
The Influence of Prenatal Exposure to Quetiapine Fumarate on the Development of Dopaminergic Neurons in the Ventral Midbrain of Mouse Embryos
 , ,
, ,  , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Results
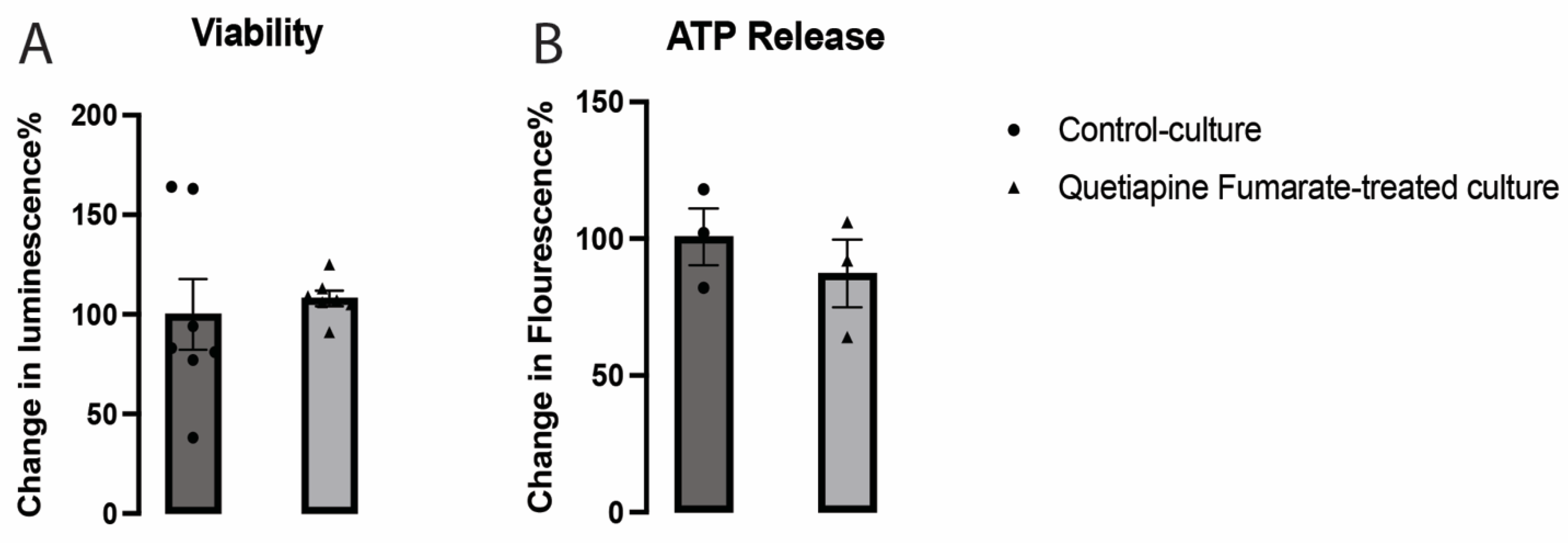
2.1. The Effects of QEPF on the Metabolic Activity of VM Neurons
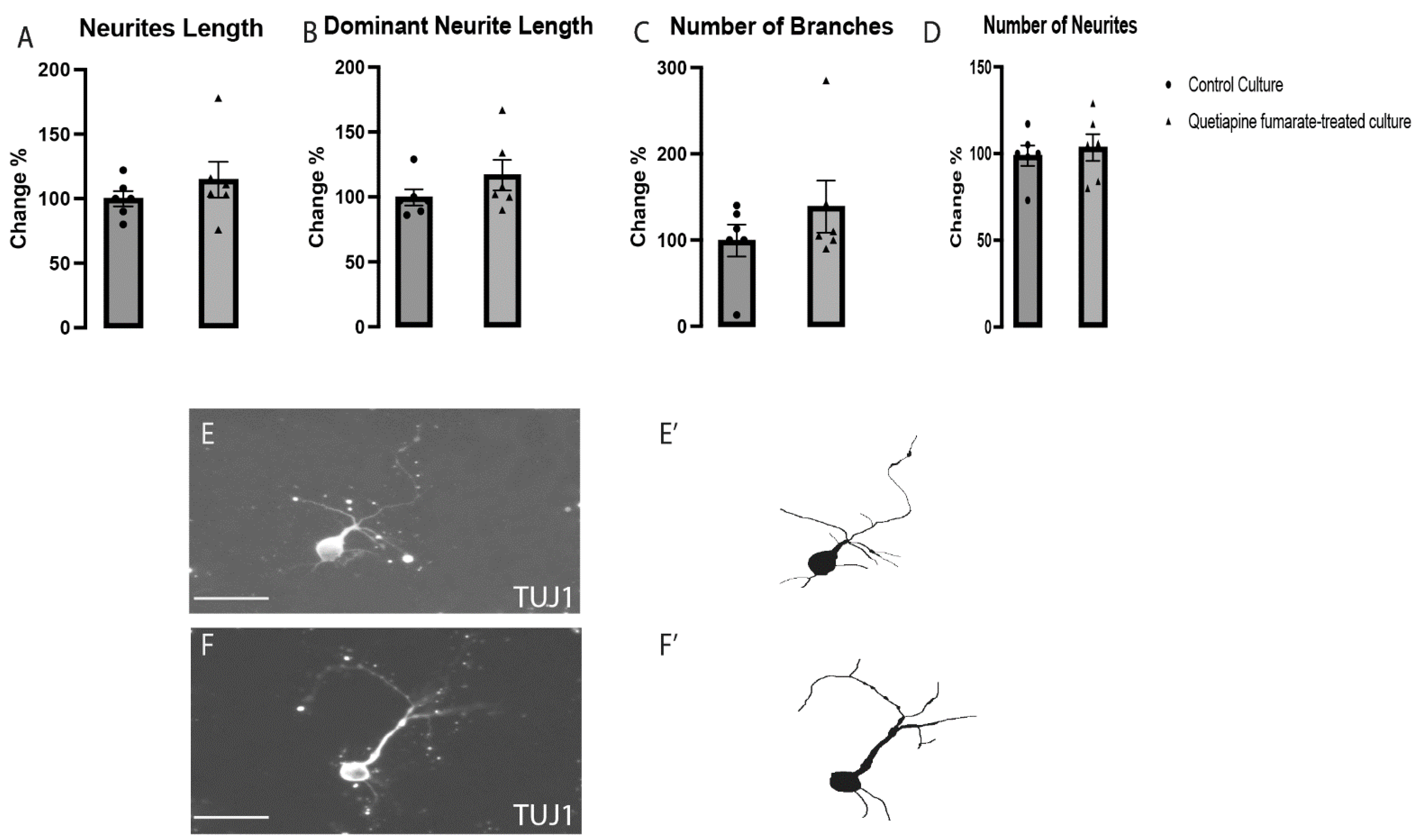
2.2. The Effects of QEPF on the Morphogenesis of Dopaminergic VM Neurons
2.3. QEPF Does Not Affect the Morphogenesis of Non-Dopaminergic VM Neurons
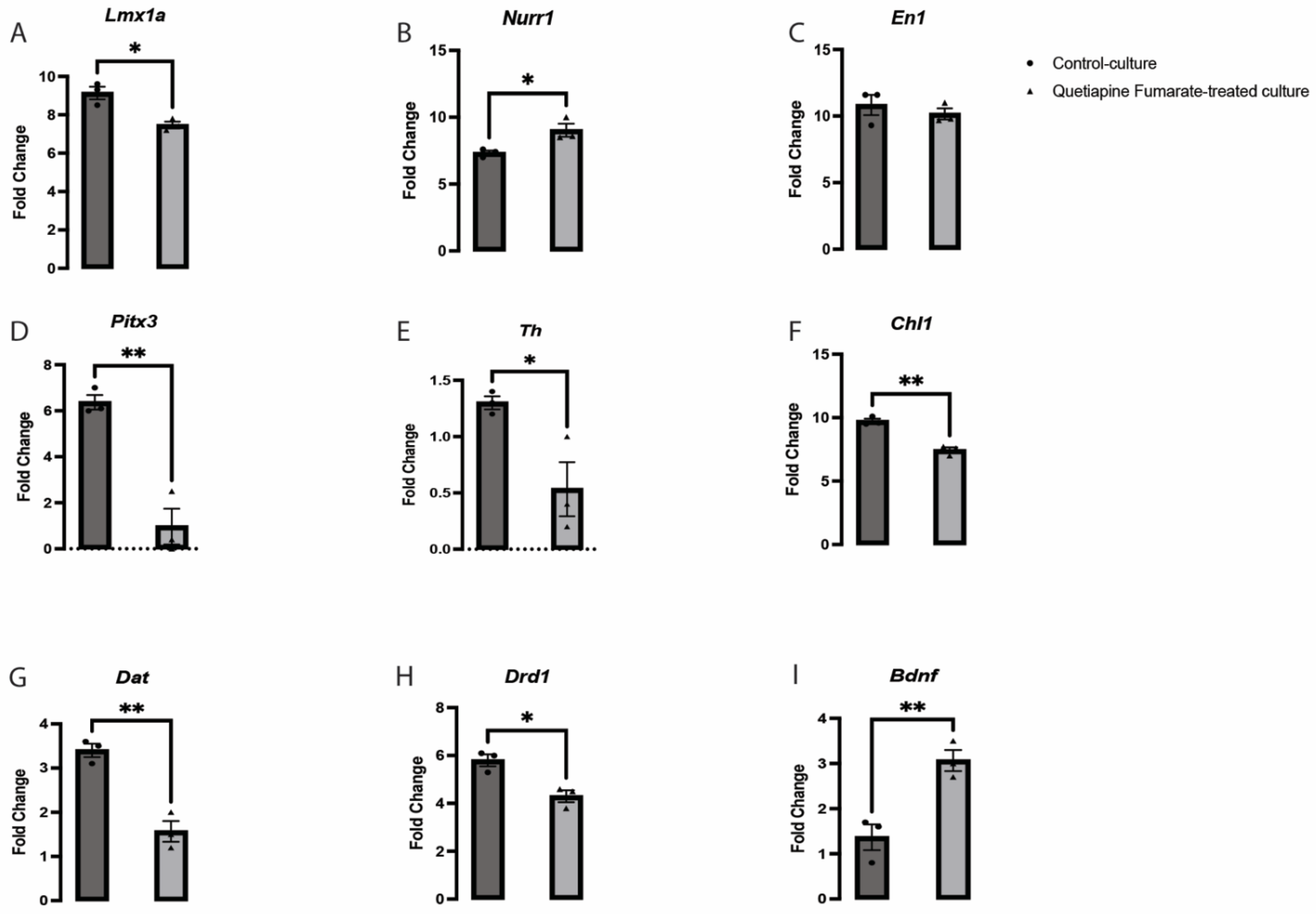
2.4. QEPF Induces Changes in the Expression of Crucial Dopaminergic-Related Genes in VM Neurons
3. Discussion
4. Materials and Methods
4.1. Isolation of Primary Mouse Embryonic VM Dopaminergic Neurons
4.2. Three-Dimensional Neuronal Cell Culture and QEPF Treatment
4.3. VM Neuronal Viability and ATP Release Assessment
4.4. Immunocytochemistry
4.5. Morphogenetic Analysis
4.6. Quantitative PCR
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Toh, S.; Li, Q.; Cheetham, T.C.; Cooper, W.O.; Davis, R.L.; Dublin, S.; Hammad, T.A.; Li, D.-K.; Pawloski, P.A.; Pinheiro, S.P. Prevalence and trends in the use of antipsychotic medications during pregnancy in the US, 2001–2007: A population-based study of 585,615 deliveries. Arch. Women’s Ment. Health 2013, 16, 149–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leucht, S.; Corves, C.; Arbter, D.; Engel, R.R.; Li, C.; Davis, J.M. Second-generation versus first-generation antipsychotic drugs for schizophrenia: A meta-analysis. Lancet 2009, 373, 31–41. [Google Scholar] [CrossRef]
- Singh, K.; Tripathi, N. Prenatal exposure to a novel antipsychotic quetiapine: Impact on neuro-architecture, apoptotic neurodegeneration in fetal hippocampus and cognitive impairment in young rats. Int. J. Dev. Neurosci. 2015, 42, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Abel, K.M. Fetal antipsychotic exposure in a changing landscape: Seeing the future. Br. J. Psychiatry 2013, 202, 321–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ennis, Z.N.; Damkier, P. Pregnancy exposure to olanzapine, quetiapine, risperidone, aripiprazole and risk of congenital malformations. A systematic review. Basic Clin. Pharmacol. Toxicol. 2015, 116, 315–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peluso, M.J.; Lewis, S.n.W.; Barnes, T.R.; Jones, P.B. Extrapyramidal motor side-effects of first-and second-generation antipsychotic drugs. Br. J. Psychiatry 2012, 200, 387–392. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.; Singh, M. Effect of prenatal haloperidol exposure on behavioral alterations in rats. Neurotoxicol. Teratol. 2002, 24, 497–502. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, L.; Pitts, D.K. Prenatal haloperidol reduces the number of active midbrain dopamine neurons in rat offspring. Neurotoxicology Teratol. 1996, 18, 49–57. [Google Scholar] [CrossRef]
- Anderson, K.N.; Ailes, E.C.; Lind, J.N.; Broussard, C.S.; Bitsko, R.H.; Friedman, J.M.; Bobo, W.V.; Reefhuis, J.; Tinker, S.C. National Birth Defects Prevention Study. Atypical antipsychotic use during pregnancy and birth defect risk: National Birth Defects Prevention Study, 1997-2011. Schizophr Res. 2020, 215, 81–88. [Google Scholar] [CrossRef]
- Hanley, G.E.; Mintzes, B. Patterns of psychotropic medicine use in pregnancy in the United States from 2006 to 2011 among women with private insurance. BMC Pregnancy Childbirth 2014, 14, 242. [Google Scholar] [CrossRef]
- Kennedy, D.; Eamus, M.; Hill, M.; Oei, J.L. Review of calls to an A ustralian teratogen information service regarding psychotropic medications over a 12-year period. Aust. New Zealand J. Obstet. Gynaecol. 2013, 53, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Soeiro-de-Souza, M.G.; Dias, V.V.; Missio, G.; Balanzá-Martinez, V.; Valiengo, L.; Carvalho, A.F.; Moreno, R.A. Role of quetiapine beyond its clinical efficacy in bipolar disorder: From neuroprotection to the treatment of psychiatric disorders. Exp. Ther. Med. 2015, 9, 643–652. [Google Scholar] [CrossRef] [Green Version]
- Larsen, E.; Damkier, P.; Pedersen, L.; Fenger-Gron, J.; Mikkelsen, R.; Nielsen, R.; Linde, V.; Knudsen, H.; Skaarup, L.; Videbech, P. Use of psychotropic drugs during pregnancy and breast-feeding. Acta Psychiatr. Scand. 2015, 132, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Hauser, C.A.; Zhang, S. Designer self-assembling peptide nanofiber biological materials. Chem. Soc. Rev. 2010, 39, 2780–2790. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Pan, F.; Xu, H.; Yaseen, M.; Shan, H.; Hauser, C.A.; Zhang, S.; Lu, J.R. Molecular self-assembly and applications of designer peptide amphiphiles. Chem. Soc. Rev. 2010, 39, 3480–3498. [Google Scholar] [CrossRef] [PubMed]
- Susapto, H.H.; Alhattab, D.; Abdelrahman, S.; Khan, Z.; Alshehri, S.; Kahin, K.; Ge, R.; Moretti, M.; Emwas, A.-H.; Hauser, C.A. Ultrashort peptide bioinks support automated printing of large-scale constructs assuring long-term survival of printed tissue constructs. Nano Lett. 2021, 21, 2719–2729. [Google Scholar] [CrossRef]
- Money, K.M.; Stanwood, G.D. Developmental origins of brain disorders: Roles for dopamine. Front. Cell. Neurosci. 2013, 7, 260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silverstone, P.H.; Lalies, M.D.; Hudson, A. Quetiapine and buspirone both elevate cortical levels of noradrenaline and dopamine in vivo, but do not have synergistic effects. Front. Psychiatry 2012, 3, 82. [Google Scholar] [CrossRef] [Green Version]
- Pira, L.; Mongeau, R.; Pani, L. The atypical antipsychotic quetiapine increases both noradrenaline and dopamine release in the rat prefrontal cortex. Eur. J. Pharmacol. 2004, 504, 61–64. [Google Scholar] [CrossRef]
- Burbach, J.P.H.; Smits, S.; Smidt, M.P. Transcription factors in the development of midbrain dopamine neurons. Ann. N. Y. Acad. Sci. 2003, 991, 61–68. [Google Scholar] [CrossRef]
- Van den Heuvel, D.M.; Pasterkamp, R.J. Getting connected in the dopamine system. Prog. Neurobiol. 2008, 85, 75–93. [Google Scholar] [CrossRef] [PubMed]
- Parish, C.L.; Thompson, L.H. Modulating Wnt signaling to improve cell replacement therapy for Parkinson’s disease. J. Mol. Cell Biol. 2014, 6, 54–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doucet-Beaupré, H.; Gilbert, C.; Profes, M.S.; Chabrat, A.; Pacelli, C.; Giguère, N.; Rioux, V.; Charest, J.; Deng, Q.; Laguna, A. Lmx1a and Lmx1b regulate mitochondrial functions and survival of adult midbrain dopaminergic neurons. Proc. Natl. Acad. Sci. USA 2016, 113, E4387–E4396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobert, O.; Westphal, H. Functions of LIM-homeobox genes. Trends Genet. 2000, 16, 75–83. [Google Scholar] [CrossRef]
- Bergman, O.; Håkansson, A.; Westberg, L.; Belin, A.C.; Sydow, O.; Olson, L.; Holmberg, B.; Fratiglioni, L.; Bäckman, L.; Eriksson, E. Do polymorphisms in transcription factors LMX1A and LMX1B influence the risk for Parkinson’s disease? J. Neural Transm. 2009, 116, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Doucet-Beaupré, H.; Lévesque, M. The role of developmental transcription factors in adult midbrain dopaminergic neurons. Neurosciences 2013, 1, 3. [Google Scholar]
- Hoekstra, E.J.; von Oerthel, L.; van der Heide, L.P.; Kouwenhoven, W.M.; Veenvliet, J.V.; Wever, I.; Jin, Y.-R.; Yoon, J.K.; van der Linden, A.J.; Holstege, F.C. Lmx1a encodes a rostral set of mesodiencephalic dopaminergic neurons marked by the Wnt/B-catenin signaling activator R-spondin 2. PLoS ONE 2013, 8, e74049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsanie, W.; Penna, V.; Schachner, M.; Thompson, L.; Parish, C. Homophilic binding of the neural cell adhesion molecule CHL1 regulates development of ventral midbrain dopaminergic pathways. Sci. Rep. 2017, 7, 9368. [Google Scholar] [CrossRef]
- Althobaiti, Y.S.; Almutairi, F.M.; Alshehri, F.S.; Altowairqi, E.; Marghalani, A.M.; Alghorabi, A.A.; Alsanie, W.F.; Gaber, A.; Alsaab, H.O.; Almalki, A.H. Involvement of the dopaminergic system in the reward-related behavior of pregabalin. Sci. Rep. 2021, 11, 10577. [Google Scholar] [CrossRef]
- Ahearn, E.P.; Mussey, M.; Johnson, C.; Krohn, A.; Krahn, D. Quetiapine as an adjunctive treatment for post-traumatic stress disorder: An 8-week open-label study. Int. Clin. Psychopharmacol. 2006, 21, 29–33. [Google Scholar] [CrossRef]
- Komossa, K.; Depping, A.M.; Gaudchau, A.; Kissling, W.; Leucht, S. Second-generation antipsychotics for major depressive disorder and dysthymia. Cochrane Database Syst. Rev. 2010, 12, 1465–1858. [Google Scholar] [CrossRef] [PubMed]
- Stathis, S.; Martin, G.; McKenna, J.G. A preliminary case series on the use of quetiapine for posttraumatic stress disorder in juveniles within a youth detention center. J. Clin. Psychopharmacol. 2005, 25, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-n.; Liu, G.-h.; Zhang, R.-g.; Xue, F.; Wu, D.; Chen, Y.-c.; Peng, Y.; Peng, Z.-w.; Tan, Q.-r. Quetiapine ameliorates schizophrenia-like behaviors and protects myelin integrity in cuprizone intoxicated mice: The involvement of notch signaling pathway. Int. J. Neuropsychopharmacol. 2016, 19, pyv088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzman, F. Mechanism of Action of Quetiapine. Available online: https://psychopharmacologyinstitute.com/publication/mechanism-of-action-of-quetiapine-2109 (accessed on 1 April 2022).
- Niederkofler, V.; Asher, T.E.; Dymecki, S.M. Functional interplay between dopaminergic and serotonergic neuronal systems during development and adulthood. ACS Chem. Neurosci. 2015, 6, 1055–1070. [Google Scholar] [CrossRef] [Green Version]
- Kondo, M.; Tajinda, K.; Colantuoni, C.; Hiyama, H.; Seshadri, S.; Huang, B.; Pou, S.; Furukori, K.; Hookway, C.; Jaaro-Peled, H. Unique pharmacological actions of atypical neuroleptic quetiapine: Possible role in cell cycle/fate control. Transl. Psychiatry 2013, 3, e243. [Google Scholar] [CrossRef] [Green Version]
- Rosengarten, H.; Quartermain, D. Effect of prenatal administration of haloperidol, risperidone, quetiapine and olanzapine on spatial learning and retention in adult rats. Pharmacol. Biochem. Behav. 2002, 72, 575–579. [Google Scholar] [CrossRef]
- Wang, M.; Ling, K.-H.; Tan, J.J.; Lu, C.-B. Development and differentiation of midbrain dopaminergic neuron: From bench to bedside. Cells 2020, 9, 1489. [Google Scholar] [CrossRef]
- Yin, M.; Liu, S.; Yin, Y.; Li, S.; Li, Z.; Wu, X.; Zhang, B.; Ang, S.-L.; Ding, Y.; Zhou, J. Ventral mesencephalon-enriched genes that regulate the development of dopaminergic neurons in vivo. J. Neurosci. 2009, 29, 5170–5182. [Google Scholar] [CrossRef] [Green Version]
- Volpicelli, F.; De Gregorio, R.; Pulcrano, S.; Perrone-Capano, C.; di Porzio, U.; Bellenchi, G.C. Direct regulation of Pitx3 expression by Nurr1 in culture and in developing mouse midbrain. PLoS ONE 2012, 7, e30661. [Google Scholar]
- Gyllborg, D.; Ahmed, M.; Toledo, E.M.; Theofilopoulos, S.; Yang, S.; Arenas, E. The matricellular protein R-Spondin 2 promotes midbrain dopaminergic neurogenesis and differentiation. Stem Cell Rep. 2018, 11, 651–664. [Google Scholar] [CrossRef] [Green Version]
- DeVane, C.L.; Nemeroff, C.B. Clinical pharmacokinetics of quetiapine. Clin. Pharmacokinet. 2001, 40, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Kim, C.H.; Kim, K.S. Lmx1a regulates dopamine transporter gene expression during ES cell differentiation and mouse embryonic development. J. Neurochem. 2012, 122, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Doucet-Beaupré, H.; Ang, S.-L.; Lévesque, M. Cell fate determination, neuronal maintenance and disease state: The emerging role of transcription factors Lmx1a and Lmx1b. FEBS Lett. 2015, 589, 3727–3738. [Google Scholar] [CrossRef] [Green Version]
- Hyman, C.; Hofer, M.; Barde, Y.-A.; Juhasz, M.; Yancopoulos, G.D.; Squinto, S.P.; Lindsay, R.M. BDNF is a neurotrophic factor for dopaminergic neurons of the substantia nigra. Nature 1991, 350, 230–232. [Google Scholar] [CrossRef] [PubMed]
- Lühder, F.; Gold, R.; Flügel, A.; Linker, R.A. Brain-derived neurotrophic factor in neuroimmunology: Lessons learned from multiple sclerosis patients and experimental autoimmune encephalomyelitis models. Arch. Immunol. Et Ther. Exp. 2013, 61, 95–105. [Google Scholar] [CrossRef]
- Xu, H.; Qing, H.; Lu, W.; Keegan, D.; Richardson, J.S.; Chlan-Fourney, J.; Li, X.-M. Quetiapine attenuates the immobilization stress-induced decrease of brain-derived neurotrophic factor expression in rat hippocampus. Neurosci. Lett. 2002, 321, 65–68. [Google Scholar] [CrossRef]
- Park, S.-W.; Lee, S.-K.; Kim, J.-M.; Yoon, J.-S.; Kim, Y.-H. Effects of quetiapine on the brain-derived neurotrophic factor expression in the hippocampus and neocortex of rats. Neurosci. Lett. 2006, 402, 25–29. [Google Scholar] [CrossRef]
- Fumagalli, F.; Molteni, R.; Bedogni, F.; Gennarelli, M.; Perez, J.; Racagni, G.; Riva, M.A. Quetiapine regulates FGF-2 and BDNF expression in the hippocampus of animals treated with MK-801. Neuroreport 2004, 15, 2109–2112. [Google Scholar] [CrossRef]
- Park, S.W.; Lee, C.H.; Cho, H.Y.; Seo, M.K.; Lee, J.G.; Lee, B.J.; Seol, W.; Kee, B.S.; Kim, Y.H. Effects of antipsychotic drugs on the expression of synaptic proteins and dendritic outgrowth in hippocampal neuronal cultures. Synapse 2013, 67, 224–234. [Google Scholar] [CrossRef]
- Heng, X.; Jin, G.; Zhang, X.; Yang, D.; Zhu, M.; Fu, S.; Li, X.; Le, W. Nurr1 regulates Top IIβ and functions in axon genesis of mesencephalic dopaminergic neurons. Mol. Neurodegener. 2012, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Kadkhodaei, B.; Alvarsson, A.; Schintu, N.; Ramsköld, D.; Volakakis, N.; Joodmardi, E.; Yoshitake, T.; Kehr, J.; Decressac, M.; Björklund, A. Transcription factor Nurr1 maintains fiber integrity and nuclear-encoded mitochondrial gene expression in dopamine neurons. Proc. Natl. Acad. Sci. USA 2013, 110, 2360–2365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegarty, S.V.; Sullivan, A.M.; O’keeffe, G.W. Midbrain dopaminergic neurons: A review of the molecular circuitry that regulates their development. Dev. Biol. 2013, 379, 123–138. [Google Scholar] [CrossRef]
- Abdollahi, M.; Fahnestock, M. Nurr1 Is Not an Essential Regulator of BDNF in Mouse Cortical Neurons. Int. J. Mol. Sci. 2022, 23, 6853. [Google Scholar] [CrossRef] [PubMed]
- Alsanie, W.F.; Bahri, O.A.; Habeeballah, H.H.; Alhomrani, M.; Almehmadi, M.M.; Alsharif, K.; Felemban, E.M.; Althobaiti, Y.S.; Almalki, A.H.; Alsaab, H.O. Generating homogenous cortical preplate and deep-layer neurons using a combination of 2D and 3D differentiation cultures. Sci. Rep. 2020, 10, 6272. [Google Scholar] [CrossRef] [Green Version]
- Handley, S.A.; Bowskill, S.V.; Patel, M.X.; Flanagan, R.J. Plasma quetiapine in relation to prescribed dose and other factors: Data from a therapeutic drug monitoring service, 2000–2011. Ther. Adv. Psychopharmacol. 2013, 3, 129–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasselstrøm, J.; Linnet, K. Quetiapine serum concentrations in psychiatric patients: The influence of comedication. Ther. Drug Monit. 2004, 26, 486–491. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence (5′ to 3′) | |
|---|---|---|
| Gapdh | Forward-primer: Reverse-primer: | TGA AGG TCG GAG TCA ACG GA CCA ATT GAT GAC AAG CTT CCC G |
| β-actin | Forward-primer: Reverse-primer | GATTACTGCTCTGGCTCCTAGC GACTCATCGTACTCCTGCTTGC |
| Th | Forward-primer: Reverse-primer: | TGA AGG AAC GGA CTG GCT TC GAG TGC ATA GGT GAG GAG GC |
| Nurr1 | Forward-primer: Reverse-primer: | GAC CAG GAC CTG CTT TTT GA ACC CCA TTG CAA AAG ATG AG |
| Lmx1a | Forward-primer: Reverse-primer: | GAG ACC ACC TGC TTC TAC CG GCA CGC ATG ACA AAC TCA TT |
| En1 | Forward-primer: Reverse-primer: | TCA CAG CAA CCC CTA GTG TG CGC TTG TCT TCC TTC TCG TT |
| Pitx3 | Forward-primer: Reverse-primer: | CAT GGA GTT TGG GCT GCT TG CCT TCT CCG AGT CAC TGT GC |
| Chl1 | Forward-primer: Reverse-primer: | TGG AAT TGC CAT TAT GTG GA CAC CTG CAC GTA TGA CTG CT |
| Dat | Forward-primer: Reverse-primer: | TTG CAG CTG GCA CAT CTA TC ATG CTG ACC ACG ACC ACA TA |
| Drd1 | Forward-primer: Reverse-primer: | CTC AAC AAC ACA GAC CAG AAT GAA CGA GAC GAT GGA GGA |
| Bdnf | Forward-primer: Reverse-primer: | ACT ATG GTT ATT TCA TAC TTC GGT T CCA TTC ACG CTC TCC AGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alsanie, W.F.; Abdelrahman, S.; Alhomrani, M.; Gaber, A.; Alosimi, E.A.; Habeeballah, H.; Alkhatabi, H.A.; Felimban, R.I.; Hauser, C.A.E.; Tayeb, H.H.; et al. The Influence of Prenatal Exposure to Quetiapine Fumarate on the Development of Dopaminergic Neurons in the Ventral Midbrain of Mouse Embryos. Int. J. Mol. Sci. 2022, 23, 12352. https://doi.org/10.3390/ijms232012352
Alsanie WF, Abdelrahman S, Alhomrani M, Gaber A, Alosimi EA, Habeeballah H, Alkhatabi HA, Felimban RI, Hauser CAE, Tayeb HH, et al. The Influence of Prenatal Exposure to Quetiapine Fumarate on the Development of Dopaminergic Neurons in the Ventral Midbrain of Mouse Embryos. International Journal of Molecular Sciences. 2022; 23(20):12352. https://doi.org/10.3390/ijms232012352
Chicago/Turabian StyleAlsanie, Walaa F., Sherin Abdelrahman, Majid Alhomrani, Ahmed Gaber, Ebtisam Abdulah Alosimi, Hamza Habeeballah, Heba A. Alkhatabi, Raed I. Felimban, Charlotte A. E. Hauser, Hossam H. Tayeb, and et al. 2022. "The Influence of Prenatal Exposure to Quetiapine Fumarate on the Development of Dopaminergic Neurons in the Ventral Midbrain of Mouse Embryos" International Journal of Molecular Sciences 23, no. 20: 12352. https://doi.org/10.3390/ijms232012352