
Punicalagin Protects against the Development of Methotrexate-Induced Hepatotoxicity in Mice via Activating Nrf2 Signaling and Decreasing Oxidative Stress, Inflammation, and Cell Death
 , , , , , and
, , , , , and 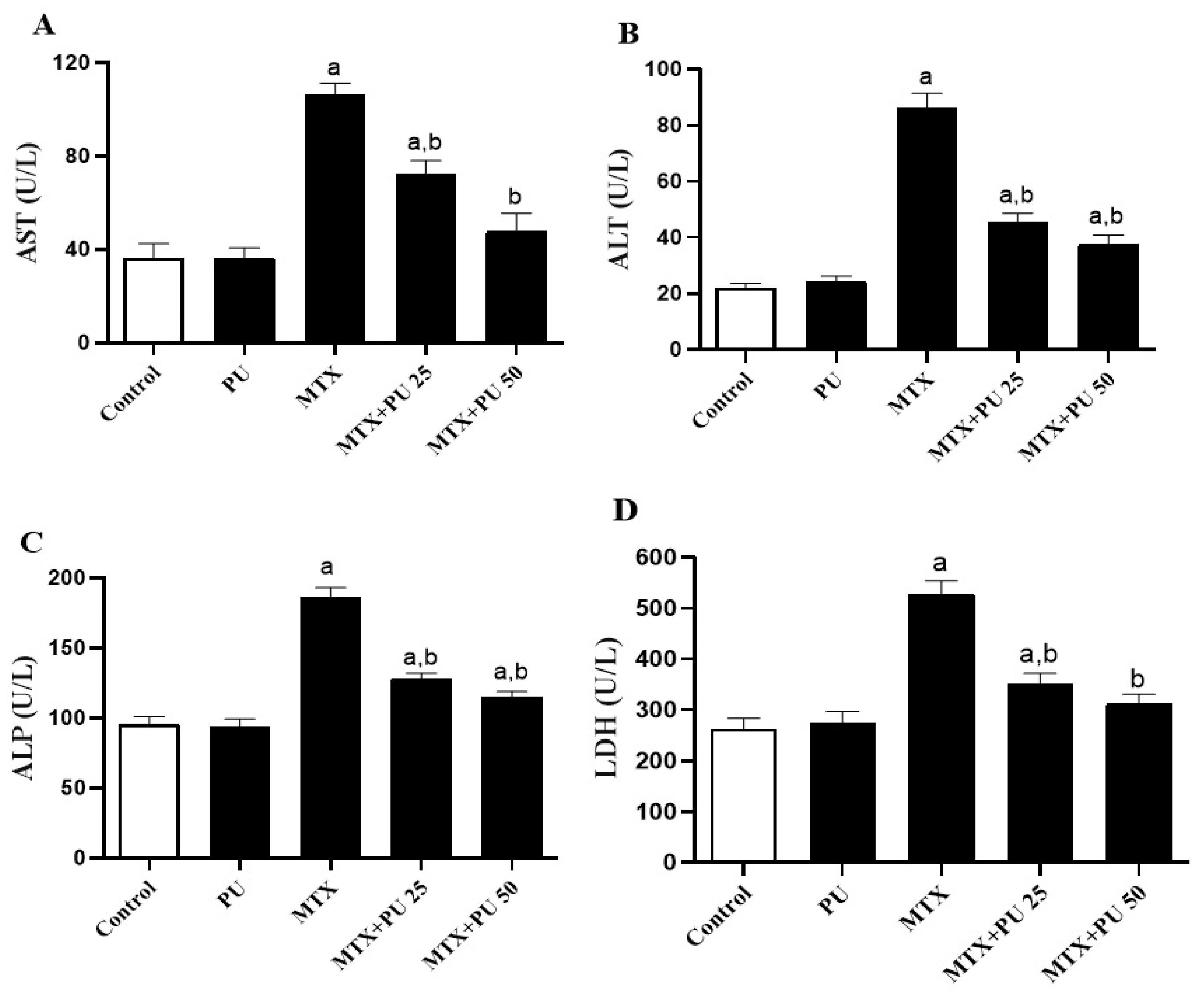{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
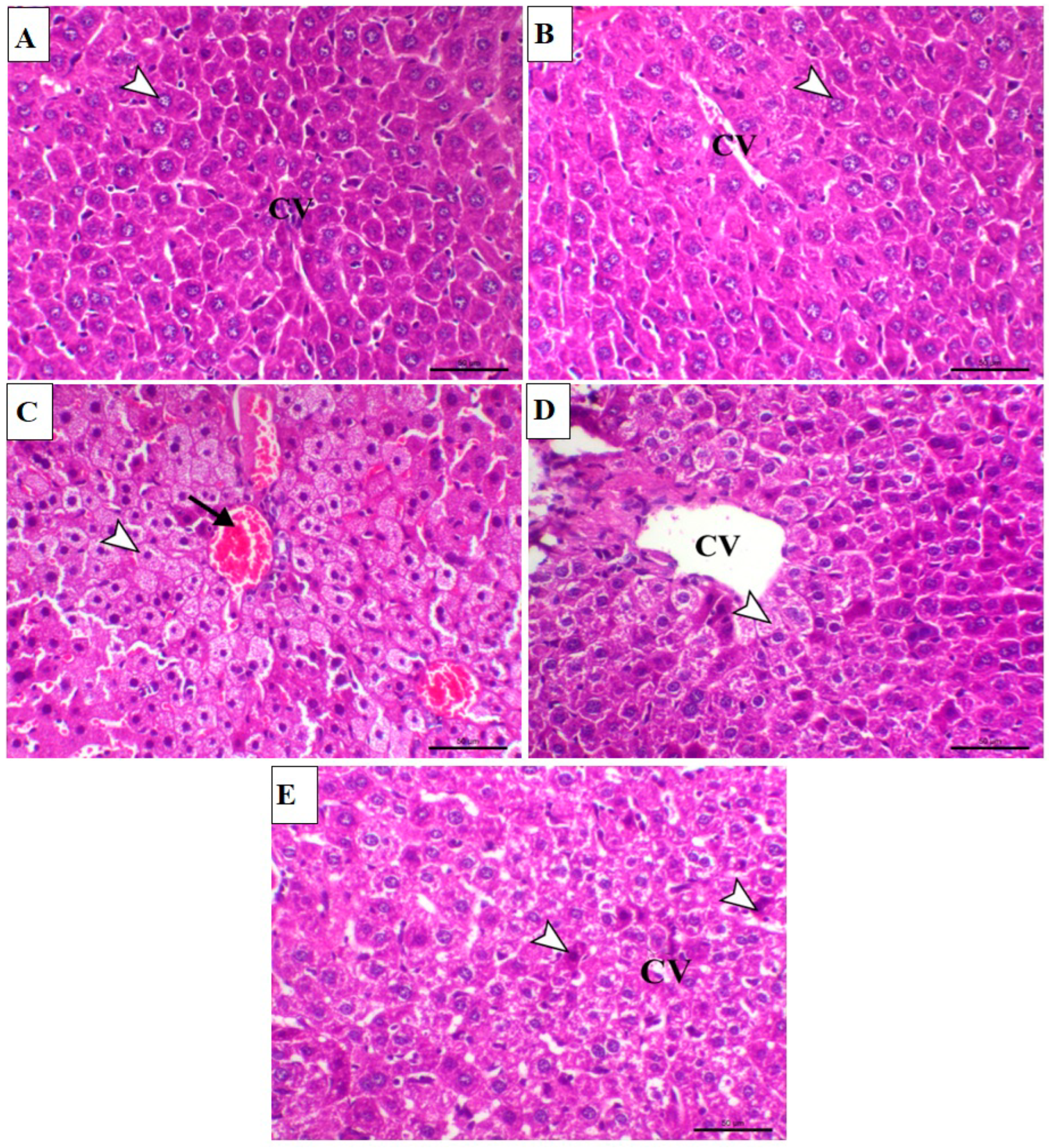
2.1. PU Prevents MTX-Induced Liver Injury in Mice
2.2. PU Attenuates Oxidative Stress and Enhances Antioxidants Defenses in Liver of MTX-Treated Mice
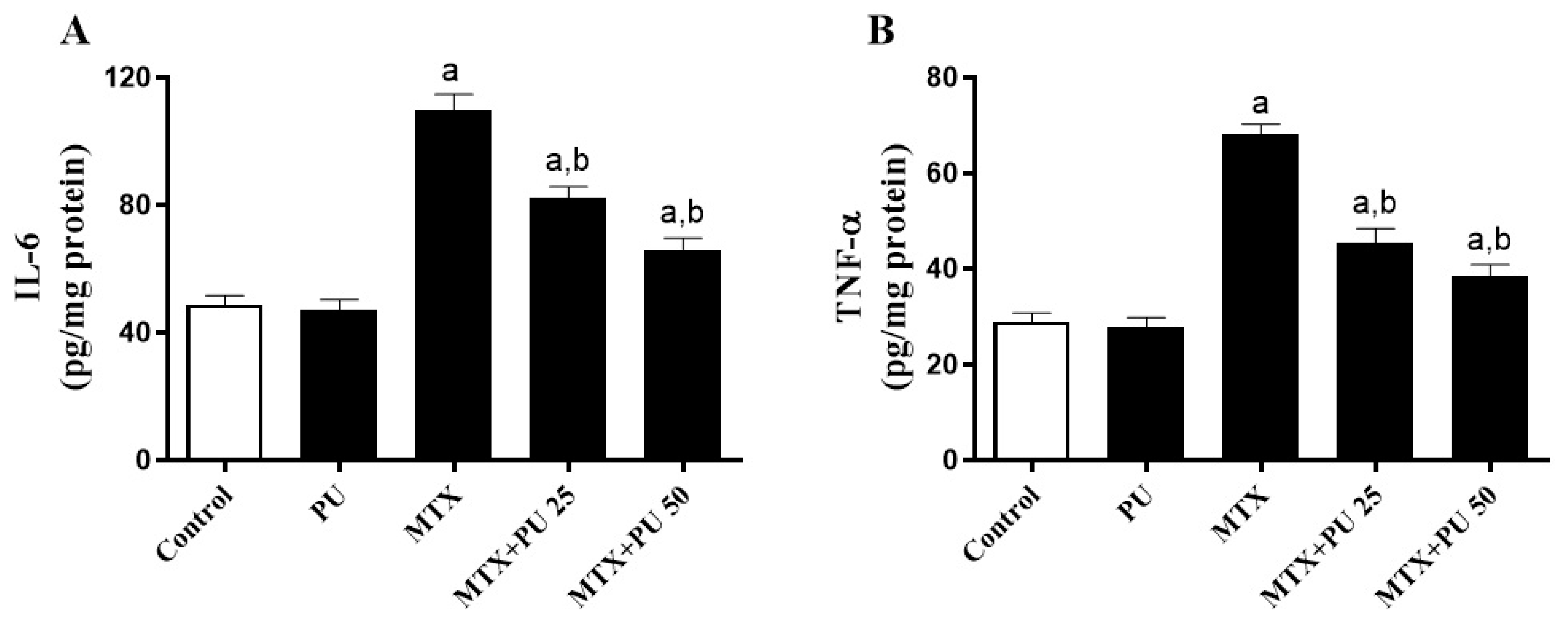
2.3. PU Suppresses the MTX-Induced Hepatic Inflammation in Mice
2.4. PU Mitigates the MTX-Induced Apoptosis in the Liver
2.5. PU Upregulates Hepatic Nrf2/Heme Oxygenase 1 (HO-1) in MTX-Treated Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Design
4.3. Estimation of Markers of Liver Function
4.4. Assessment of Oxidative Stress Markers and Antioxidant Contents in Liver Tissues
4.5. Estimation of Pro-Inflammatory Cytokines in Liver
4.6. Histological Examination of Liver Sections
4.7. Immunohistochemistry
4.8. Analysis of Data
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Katarey, D.; Verma, S. Drug-induced liver injury. Clin. Med. 2016, 16, s104. [Google Scholar] [CrossRef] [PubMed]
- Andrade, R.J.; Chalasani, N.; Björnsson, E.S.; Suzuki, A.; Kullak-Ublick, G.A.; Watkins, P.B.; Devarbhavi, H.; Merz, M.; Lucena, M.I.; Kaplowitz, N. Drug-induced liver injury. Nat. Rev. Dis. Prim. 2019, 5, 1–22. [Google Scholar] [CrossRef] [Green Version]
- David, S.; Hamilton, J.P. Drug-induced liver injury. US Gastroenterol. Hepatol. Rev. 2010, 6, 73. [Google Scholar] [PubMed]
- Puig, L. Methotrexate: New therapeutic approaches. Actas Dermo-Sifiliográficas 2014, 105, 583–589. [Google Scholar] [CrossRef]
- Sahindokuyucu-Kocasari, F.; Akyol, Y.; Ozmen, O.; Erdemli-Kose, S.; Garli, S. Apigenin alleviates methotrexate-induced liver and kidney injury in mice. Hum. Exp. Toxicol. 2021, 40, 1721–1731. [Google Scholar] [CrossRef] [PubMed]
- Goljan-Geremek, A.; Bednarek, M.; Franczuk, M.; Puścińska, E.; Nowiński, A.; Czystowska, M.; Kamiński, D.; Korzybski, D.; Stokłosa, A.; Kowalska, A. Methotrexate as a single agent for treating pulmonary sarcoidosis: A single centre real-life prospective study. Adv. Respir. Med. 2014, 82, 518–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kremer, J.M. Toward a better understanding of methotrexate. Arthritis Rheum. 2004, 50, 1370–1382. [Google Scholar] [CrossRef] [PubMed]
- Howard, S.C.; McCormick, J.; Pui, C.H.; Buddington, R.K.; Harvey, R.D. Preventing and managing toxicities of high-dose methotrexate. Oncologist 2016, 21, 1471–1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Zhou, H.; Liu, L. Side effects of methotrexate therapy for rheumatoid arthritis: A systematic review. Eur. J. Med. Chem. 2018, 158, 502–516. [Google Scholar] [CrossRef] [PubMed]
- Dalaklioglu, S.; Genc, G.; Aksoy, N.; Akcit, F.; Gumuslu, S. Resveratrol ameliorates methotrexate-induced hepatotoxicity in rats via inhibition of lipid peroxidation. Hum. Exp. Toxicol. 2013, 32, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Yélamos, O.; Català, A.; Vilarrasa, E.; Roé, E.; Puig, L. Acute severe methotrexate toxicity in patients with psoriasis: A case series and discussion. Dermatology 2014, 229, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Ahmadzadeh, A.; Zamani, N.; Hassanian-Moghaddam, H.; Hadeiy, S.K.; Parhizgar, P. Acute versus chronic methotrexate poisoning; a cross-sectional study. BMC Pharmacol. Toxicol. 2019, 20, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bannwarth, B.; Péhourcq, F.; Schaeverbeke, T.; Dehais, J. Clinical pharmacokinetics of low-dose pulse methotrexate in rheumatoid arthritis. Clin. Pharmacokinet. 1996, 30, 194–210. [Google Scholar] [CrossRef] [PubMed]
- Maksimovic, V.; Pavlovic-Popovic, Z.; Vukmirovic, S.; Cvejic, J.; Mooranian, A.; Al-Salami, H.; Mikov, M.; Golocorbin-Kon, S. Molecular mechanism of action and pharmacokinetic properties of methotrexate. Mol. Biol. Rep. 2020, 47, 4699–4708. [Google Scholar] [CrossRef] [PubMed]
- Ezhilarasan, D. Hepatotoxic potentials of methotrexate: Understanding the possible toxicological molecular mechanisms. Toxicology 2021, 458, 152840. [Google Scholar] [CrossRef]
- Dalvie, D.; Di, L. Aldehyde oxidase and its role as a drug metabolizing enzyme. Pharmacol. Ther. 2019, 201, 137–180. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Mori, N. Involvement of multiple transporters-mediated transports in mizoribine and methotrexate pharmacokinetics. Pharmaceuticals 2012, 5, 802–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widemann, B.C.; Adamson, P.C. Understanding and managing methotrexate nephrotoxicity. Oncologist 2006, 11, 694–703. [Google Scholar] [CrossRef] [Green Version]
- Vardi, N.; Parlakpinar, H.; Cetin, A.; Erdogan, A.; Cetin Ozturk, I. Protective effect of β-carotene on methotrexate–induced oxidative liver damage. Toxicol. Pathol. 2010, 38, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Heidari, R.; Ahmadi, A.; Mohammadi, H.; Ommati, M.M.; Azarpira, N.; Niknahad, H. Mitochondrial dysfunction and oxidative stress are involved in the mechanism of methotrexate-induced renal injury and electrolytes imbalance. Biomed. Pharmacother. 2018, 107, 834–840. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.; Rashid, S.; Nafees, S.; Hasan, S.K.; Shahid, A.; Majed, F.; Sultana, S. Protective effect of Chlorogenic acid against methotrexate induced oxidative stress, inflammation and apoptosis in rat liver: An experimental approach. Chem.-Biol. Interact. 2017, 272, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.; Rashid, S.; Nafees, S.; Hasan, S.K.; Sultana, S. Beneficial effects of Chrysin against Methotrexate-induced hepatotoxicity via attenuation of oxidative stress and apoptosis. Mol. Cell. Biochem. 2014, 385, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Clarke, J.L.; Murray, J.B.; Park, B.K.; Copple, I.M. Roles of Nrf2 in drug and chemical toxicity. Curr. Opin. Toxicol. 2016, 1, 104–110. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, A.M.; Alexander, M.Y.; Tutar, Y.; Wilkinson, F.L.; Venditti, A. Oxidative stress in metabolic disorders and drug-induced injury: The potential role of Nrf2 and PPARs activators. Oxidative Med. Cell. Longev. 2017, 2017, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemeida, R.; Mohafez, O.M. Curcumin attenuates methotraxate-induced hepatic oxidative damage in rats. J. Egypt. Natl. Cancer Inst. 2008, 20, 141–148. [Google Scholar]
- Pradhan, A.; Sengupta, S.; Sengupta, R.; Chatterjee, M. Attenuation of methotrexate induced hepatotoxicity by epigallocatechin 3-gallate. Drug Chem. Toxicol. 2022, 1–9. [Google Scholar] [CrossRef]
- Goudarzi, M.; Basir, Z.; Malayeri, A.; Nesari, A.; Zaeemzadeh, N. Zingerone Attenuates Methotrexate-Induced Hepatotoxicity in Rats. Jundishapur J. Nat. Pharm. Prod. 2021, 17, e118745. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Hussein, O.E.; Hozayen, W.G.; Abd El-Twab, S.M. Methotrexate hepatotoxicity is associated with oxidative stress, and down-regulation of PPARγ and Nrf2: Protective effect of 18β-Glycyrrhetinic acid. Chem. -Biol. Interact. 2017, 270, 59–72. [Google Scholar] [CrossRef]
- Ge, S.; Duo, L.; Wang, J.; Yang, J.; Li, Z.; Tu, Y. A unique understanding of traditional medicine of pomegranate, Punica granatum L. and its current research status. J. Ethnopharmacol. 2021, 271, 113877. [Google Scholar] [CrossRef]
- Ruan, J.-H.; Li, J.; Adili, G.; Sun, G.-Y.; Abuduaini, M.; Abdulla, R.; Maiwulanjiang, M.; Aisa, H.A. Phenolic Compounds and Bioactivities from Pomegranate (Punica granatum L.) Peels. J. Agric. Food Chem. 2022, 70, 3678–3686. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Hosseini, M.S.; Daneshvar Hakimi Meybodi, N.; Peijnenburg, W. Mitigation of the effect of drought on growth and yield of pomegranates by foliar spraying of different sizes of selenium nanoparticles. J. Sci. Food Agric. 2021, 101, 5202–5213. [Google Scholar] [CrossRef] [PubMed]
- Fahmy, H.A.; Farag, M.A. Ongoing and potential novel trends of pomegranate fruit peel; a comprehensive review of its health benefits and future perspectives as nutraceutical. J. Food Biochem. 2022, 46, e14024. [Google Scholar] [CrossRef] [PubMed]
- Cao, K.; Xu, J.; Pu, W.; Dong, Z.; Sun, L.; Zang, W.; Gao, F.; Zhang, Y.; Feng, Z.; Liu, J. Punicalagin, an active component in pomegranate, ameliorates cardiac mitochondrial impairment in obese rats via AMPK activation. Sci. Rep. 2015, 5, 14014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aladaileh, S.H.; Al-Swailmi, F.K.; Abukhalil, M.H.; Ahmeda, A.F.; Mahmoud, A.M. Punicalagin prevents cisplatin-induced nephrotoxicity by attenuating oxidative stress, inflammatory response, and apoptosis in rats. Life Sci. 2021, 286, 120071. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Cao, K.; Liu, X.; Zhao, L.; Feng, Z.; Liu, J. Punicalagin Regulates Signaling Pathways in Inflammation-Associated Chronic Diseases. Antioxidants 2021, 11, 29. [Google Scholar] [CrossRef]
- El-Missiry, M.A.; Amer, M.A.; Hemieda, F.A.; Othman, A.I.; Sakr, D.A.; Abdulhadi, H.L. Cardioameliorative effect of punicalagin against streptozotocin-induced apoptosis, redox imbalance, metabolic changes and inflammation. Egypt. J. Basic Appl. Sci. 2015, 2, 247–260. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.C.; Hsu, Y.F.; Lin, T.C.; Hsu, H.Y. Antioxidant and hepatoprotective effects of punicalagin and punicalin on acetaminophen-induced liver damage in rats. Phytother. Res. 2001, 15, 206–212. [Google Scholar] [CrossRef]
- Zhang, Y.; Tan, X.; Cao, Y.; An, X.; Chen, J.; Yang, L. Punicalagin Protects against Diabetic Liver Injury by Upregulating Mitophagy and Antioxidant Enzyme Activities. Nutrients 2022, 14, 2782. [Google Scholar] [CrossRef]
- Fouad, A.A.; Qutub, H.O.; Al-Melhim, W.N. Punicalagin alleviates hepatotoxicity in rats challenged with cyclophosphamide. Environ. Toxicol. Pharmacol. 2016, 45, 158–162. [Google Scholar] [CrossRef]
- Foroutanfar, A.; Mehri, S.; Kamyar, M.; Tandisehpanah, Z.; Hosseinzadeh, H. Protective effect of punicalagin, the main polyphenol compound of pomegranate, against acrylamide-induced neurotoxicity and hepatotoxicity in rats. Phytother. Res. 2020, 34, 3262–3272. [Google Scholar] [CrossRef]
- Luo, J.; Long, Y.; Ren, G.; Zhang, Y.; Chen, J.; Huang, R.; Yang, L. Punicalagin reversed the hepatic injury of Tetrachloromethane by Antioxidation and enhancement of autophagy. J. Med. Food 2019, 22, 1271–1279. [Google Scholar] [CrossRef]
- El-Missiry, M.A.; ElKomy, M.A.; Othman, A.I.; AbouEl-Ezz, A.M. Punicalagin ameliorates the elevation of plasma homocysteine, amyloid-β, TNF-α and apoptosis by advocating antioxidants and modulating apoptotic mediator proteins in brain. Biomed. Pharmacother. 2018, 102, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Abdulhadi, H.L.; Dabdoub, B.R.; Ali, L.H.; Othman, A.I.; Amer, M.E.; El-Missiry, M.A. Punicalagin protects against the development of pancreatic injury and insulitis in rats with induced T1DM by reducing inflammation and oxidative stress. Mol. Cell. Biochem. 2022, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Venusova, E.; Kolesarova, A.; Horky, P.; Slama, P. Physiological and immune functions of punicalagin. Nutrients 2021, 13, 2150. [Google Scholar] [CrossRef] [PubMed]
- Ojo, O.O.; Ajayi, O.O.; Ogunbiyi, B.T. Down-regulation of BMP8A, SMADs 1/5/8 and BAX Proteins Following Methotrexate-treatment in Testicular Tissue of Swiss Albino Mice. Annu. Res. Rev. Biol. 2021, 1–9. [Google Scholar] [CrossRef]
- Karimi, M.Y.; Fatemi, I.; Kalantari, H.; Mombeini, M.A.; Mehrzadi, S.; Goudarzi, M. Ellagic acid prevents oxidative stress, inflammation, and histopathological alterations in acrylamide-induced hepatotoxicity in wistar rats. J. Diet. Suppl. 2020, 17, 651–662. [Google Scholar] [CrossRef]
- Zheng, D.; Liu, Z.; Zhou, Y.; Hou, N.; Yan, W.; Qin, Y.; Ye, Q.; Cheng, X.; Xiao, Q.; Bao, Y. Urolithin B, a gut microbiota metabolite, protects against myocardial ischemia/reperfusion injury via p62/Keap1/Nrf2 signaling pathway. Pharmacol. Res. 2020, 153, 104655. [Google Scholar] [CrossRef]
- Tang, L.; Mo, Y.; Li, Y.; Zhong, Y.; He, S.; Zhang, Y.; Tang, Y.; Fu, S.; Wang, X.; Chen, A. Urolithin A alleviates myocardial ischemia/reperfusion injury via PI3K/Akt pathway. Biochem. Biophys. Res. Commun. 2017, 486, 774–780. [Google Scholar] [CrossRef]
- Cerdá, B.; Llorach, R.; Cerón, J.J.; Espín, J.C.; Tomás-Barberán, F.A. Evaluation of the bioavailability and metabolism in the rat of punicalagin, an antioxidant polyphenol from pomegranate juice. Eur. J. Nutr. 2003, 42, 18–28. [Google Scholar] [CrossRef]
- Alfei, S.; Marengo, B.; Zuccari, G. Oxidative stress, antioxidant capabilities, and bioavailability: Ellagic acid or urolithins? Antioxidants 2020, 9, 707. [Google Scholar] [CrossRef]
- Wang, G.; Peng, X. A review of clinical applications and side effects of methotrexate in ophthalmology. J. Ophthalmol. 2020, 2020, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, M.M.; Bakr, A.G.; Osman, A.T. Protective effects of phloridzin against methotrexate-induced liver toxicity in rats. Biomed. Pharmacother. 2017, 95, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.M.; Hozayen, W.G.; Ramadan, S.M. Berberine ameliorates methotrexate-induced liver injury by activating Nrf2/HO-1 pathway and PPARγ, and suppressing oxidative stress and apoptosis in rats. Biomed. Pharmacother. 2017, 94, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.M.; Hussein, O.E.; Hozayen, W.G.; Bin-Jumah, M.; El-Twab, A.; Sanaa, M. Ferulic acid prevents oxidative stress, inflammation, and liver injury via upregulation of Nrf2/HO-1 signaling in methotrexate-induced rats. Environ. Sci. Pollut. Res. 2020, 27, 7910–7921. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, M.E.; Brosnan, J.T. Hepatic glutamate metabolism: A tale of 2 hepatocytes. Am. J. Clin. Nutr. 2009, 90, 857S–861S. [Google Scholar] [CrossRef] [Green Version]
- Kalra, A.; Yetiskul, E.; Wehrle, C.J.; Tuma, F. Physiology, Liver. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Manthorpe, E.M.; Jerrett, I.V.; Rawlin, G.T.; Woolford, L. Plant and fungal hepatotoxicities of cattle in australia, with a focus on minimally understood toxins. Toxins 2020, 12, 707. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Shikano, K.; Nanki, T.; Kawai, S. Folylpolyglutamate synthase is a major determinant of intracellular methotrexate polyglutamates in patients with rheumatoid arthritis. Sci. Rep. 2016, 6, 35615. [Google Scholar] [CrossRef]
- West, S.G. Methotrexate hepatotoxicity. Rheum. Dis. Clin. N. Am. 1997, 23, 883–915. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxidative Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Z.; Yan, L.-J. Protein oxidative modifications: Beneficial roles in disease and health. J. Biochem. Pharmacol. Res. 2013, 1, 15–26. [Google Scholar] [PubMed]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [PubMed]
- Joardar, S.; Dewanjee, S.; Bhowmick, S.; Dua, T.K.; Das, S.; Saha, A.; De Feo, V. Rosmarinic acid attenuates cadmium-induced nephrotoxicity via inhibition of oxidative stress, apoptosis, inflammation and fibrosis. Int. J. Mol. Sci. 2019, 20, 2027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Tovar, E.; Muriel, P. Molecular mechanisms that link oxidative stress, inflammation, and fibrosis in the liver. Antioxidants 2020, 9, 1279. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Wilkinson, F.L.; Sandhu, M.A.; Lightfoot, A.P. The interplay of oxidative stress and inflammation: Mechanistic insights and therapeutic potential of antioxidants. Oxidative Med. Cell. Longev. 2021, 2021, 9851914. [Google Scholar] [CrossRef]
- Yan, C.; Sun, W.; Wang, X.; Long, J.; Liu, X.; Feng, Z.; Liu, J. Punicalagin attenuates palmitate-induced lipotoxicity in HepG2 cells by activating the Keap1-Nrf2 antioxidant defense system. Mol. Nutr. Food Res. 2016, 60, 1139–1149. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.M.; Al Dera, H.S. 18β-Glycyrrhetinic acid exerts protective effects against cyclophosphamide-induced hepatotoxicity: Potential role of PPARγ and Nrf2 upregulation. Genes Nutr. 2015, 10, 41. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.M.; Germoush, M.O.; Alotaibi, M.F.; Hussein, O.E. Possible involvement of Nrf2 and PPARγ up-regulation in the protective effect of umbelliferone against cyclophosphamide-induced hepatotoxicity. Biomed. Pharmacother. 2017, 86, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Chen, S.; Xiong, H.; Wang, M.; Hang, W.; Zhu, X.; Zheng, Y.; Ge, B.; Li, R.; Cui, H. Astaxanthin from Haematococcus pluvialis ameliorates the chemotherapeutic drug (doxorubicin) induced liver injury through the Keap1/Nrf2/HO-1 pathway in mice. Food Funct. 2020, 11, 4659–4671. [Google Scholar] [CrossRef]
- Lv, H.; Xiao, Q.; Zhou, J.; Feng, H.; Liu, G.; Ci, X. Licochalcone a upregulates Nrf2 antioxidant pathway and thereby alleviates acetaminophen-induced hepatotoxicity. Front. Pharmacol. 2018, 9, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Li, H.; Hou, X.; Li, D.; He, S.; Wan, C.; Yin, P.; Liu, M.; Liu, F.; Xu, J. Punicalagin induces Nrf2/HO-1 expression via upregulation of PI3K/AKT pathway and inhibits LPS-induced oxidative stress in RAW264. 7 macrophages. Mediat. Inflamm. 2015, 2015, 380218. [Google Scholar] [CrossRef] [Green Version]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15N]nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Nishikimi, M.; Rao, N.A.; Yagi, K. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun. 1972, 46, 849–854. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Griffith, O.W. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-khawalde, A.A.-m.A.; Abukhalil, M.H.; Jghef, M.M.; Alfwuaires, M.A.; Alaryani, F.S.; Aladaileh, S.H.; Algefare, A.I.; Karimulla, S.; Alasmari, F.; Aldal’in, H.K.; et al. Punicalagin Protects against the Development of Methotrexate-Induced Hepatotoxicity in Mice via Activating Nrf2 Signaling and Decreasing Oxidative Stress, Inflammation, and Cell Death. Int. J. Mol. Sci. 2022, 23, 12334. https://doi.org/10.3390/ijms232012334
Al-khawalde AA-mA, Abukhalil MH, Jghef MM, Alfwuaires MA, Alaryani FS, Aladaileh SH, Algefare AI, Karimulla S, Alasmari F, Aldal’in HK, et al. Punicalagin Protects against the Development of Methotrexate-Induced Hepatotoxicity in Mice via Activating Nrf2 Signaling and Decreasing Oxidative Stress, Inflammation, and Cell Death. International Journal of Molecular Sciences. 2022; 23(20):12334. https://doi.org/10.3390/ijms232012334
Chicago/Turabian StyleAl-khawalde, Alayn’ Al-marddyah A., Mohammad H. Abukhalil, Muthana M. Jghef, Manal A. Alfwuaires, Fatima S. Alaryani, Saleem H. Aladaileh, Abdulmohsen I. Algefare, Shaik Karimulla, Fawaz Alasmari, Hammad Khalifeh Aldal’in, and et al. 2022. "Punicalagin Protects against the Development of Methotrexate-Induced Hepatotoxicity in Mice via Activating Nrf2 Signaling and Decreasing Oxidative Stress, Inflammation, and Cell Death" International Journal of Molecular Sciences 23, no. 20: 12334. https://doi.org/10.3390/ijms232012334