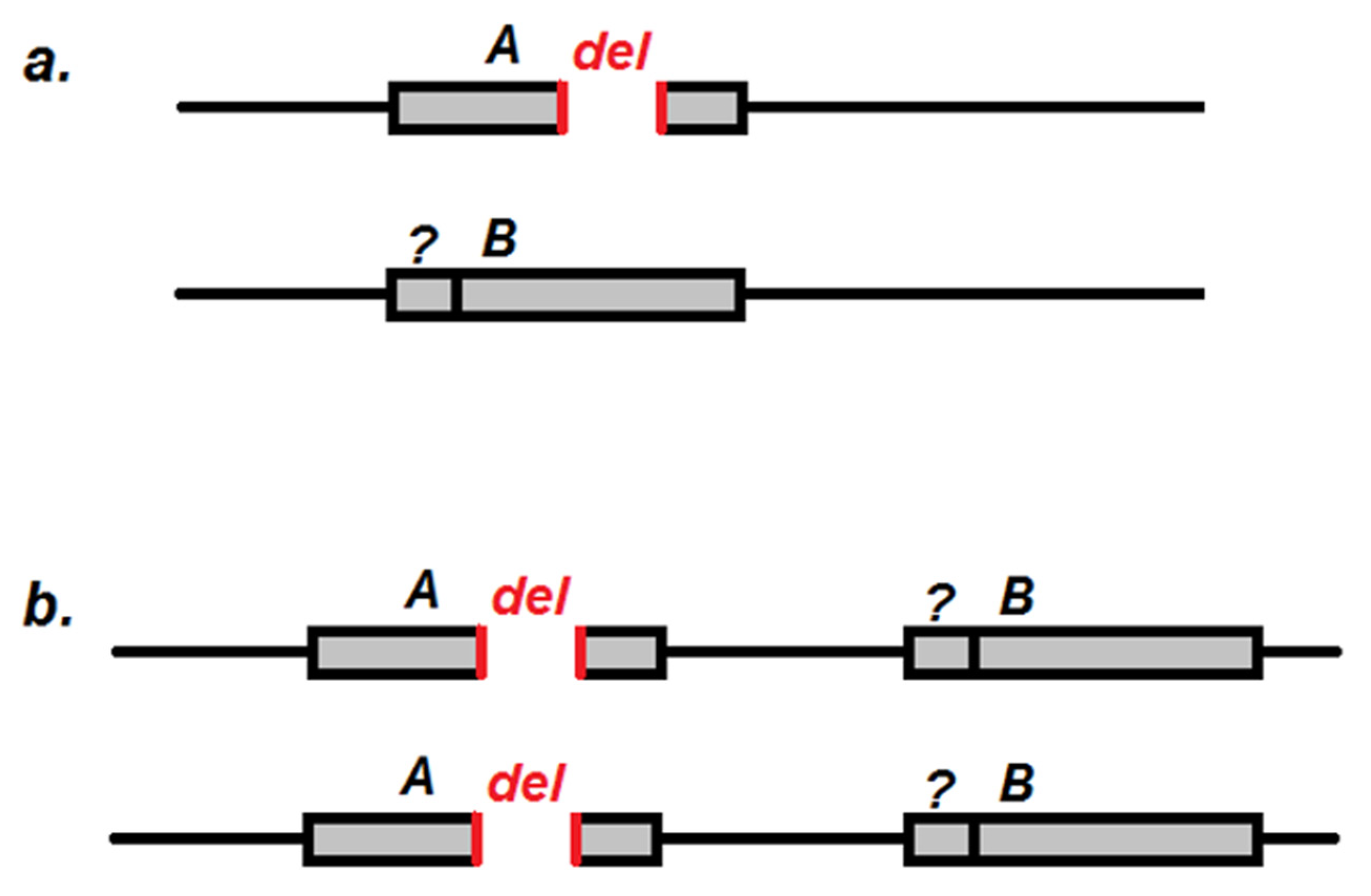
2.1. PCR Mapping of the cd Gene
2.2. The cd Gene Sequencing in CS and cd1
2.3. The cd Gene Expression in Late Pupae
2.4. Learning and Long-Term Memory Formation
2.5. Dopaminergic Neurons in Adult Drosophila Brain
3. Discussion
- The olfactory sensory neurons Or67d for anti-aphrodisiac cis-vaccenyl acetate (cVA), which is crucial for courtship suppression [34].
- The visual projection neurons (VPN) from the optic lobe (OL).
- MB calyx (Cal) and intrinsic neurons. MB are responsible for the classical olfactory conditioning [46] and are not required for leaning in CCSP [47,48]. However, Kenyon cells (KCs) of MB receive inputs not only from the olfactory AL. Subesophageal ganglion (SEG) responsible for taste sensitivity activates the dorsal accessory calyx (dAC) [49]. γd neurons of the ventral accessory calyx (vAC) participate in CCSP STM formation, receiving visual inputs from OL [48,50]. These neurons project to the MB γ5 area, crucial for courtship suppression [51].
- The intrinsic neurons of MB projections to the MB output neurons (MBONs). Their synaptic contacts are tightly regulated by DANs, activated by unconditioned stimuli, such as electric shock or sugar. They do not encode specific modality of a stimulus, rather its positive or negative valence. After learning, the mutual MBON effect is shifted towards attraction or avoidance, biasing the fly’s behavior [52].
- MBON project to the higher brain integrative centers, such as SMP, lateral protocerebrum (LP) and lateral horn (LH), also containing about 90% of the DANs dendritic arbors [50,52]. LP is the major site for the integration of discrete sensory stimulus, aggregated by P1 cluster, which triggers male courtship [44,45].
4. Materials and Methods
4.1. Drosophila Strains
- Canton-S (CS), the wild type strain; dark red eyes.
- cinnabar (cn1)—this carries a mutation in the kynurenine monooxygenase gene; lack of 3-HOK, KYNA excess, bright red eyes.
- cardinal (cd1; Bloomington Drosophila Stock Center, #3052)—this carries a mutation in the cd gene; increase in 3-HOK level, bright red eyes after eclosion; in 3-day-old flies the eye color becomes dark red.
- 4.
- Strains from Bloomington Drosophila Stock Center:
4.2. DNA Extraction
4.3. Polymerase Chain Reaction
4.4. DNA Purification and Sequencing
4.5. Bioinformatics Analysis
4.6. RNA Extraction, Reverse Transcription and Semi-Quantitative Real-Time Polymerase Chain Reaction (sq-RT-PCR)
4.7. Long-Term Memory in Conditioned Courtship Suppression Paradigm
4.8. Dopaminergic Neuron Visualization
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Lapin, I.P. Kynurenines as probable participants of depression. Pharmakopsychiatr. Neuro-Psychopharmacol. 1973, 6, 273–279. [Google Scholar] [CrossRef]
- Oxenkrug, G. Serotonin-kynurenine hypothesis of depression: Historical overview and recent developments. Curr Drug Targets 2013, 14, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Bryleva, E.Y.; Brundin, L. Kynurenine pathway metabolites and suicidality. Neuropharmacology 2017, 112, 324–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarcz, R.; Bruno, J.P.; Muchowski, P.J.; Wu, H.Q. Kynurenines in the mammalian brain: When physiology meets pathology. Nat. Rev. Neurosci 2012, 13, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.M.; Charych, E.; Lee, A.W.; Möller, T. Kynurenines in CNS disease: Regulation by inflammatory cytokines. Front Neurosci. 2014, 8, 12. [Google Scholar] [CrossRef] [Green Version]
- Lapin, I. Stress, Anxiety, Depression, Alcoholism, Epilepsy (The Neuroactivities of Kynurenines and New Approaches to Treatment); DEAN: Saint Petersburg, Russia, 2004. [Google Scholar]
- Badawy, A.A. Kynurenine pathway of tryptophan metabolism: Regulatory and functional aspects. Int. J. Tryptophan. Res. 2017, 10, 1938. [Google Scholar] [CrossRef] [Green Version]
- Linzen, B. The tryptophan to ommochrome pathway in insects. Adv. Insect Physiol. 1974, 10, 117–246. [Google Scholar] [CrossRef]
- Zhuravlev, A.V.; Nikitina, E.A.; Savvateeva-Popova, E.V. Role of kynurenines in regulation of behavior and memory processes in Drosophila. Integr. Physiol. 2020, 1, 40–50. [Google Scholar] [CrossRef]
- Summers, K.M.; Howells, A.J.; Pyliotis, N.A. Biology of eye pigmentation in insects. Adv. Insect Physiol 1982, 16, 119–166. [Google Scholar] [CrossRef]
- Ryall, R.L.; Howells, A.J. Ommochrome biosynthetic pathway of Drosophila melanogaster: Variations in levels of enzyme activities and intermediates during adult development. Insect Biochem. 1974, 4, 47–61. [Google Scholar] [CrossRef]
- Ferré, J. Accumulation of kynurenic acid in the cinnabar mutant of D. melanogaster as revealed by thin-layer chromatography. Insect Biochem. 1983, 13, 289–294. [Google Scholar] [CrossRef]
- Phillips, J.P.; Forrest, H.S.; Kulkarni, A.D. Terminal synthesis of xanthommatin in Drosophila melanogaster. 3. Mutational pleiotropy and pigment granule association of phenoxazinone synthetase. Genetics 1973, 73, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Howells, A.J.; Summers, K.M.; Ryall, R.L. Developmental patterns of 3-hydroxykynurenine accumulation in white and various other color mutants of Drosophila melanogaster. Biochem Genet. 1977, 15, 1049–1059. [Google Scholar] [CrossRef]
- Li, J.; Beerntsen, B.T.; James, A.A. Oxidation of 3-hydroxykynurenine to produce xanthommatin for eye pigmentation: A major branch pathway of tryptophan catabolism during pupal development in the yellow fever mosquito, Aedes Aegypti. Insect Biochem. Mol. Biol. 1999, 29, 329–338. [Google Scholar] [CrossRef]
- Zhuravlev, A.V.; Vetrovoy, O.; Savvateeva-Popova, E.V. Enzymatic and non-enzymatic pathways of kynurenines’ dimerization: The molecular factors for oxidative stress development kynyrenines’ dimerization and its impact in oxidative stress. PLoS Comput. Biol. 2018, 14, e1006672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christen, S.; Peterhans, E.; Stocker, R. Antioxidant activities of some tryptophan metabolites: Possible implication for inflammatory diseases. Proc. Natl. Acad. Sci. USA 1990, 87, 2506–2510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuravlev, A.V.; Zakharov, G.A.; Shchegolev, B.F.; Savvateeva-Popova, E.V. Antioxidant properties of kynurenines: Density functional theory calculations. PLoS Comput. Biol. 2016, 12, e1005213. [Google Scholar] [CrossRef] [Green Version]
- Okuda, S.; Nishiyama, N.; Saito, H.; Katsuki, H. Hydrogen peroxide-mediated neuronal cell death induced by an endogenous neurotoxin, 3-hydroxykynurenine. Proc. Natl. Acad. Sci. USA 1996, 93, 12553–12558. [Google Scholar] [CrossRef] [Green Version]
- Okuda, S.; Nishiyama, N.; Saito, H.; Katsuki, H. 3-Hydroxykynurenine, an endogenous oxidative stress generator, causes neuronal cell death with apoptotic features and region selectivity. J. Neurochem 1998, 70, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Colín-González, A.L.; Maya-López, M.; Pedraza-Chaverrí, J.; Ali, S.F.; Chavarría, A.; Santamaría, A. The Janus faces of 3-hydroxykynurenine: Dual redox modulatory activity and lack of neurotoxicity in the rat striatum. Brain Res. 2014, 1589, 1–14. [Google Scholar] [CrossRef]
- Kessler, M.; Terramani, T.; Lynch, G.; Baudry, M. A glycine site associated with N-methyl-D-aspartic acid receptors: Characterization and identification of a new class of antagonists. J. Neurochem. 1989, 52, 1319–1328. [Google Scholar] [CrossRef]
- Zhuravlev, A.V.; Zakharov, G.A.; Shchegolev, B.F.; Savvateeva-Popova, E.V. Stacking interaction and its role in kynurenic acid binding to glutamate ionotropic receptors. J. Mol. Model. 2012, 18, 1755–1766. [Google Scholar] [CrossRef] [PubMed]
- Hilmas, C.; Pereira, E.F.; Alkondon, M.; Rassoulpour, A.; Schwarcz, R.; Albuquerque, E.X. The brain metabolite kynurenic acid inhibits alpha7 nicotinic receptor activity and increases non-alpha7 nicotinic receptor expression: Physiopathological implications. J. Neurosci. 2001, 21, 7463–7473. [Google Scholar] [CrossRef] [PubMed]
- Campesan, S.; Green, E.W.; Breda, C.; Sathyasaikumar, K.; Muchowski, P.J.; Schwarcz, R.; Kyriacou, C.P.; Giorgini, F. The kynurenine pathway modulates neurodegeneration in a Drosophila model of Huntington’s disease. Curr. Biol. 2011, 21, 961–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, E.W.; Campesan, S.; Breda, C.; Sathyasaikumar, K.V.; Muchowski, P.J.; Schwarcz, R.; Kyriacou, C.P.; Giorgini, F. Drosophila eye color mutants as therapeutic tools for Huntington disease. Fly 2012, 6, 117–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savvateeva, E.; Popov, A.; Kamyshev, N.; Bragina, J.; Heisenberg, M.; Senitz, D.; Kornhuber, J.; Riederer, P. Age-dependent memory loss, synaptic pathology and altered brain plasticity in the Drosophila mutant cardinal accumulating 3-hydroxykynurenine. J. Neural Transm. 2000, 107, 581–601. [Google Scholar] [CrossRef] [PubMed]
- Savvateeva-Popova, E.V.; Popov, A.V.; Heinemann, T.; Riederer, P. Drosophila mutants of the kynurenine pathway as a model for ageing studies. Adv. Exp. Med. Biol. 2003, 527, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Zhuravlev, A.V.; Vetrovoy, O.V.; Ivanova, P.N.; Savvateeva-Popova, E.V. 3-hydroxykynurenine in regulation of Drosophila behavior: The novel mechanisms for cardinal phenotype manifestations. Front. Physiol. 2020, 11, 971. [Google Scholar] [CrossRef]
- Morgan, T.H.; Bridges, C.B.; Sturtevant, A.H. The Genetics of Drosophila; Bibliographia Genetica: Gravenhage, The Netherlands, 1925; Volume 2, pp. 1–262. [Google Scholar]
- Yamamoto, M.; Howells, A.J.; Ryall, R.L. The ommochrome biosynthetic pathway in Drosophila melanogaster: The head particulate phenoxazinone synthase and the developmental onset of xanthommatin synthesis. Biochem. Genet. 1976, 14, 1077–1090. [Google Scholar] [CrossRef]
- Wiley, K.; Forrest, H.S. Terminal synthesis of xanthommatin in Drosophila melanogaster. IV. Enzymatic and nonenzymatic catalysis. Biochem. Genet. 1981, 19, 1211–1221. [Google Scholar] [CrossRef]
- Harris, D.A.; Kim, K.; Nakahara, K.; Vásquez-Doorman, C.; Carthew, R.W. Cargo sorting to lysosome-related organelles regulates siRNA-mediated gene silencing. J. Cell Biol. 2011, 194, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Keleman, K.; Vrontou, E.; Krüttner, S.; Yu, J.Y.; Kurtovic-Kozaric, A.; Dickson, B.J. Dopamine neurons modulate pheromone responses in Drosophila courtship learning. Nature 2012, 489, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Davis, R.L. Eight different types of dopaminergic neurons innervate the Drosophila mushroom body neuropil: Anatomical and physiological heterogeneity. Front. Neural Circuits 2009, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, P.C.; Waldeck, K.; Ganetzky, B.; Babcock, D.T. Neurodegeneration and locomotor dysfunction in Drosophila scarlet mutants. J. Cell Sci. 2018, 131, jcs216697. [Google Scholar] [CrossRef] [Green Version]
- Navarro, J.A.; Heßner, S.; Yenisetti, S.C.; Bayersdorfer, F.; Zhang, L.; Voigt, A.; Schneuwly, S.; Botella, J.A. Analysis of dopaminergic neuronal dysfunction in genetic and toxin-induced models of Parkinson’s disease in Drosophila. J. Neurochem 2014, 131, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.; Wang, B.; Zhang, J.; Yan, T.; Yang, W.; Jiao, F.; Liu, J.; Wang, S. Localization of serotonin/tryptophan-hydroxylase-immunoreactive cells in the brain and suboesophageal ganglion of Drosophila melanogaster. Cell Tissue Res. 2010, 340, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Kasture, A.S.; Hummel, T.; Sucic, S.; Freissmuth, M. Big Lessons from Tiny Flies: Drosophila melanogaster as a Model to Explore Dysfunction of Dopaminergic and Serotonergic Neurotransmitter Systems. Int. J. Mol. Sci. 2018, 19, 1788. [Google Scholar] [CrossRef] [Green Version]
- Benzer, S. From the Gene to Behavior. JAMA 1971, 218, 1015–1022. [Google Scholar] [CrossRef] [PubMed]
- Zhuravlev, A.V.; Shchegolev, B.F.; Zakharov, G.A.; Ivanova, P.N.; Nikitina, E.A.; Savvateeva-Popova, E.V. 3-Hydroxykynurenine as a Potential Ligand for Hsp70 Proteins and Its Effects on Drosophila Memory After Heat Shock. Mol. Neurobiol. 2022, 59, 1862–1871. [Google Scholar] [CrossRef]
- Nikitina, E.A.; Zhuravlev, A.V.; Savvateeva-Popova, E.V. Effect of impaired kynurenine synthesis on memory in Drosophila. Integr. Physiology. 2021, 2, 49–60. (In Russian) [Google Scholar] [CrossRef]
- Zhuravlev, A.V.; Nikitina, E.A.; Savvateeva-Popova, E.V. Drosophila learning and memory: Physiological and genetic bases. Uspekhi. Fiziol. Nauk. 2015, 46, 76–92. (In Russian) [Google Scholar]
- Yu, J.Y.; Kanai, M.I.; Demir, E.; Jefferis, G.S.; Dickson, B.J. Cellular organization of the neural circuit that drives Drosophila courtship behavior. Curr. Biol. 2010, 20, 1602–1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlou, H.J.; Goodwin, S.F. Courtship behavior in Drosophila melanogaster: Towards a ‘courtship connectome’. Curr. Opin. Neurobiol. 2013, 23, 76–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heisenberg, M.; Borst, A.; Wagner, S.; Byers, D. Drosophila mushroom body mutants are deficient in olfactory learning. J. Neurogenet. 1985, 2, 1–30. [Google Scholar] [CrossRef]
- Wolf, R.; Wittig, T.; Liu, L.; Wustmann, G.; Eyding, D.; Heisenberg, M. Drosophila mushroom bodies are dispensable for visual, tactile, and motor learning. Learn. Mem. 1998, 5, 166–178. [Google Scholar] [CrossRef]
- Montague, S.A.; Baker, B.S. Memory elicited by courtship conditioning requires mushroom body neuronal subsets similar to those utilized in appetitive memory. PLoS ONE 2016, 11, e0164516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagi, R.; Mabuchi, Y.; Mizunami, M.; Tanaka, N.K. Convergence of multimodal sensory pathways to the mushroom body calyx in Drosophila melanogaster. Sci. Rep. 2016, 6, 29481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, K.; Aso, Y.; Hige, T.; Knapek, S.; Ichinose, T.; Friedrich, A.B.; Turner, G.C.; Rubin, G.M.; Tanimoto, H. Direct neural pathways convey distinct visual information to Drosophila mushroom bodies. eLife 2016, 5, e14009. [Google Scholar] [CrossRef] [PubMed]
- Aso, Y.; Hattori, D.; Yu, Y.; Johnston, R.M.; Iyer, N.A.; Ngo, T.T.; Dionne, H.; Abbott, L.F.; Axel, R.; Tanimoto, H.; et al. The neuronal architecture of the mushroom body provides a logic for associative learning. eLife 2014, 3, e04577. [Google Scholar] [CrossRef] [PubMed]
- Aso, Y.; Sitaraman, D.; Ichinose, T.; Kaun, K.R.; Vogt, K.; Belliart-Guérin, G.; Plaçais, P.Y.; Robie, A.A.; Yamagata, N.; Schnaitmann, C.; et al. Mushroom body output neurons encode valence and guide memory-based action selection in Drosophila. eLife 2014, 3, e04580. [Google Scholar] [CrossRef]
- Kamyshev, N.G.; Iliadi, K.G.; Bragina, J.V. Drosophila conditioned courtship: Two ways of testing memory. Learn. Mem. 1999, 6, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Redt-Clouet, C.; Trannoy, S.; Boulanger, A.; Tokmatcheva, E.; Savvateeva-Popova, E.; Parmentier, M.L.; Preat, T.; Dura, J.M. Mushroom body neuronal remodelling is necessary for short-term but not for long-term courtship memory in Drosophila. Eur. J. Neurosci 2012, 35, 1684–1691. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Lenek, D.; Dag, U.; Dickson, B.J.; Keleman, K. Persistent activity in a recurrent circuit underlies courtship memory in Drosophila. Elife 2018, 7, e31425. [Google Scholar] [CrossRef]
- Keleman, K.; Krüttner, S.; Alenius, M.; Dickson, B.J. Function of the Drosophila CPEB protein Orb2 in long-term courtship memory. Nat. Neurosci 2007, 10, 1587–1593. [Google Scholar] [CrossRef] [PubMed]
- Krüttner, S.; Traunmüller, L.; Dag, U.; Jandrasits, K.; Stepien, B.; Iyer, N.; Fradkin, L.G.; Noordermeer, J.N.; Mensh, B.D.; Keleman, K. Synaptic Orb2A bridges memory acquisition and late memory consolidation in Drosophila. Cell Rep. 2015, 11, 1953–1965. [Google Scholar] [CrossRef] [Green Version]
- Dag, U.; Lei, Z.; Le, J.Q.; Wong, A.; Bushey, D.; Keleman, K. Neuronal reactivation during post-learning sleep consolidates long-term memory in Drosophila. Elife 2019, 8, e42786. [Google Scholar] [CrossRef]
- Inami, S.; Sato, S.; Kondo, S.; Tanimoto, H.; Kitamoto, T.; Sakai, T. Environmental Light Is Required for Maintenance of Long-Term Memory in Drosophila. J. Neurosci. 2020, 40, 1427–1439. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 1 December 2021).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Li, H. Aligning Sequence Reads, Clone Sequences and Assembly Contigs with BWA-MEM. Available online: https://doi.org/10.6084/M9.FIGSHARE.963153.V1 (accessed on 1 December 2021). [CrossRef]
- Samtools. Available online: https://github.com/samtools/ (accessed on 1 December 2021).
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative Genomics Viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef] [Green Version]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef]
- PROMO 3.0. Available online: http://alggen.lsi.upc.es/cgi-bin/promo_v3/promo/promoinit.cgi?dirDB=TF_8.3 (accessed on 1 December 2021).
- Sali, A.; Blundell, T.L. Comparative protein modelling by satisfaction of spatial restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.W.; Hall, J.C. Conditioned responses in courtship behavior of normal and mutant Drosophila. Proc. Natl. Acad. Sci. USA 1979, 76, 3430–3434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgington, E. Randomization Tests; Marcel Dekker: New York, NY, USA, 1995. [Google Scholar]
- Thapa, A.; Sullivan, S.M.; Nguyen, M.Q.; Buckley, D.; Ngo, V.T.; Dada, A.O.; Blankinship, E.; Cloud, V.; Mohan, R.D. Brief freezing steps lead to robust immunofluorescence in the Drosophila nervous system. Biotechniques 2019, 67, 299–305. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhuravlev, A.V.; Ivanova, P.N.; Makaveeva, K.A.; Zakharov, G.A.; Nikitina, E.A.; Savvateeva-Popova, E.V. cd1 Mutation in Drosophila Affects Phenoxazinone Synthase Catalytic Site and Impairs Long-Term Memory. Int. J. Mol. Sci. 2022, 23, 12356. https://doi.org/10.3390/ijms232012356
Zhuravlev AV, Ivanova PN, Makaveeva KA, Zakharov GA, Nikitina EA, Savvateeva-Popova EV. cd1 Mutation in Drosophila Affects Phenoxazinone Synthase Catalytic Site and Impairs Long-Term Memory. International Journal of Molecular Sciences. 2022; 23(20):12356. https://doi.org/10.3390/ijms232012356
Chicago/Turabian StyleZhuravlev, Aleksandr V., Polina N. Ivanova, Ksenia A. Makaveeva, Gennadii A. Zakharov, Ekaterina A. Nikitina, and Elena V. Savvateeva-Popova. 2022. "cd1 Mutation in Drosophila Affects Phenoxazinone Synthase Catalytic Site and Impairs Long-Term Memory" International Journal of Molecular Sciences 23, no. 20: 12356. https://doi.org/10.3390/ijms232012356