1. Introduction
Hepatocellular carcinoma (HCC) is one common malignant tumor of the digestive system, and is a major threat to public health, especially in China [
1]. It is a heterogeneous malignant tumor and has the sixth highest morbidity and fourth highest mortality in the world [
2]. Currently, surgical resection and liver transplantation may be the best ways to treat HCC, but the 5-year overall survival rate is only 7% [
3]. Although great advances have been made in the experimental and clinical treatment of HCC, its diagnosis is very difficult, and its prognosis is poor due to complicated etiology and multiple genes’ alteration in individuals [
4,
5,
6]. Therefore, a better understanding of the molecular mechanisms and highly reliable biomarkers of HCC is of great significance to promoting the diagnosis and development of therapeutic strategies for HCC.
RNA levels are regulated during cell differentiation and development in responses to intrinsic and extrinsic stimulation. Recently, RNA degradation has begun to be regarded as a significant mechanism of RNA regulation in the control of the gene expression involved in tumor progression [
7]. Ample information accumulated on RNA degradation has been studied intensely [
8]. It is noteworthy that the exosome complex (EXOSC) plays a central role in the degradation process of multiple
RNAs both in the nucleus and cytoplasm [
9]. In humans, the EXOSC is composed of ten evolutionarily conserved subunits, including the barrel-shaped core (EXOSC4-9 subunits) and three-subunit cap (EXOSC1-3) covering its surface, while EXOSC10 is bound to the catalytically inactive core and cap subunits of the EXOSC [
10]. Apart from the RNA degradation function of the EXOSC, its components are also associated with carcinogenesis. It has been reported that EXOSC1 could cleave single-stranded DNA and sensitize human kidney renal clear cell carcinoma cells to poly (ADP-ribose) polymerase inhibitors [
11]. Additionally, the enhancing expression of EXOSC2 directly regulated by tRNAGluUUC is closely related to breast cancer metastasis [
12], while EXOSC3 and EXOSC4 can trigger the development of colonic and colorectal cancer, respectively [
13,
14]. Similarly, EXOSC5, EXOSC8, EXOSC9 and EXOSC10 are also remarkably involved in the progression of various cancers, but there is no evidence to show the association between EXOSC6/7 and cancers at present. Interestingly, growing evidence suggests the role of EXOSC5 in the progression of cancers [
15,
16]. It has been reported that EXOSC5 is the no-catalytic component of EXOSC, while it can directly interact with the zinc-finger antiviral protein to degrade the target RNA. Currently, most studies have reported that up-regulation of EXOSC5 in various epithelial and hematopoietic tumor and cell lines [
15]. Recently, Pan et al. have indicated that the upregulation of EXOSC5 increases cell proliferation in colorectal cancer via the ERK and AKT signaling pathways [
15], suggesting the effects of EXOSC5 on digestive cancers. However, whether EXOSC5 is implicated in the process of HCC is still unclear. Thus, we aim to indicate the role and underlying mechanism of EXOSC5 in HCC via public databases and preliminarily validation from experiments in situ and in vitro.
A previous study demonstrated that the signal transducer and activator of transcription 3 (STAT3) signaling pathway played a key role in the proliferation of gastric cancer mediated by EXOSC5 [
10]. STAT3 is one of the members of the STAT family, which was first identified from a mouse liver cDNA library in the study of IL-6 signaling (NF-κB and STAT3) [
17,
18]. Several studies have found that STAT3 may be critical for the development of chemically induced HCC because it was a transcriptional factor closely related to immune responses, inflammation and tumorigenesis [
19]. However, the events that regulate STAT3 activation in human HCC are not known.
In this study, based on a variety of public databases and experimental analysis, for the first time we extended the research field of HCC with the purpose of determining the values of EXOSC family members (EXOSCs) in HCC. Moreover, the role of EXOSC5 in the cell growth of HCC and the underlying mechanism were also determined. The findings in this report shed light on the relationship between EXOSCs, especially EXOSC5, and HCC. It also provided a potential target for the diagnosis of HCC and indicated the possible mechanism of EXOSC5 in promoting HCC proliferation.
3. Discussion
HCC is a serious problem with an increased incidence in both developing and developed countries. Many factors, including virus infection and cirrhosis, have been proven to be closely related to HCC development [
20]. HCC is typically characterized by a low rate of early diagnosis and poor prognosis. Currently, elevated levels are regarded as a conventional biomarker for the diagnosis of HCC in clinical practice [
21], but it has its own limitations due to the fact that many advanced HCC patients are AFP-negative. Fortunately, systemic treatment options are available for advanced HCC in recent years with the advent of new therapeutic targets and immunotherapy, bringing new hope to HCC patients [
22]. Although advances have been made in the therapeutic management of HCC, the prognosis for it remains unknown and without established biomarkers. Therefore, HCC-related genes or biomarkers have yet to be identified to clarify the underlying mechanisms of its diagnosis, prognosis and treatment. In the present study, we comprehensively analyzed, for the first time, EXOSCs in terms of expression, mutation, prognostic value and immune cell infiltration in HCC. Even with evidence that EXOSCs have been confirmed to be associated with the development of many digestive cancers, distinct roles of EXOSCs in HCC remained to be investigated. Furthermore, the function of EXOSC5 and related mechanisms were verified by the cell assay and immunohistochemical staining.
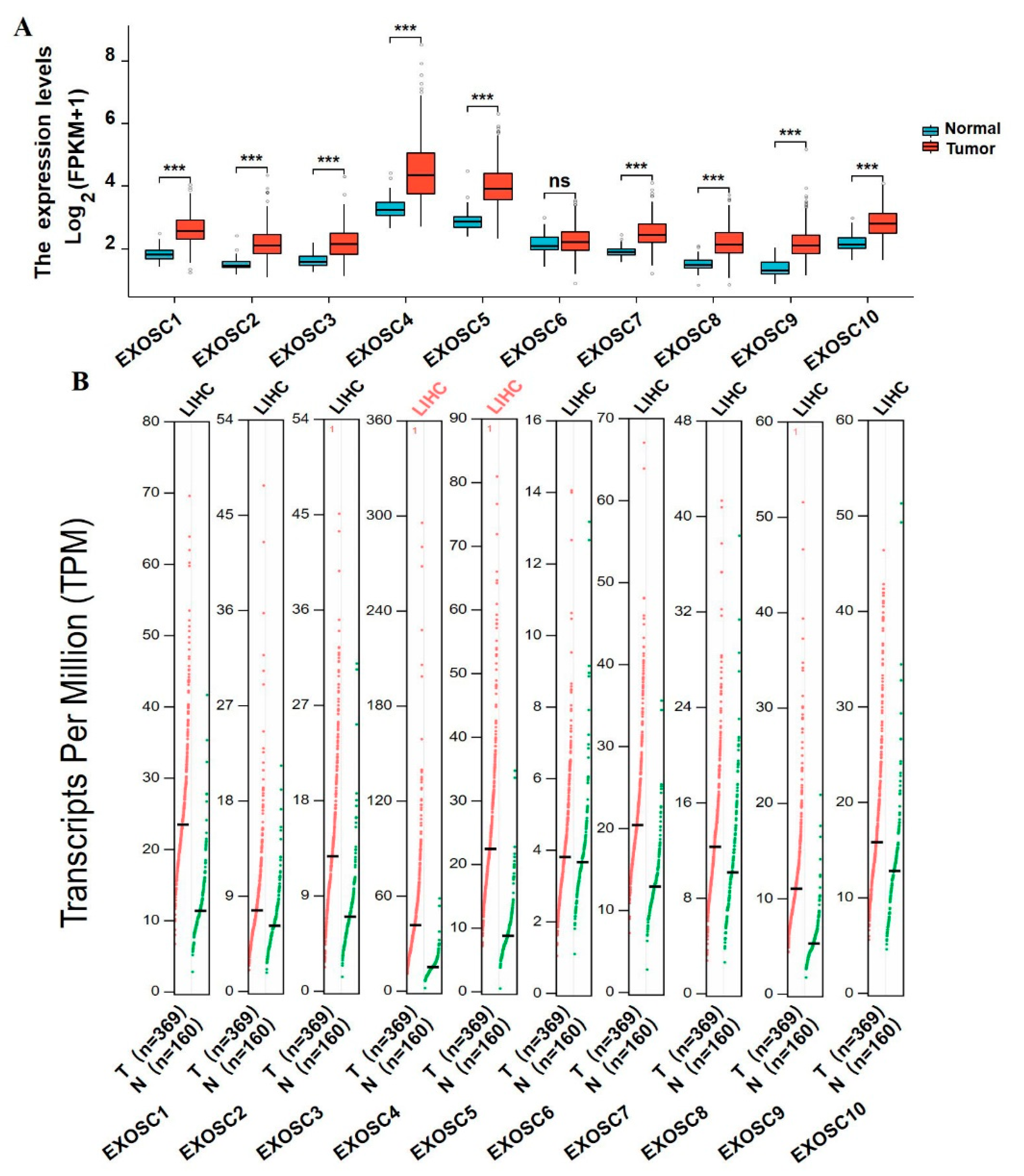
Being important components in RNA degradation, EXOSCs are crucial for controlling coding and non-coding RNA levels, profoundly participating in the elimination of defective RNAs in all organisms [
23]. In addition to DNA damage, results from the past couple of decades have unequivocally elucidated that RNA degradation takes part in the development and progression of multiple cancers [
24]. Therefore, EXOSCs that serve as an indispensable part of RNA stability may be associated with the process of cancers, making them an attractive target for cancer diagnosis and treatment. For instance, compared with healthy cells, cancer cells (i.e., MDA-MB-231, MCF-7 and Hela cells) showed higher activity of EXOSC9 [
25]. Additionally, functional experiments also confirmed the oncogenic roles of EXOSC8 in colorectal carcinoma [
26]. Results from our study showed that the mRNA expressions of EXOSCs, except EXOSC6, were differentially expressed in HCC, especially EXOSC4 and EXOSC5. Not only were the over-expression of EXOSCs proteins found, but a marked correlation between up-regulated EXOSC2/3/6/7/8/9/10 and clinical tumor stage was demonstrated. Moreover, we analyzed the expression of EXOSCs in multiple tumors relative to normal tissues. The results pointed out that EXOSCs were remarkably upregulated in various tumors, including cholangiocarcinoma (CHOL), kidney chromophobe (KICH) cancer, kidney renal clear cell carcinoma (KIRC), HCC, etc. (
Supplementary Figure S1). These data suggested the higher levels of EXOSCs in tumors, which were similar to those stated in previous reports.
Besides this, the correlation between EXOSCs and AFP, as well as the ROC, were used to analyze the ability of differentially expressed EXOSCs to predict HCC, suggesting that EXOSCs had a certain accuracy in predicting the outcome of HCC. Similarly, previous studies also identified the roles of EXOSC4 and EXOSC5 in predicting the prognosis of colorectal cancer and renal cell carcinoma, respectively [
14,
16]. At the same time, we also found that the high expression of EXOSCs was correlated with shorter OS and DFS, although there were some differences in the results from different databases analysis, indicating that differentially expressed EXOSCs influenced the prognosis of HCC patients. As a result, we speculated that EXOSCs may serve as potential diagnostic and prognostic markers for HCC. However, further verifications are required in order to discuss which EXOSC family members are directly related to poor prognosis of HCC.
Gene mutation is essential for biological evolution and tumorigenesis. There are currently few studies on the gene mutations of EXOSCs in the development and progression of cancer, while evidence has indicated that EXOSCs were closely related to many gene mutations [
11]. A previous review reported that the mutation of EXOSC2/3/8 could underly dysfunction and induced various diseases [
27]. Meanwhile, our genetic analysis indicated moderate genetic alterations of EXOSCs in HCC, which was consistent with previous reports to some extent. However, the association between EXOSCs mutations and HCC needs to be further explored. Array CGH and PPI mapping integrating study has proved that EXOSC1/3/8 were predicted to be the functional interaction factors in the network of the 6q23.3 locus in melanoma [
28]. Our data also uncovered the correlation between EXOSCs family members, and the similar results with previous study further prompted the significance of the interaction of EXOSCs family members. In this study, we also showed that EXOSCs participated in various biological processes, especially the metabolism of RNA, playing a key role in the development of multiple cancers. This not only confirmed the role of EXOSCs in cancer development [
29], but also provided clues for the future research on the function of EXOSCs in HCC.
The tumor microenvironment (TME) is a key factor in tumor progression and recurrence, causing it to become an increasingly popular topic. Growing evidence indicates that innate and adaptive immune cells could release inflammatory mediators such as cytokines, chemokines, growth factors and proteolytic enzymes, as well as activate transcription factors (NF-κB, STAT3, etc.), contributing to tumor progression when present in the TME [
30]. Therefore, immune cells are of significance in both clinical outcomes and the response to immunotherapy [
31,
32]. It has been reported that adaptive immune cells, such as CD8+ T cells, Th17 cells and B cells could stimulate HCC development, while adaptive immunity promotes immune surveillance to eradicate early HCC [
33]. According to these findings, we found that the expression of EXOSCs might be correlated with the infiltration of B cells, CD4+ T cells, CD8+ T cells, neutrophils, macrophages and dendritic cells, indicating that EXOSCs may reflect the immune status alongside disease prognosis. Thus, the study provides new insights into understanding the role of EXOSCs in the immune cell infiltration of HCC and provides detailed information to design new immunotherapies.
On the basis of these results from public databases, which suggested the significantly different expression and diagnostic, as well as prognostic, roles of EXOSC5, which are similar conclusions to those found in a previous report, exhibiting the potential clinical predictive and prognostic value of EXOSC5 [
15]. EXOSC5, also known as Rrp46p or CML28, is involved in a multitude of cellular RNA processing and degradation events via its exoribonuclease activity, showing an effect on HBV RNA degradation [
34,
35]. Thus, we decided to preliminarily validate the role of EXOSC5 in HCC proliferation. Yang et al. first identified EXOSC5, which is overexpressed in various tumor cell lines, but not in normal tissues [
36]. Wu et al. revealed that the elevation of EXOSC5 was observed in leukemic blasts from patients with acute myelogenous leukemia and chronic myelogenous leukemia blast crisis, while it is barely detected in normal bone marrow and peripheral blood [
37]. Additionally, the overexpression of EXOSC5 was also detected in various histological tumors [
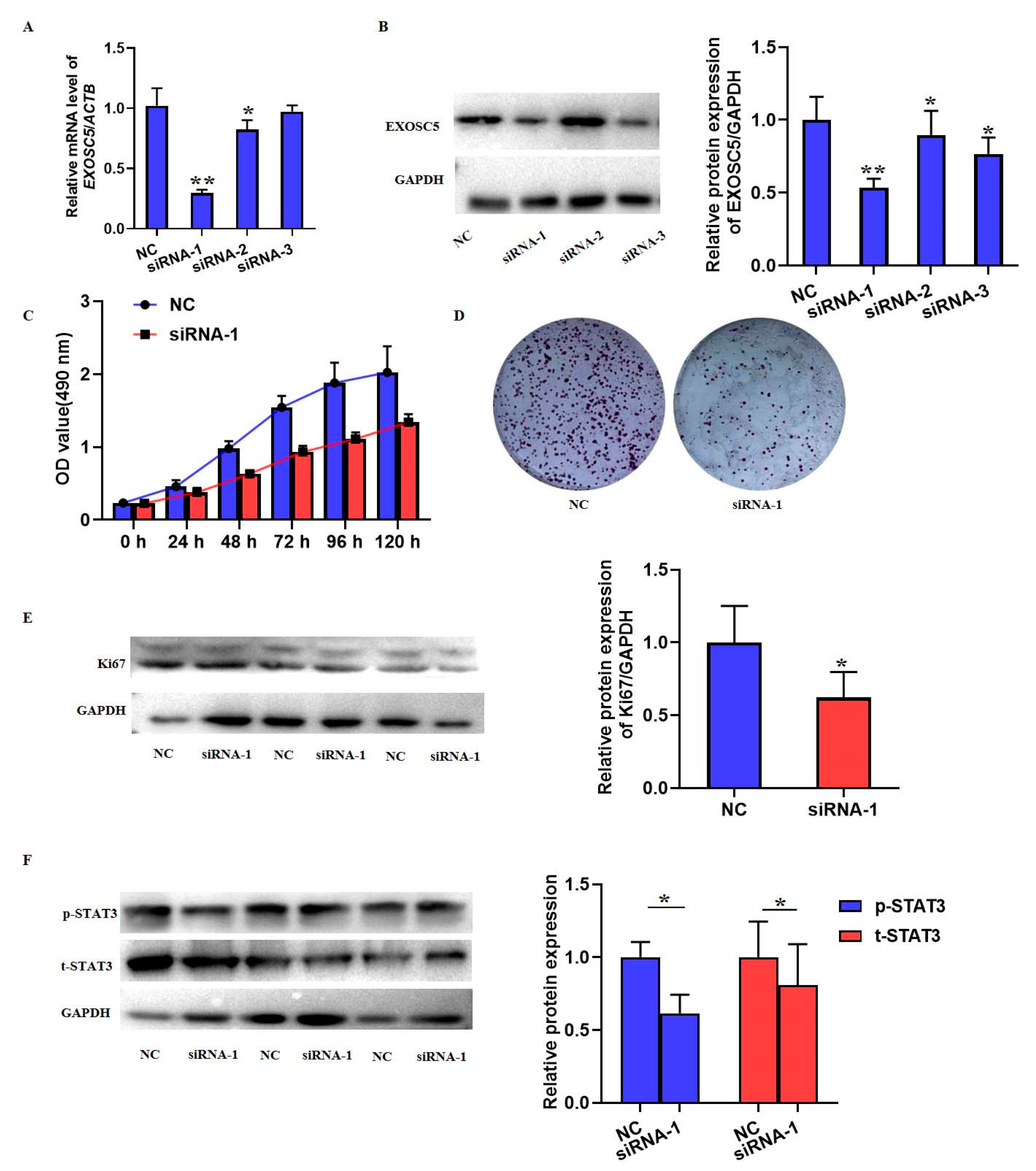
38]. Although results have highlighted the oncogenic role of EXOSC5 in colorectal cancer, there have been few studies on the role and related molecular mechanism of EXOSC5 in solid tumors. Herein, we found the upregulation of EXOSC5 mRNA and protein levels in HCC tumor tissues, and similar results were also found in HCC cell lines. Having a larger dataset about the expression of other members of EXOSCs family in HCC would offer further insights and more research is necessary in the future regarding EXOSCs. Evidence in our study also indicated that EXOSC5 knockdown suppressed the cell growth and proliferation of SMMC7721 cell lines. As far as we all know, this is the first study to report the role of EXOSC5 in HCC by in situ and in vitro experiments. A previous study reported that the STAT3 signaling pathway was involved in the EXOSC5-regulated proliferation of gastric cancer [
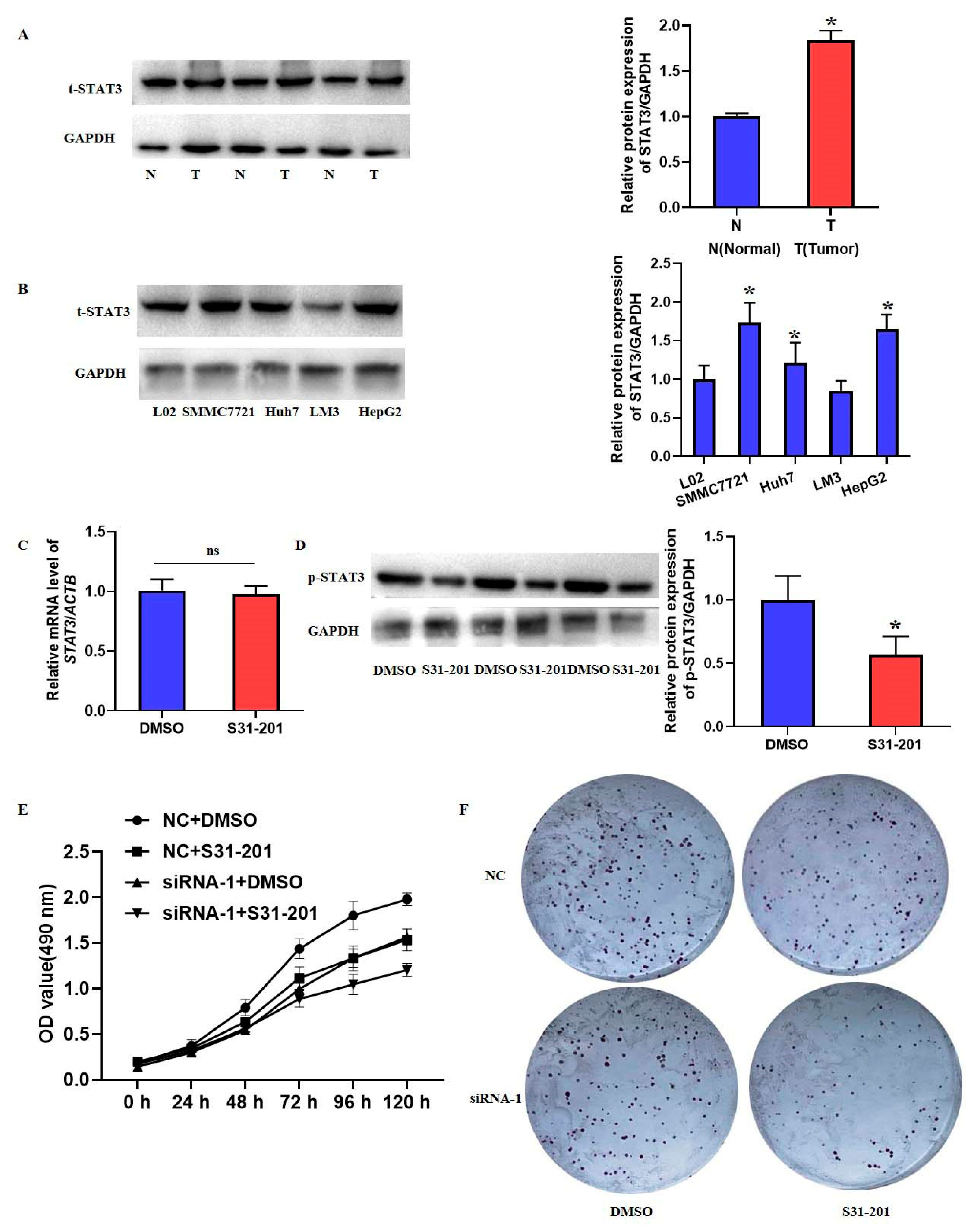
39], and the elevated phosphorylation of STAT3 was also shown in HCC tissues and cell lines, which could be inhibited by the down-regulation of EXOSC5 in the present study. Meanwhile, the increased phosphorylation of STAT3 was implicated in cell proliferation that is consistent with the function of STAT3 in cancers [
19]. Therefore, we suggested that EXOSC5 played important roles in HCC proliferation through activating STAT3 signaling. However, further studies are warranted to clarify the direct target and underlying mechanism of EXOSC5 in HCC patients.
Taken together, we systematically analyzed the differential expression and prognostic value of EXOSCs in HCC in silico for the first time. Furthermore, we deeply indicated that the over-expression of EXOSC5 was significantly related to proliferation of HCC via the STAT3 signaling pathway. Our study provided preliminary evidence for the role and a possible signaling for EXOSC5 in promoting HCC proliferation, but further research about molecular mechanisms of EXOSC5 in HCC is required.
Nevertheless, some limitations in our study should be recognized. Such as the location of these data only from public databases and lack of larger HCC cohorts. Moreover, other members of EXOSCs were also differentially expressed, except EXOSC5, but we did not provide the expression of these EXOSCs in the current study. In addition, the conclusion of our research only preliminarily confirmed the correlation between higher EXOSCs and HCC, as well as a related signaling of EXOSC5 promoting cell proliferation in vitro. More prospective data and larger experiments using tumor tissues, cells and animal models are required to confirm clinical suitability of EXOSCs in HCC.
4. Materials and Method
4.1. Data Resources for Clinical and Pathological Information of EXOSCs in HCC
In the study, we analyzed the expression of EXOSCs in HCC through multiple databases. TCGA (
https://www.cancer.gov/) (accessed on 22 July 2022) is a landmark cancer genomics program that provides molecular characterization of tumor samples and matched normal samples of more than 20,000 primary cancers across 33 cancer types. Clinical information on patients with HCC and high-throughput RNA-sequencing data were downloaded from the TCGA database. The transcript expression levels were estimated using the fragment per kilobase per million fragments mapped (FPKM) method in HTSeq. R (version 3.6.3) was used to analyze the expression levels of EXOSCs in tumor and adjacent tissues in the TCGA database and visualized with ggplot2 (3.3.3). Furthermore, other newly developed analytical tools, GEPIA2 (
http://gepia2.cancer-pku.cn)(accessed on 22 July 2022) [
40] and TIMER2 (
http://timer.cistrome.org/) (accessed on 22 July 2022) [
41], were applied to assess the mRNA expression of EXOSCs in HCC and normal tissues. GEPIA 2 is a newly developed tool using a standard processing pipeline to provide com-prehensive analysis, such as differential expression analysis, profiling plotting, correlation analysis and patient survival analysis, etc. TIMER2 is commonly used to analyze the relationship between immune infiltration and gene expression, clinical outcome and somatic mutations. Herein, we used it to investigate the differential expression of EXOSCs in multiple tumors. Besides, the total expression of EXOSCs in the Clinical Proteomic Tumor Analysis Consortium (CPTAC) database (
https://proteomics.cancer.gov/) (accessed on 22 July 2022) by The University of ALabama at Birmingham CANcer data analysis Portal (Ualcan) (
http://ualcan.path.uab.edu) (accessed on 22 July 2022) database, which could quantify and identify the constituent proteins of each tumor sample tumor biospecimen using mass spectrometry [
42,
43]. We analyzed the data of 379 HCC patients and 50 healthy people in TCGA and 110 healthy people in The Genotype-Tissue Expression (GTEx) (
www.gtexportal.org) (accessed on 22 July 2022). The Human Protein Atlas database (HPA) (
https://www.proteinatlas.org) (accessed on 22 July 2022) [
44,
45] is a Swedish initiative launched in 2003 to map all human proteins in cells, tissues and organs using the integration of various omics technologies, including antibody-based imaging, mass spectrometry-based proteomics and transcriptome science. The initiative also provides free access to immunohistochemical images of human-related tumor tissues and corresponding normal tissues. In this study, HPA was used to compare the protein levels of EXOSCs in HCC tumor and normal tissues.
4.2. Diagnostic Performance of EXOSCs
The serum AFP levels of HCC patients and normal were obtained by using the TCGA database, which were divided into two groups with a boundary of 400 ng/mL. R (version 3.6.3) was used to analyze the relationship between the expressions of EXOSCs and serum AFP. ggplot2 (3.3.3) was used for visual analysis. Additionally, we used GEPIA2.0 to draw ROC curve and calculated the AUC.
4.3. Survival Analysis for EXOSC Family Genes in HCC
The GEPIA2 website was used to analyze the survival data related to different cancer patients in the GTEx database (
www.gtexportal.org) (accessed on 22 July 2022). The influence of EXOSCs levels on the prognosis of HCC was analyzed. In addition, according to the median mRNA expression of EXOSCs, all HCC patients were divided into EXOSCs mRNA high and low expression groups from the TCGA database. Finally, the Kaplan–Meier survival curve was drawn by the survminer (version 4.9) package and the survival package (version 3.2-10) to analyze the effects of EXOSCs on the clinical prognosis of HCC patients. Furthermore, the results were verified again through the Kaplan–Meier plotter website (
www.kmplot.com/) (accessed on 22 July 2022). The website data comes from Gene Expression Omnibus (GEO), European Genome-phenome Archive (EGA) and TCGA. The Kaplan–Meier survival curve showed the relationship between EXOSCs and OS in HCC patients.
4.4. Genomic Mutation and Correlation Analysis of EXOSCs
cBioPortal (
http://cbioportal.org) (accessed on 22 July 2022) is an open-access resource that provides data from more than 5000 tumor samples from 20 cancer studies and explores the interaction of multidimensional cancer genomics data sets (The cBioportal: an open platform for exploring multidimensional cancer genomics data). This database was utilized to analyze alterations of EXOSCs in TCGA HCC samples, which further evaluated the correlation of these mutations with important clinicopathological factors.
4.5. Correlation and Interaction Analysis of EXOSCs
The TIMER (
http://timer.cistrome.org/) (accessed on 22 July 2022) database was applied to analyze the correlation of the expression of EXOSCs in HCC, and the statistical significance was evaluated by a Spearman’s test. In addition, STRING (
https://string-preview.org/) (accessed on 22 July 2022) provides users with protein interactions network functional enrichment analysis. Both of these were utilized to investigate the EXOSCs gene–gene and protein–protein interactions.
4.6. Functional Enrichment Analysis of EXOSCs
Metascape (
http://metascape.org) (accessed on 22 July 2022) is widely used in GO enrichment analysis of BP, CC, MF, as well as in pathway enrichment analysis (Metascape provides a biologist-oriented resource for the analysis of systems-level datasets). Herein, Metascape was used to perform pathway and process enrichment analysis of EXOSCs with the surrounding genes that were remarkably related to the expression of EXOSCs. A subset of enriched terms that were selected was displayed as a network plot, such that we could better understand the relationships between these terms. To further identify the closely related network components, the molecular complex detection (MCODE) algorithm was administered.
4.7. Immune Infiltration Analysis of EXOSCs
TIMER (
https://cistrome.shinyapps.io/timer/) (accessed on 22 July 2022) is a web-based application that could conduct a comprehensive analysis of the immune cell infiltration. In the present study, this database was used to analyze the correlation between the genes of EXOSCs and immune cells infiltration in HCC. Additionally, the TISIDB (
http://cis.hku.hk/TISIDB/) (accessed on 22 July 2022) database was administered to assess the expression of EXOSCs expression in different immune subtypes of HCC patients.
4.8. IHC
A total of 15 HCC and adjacent non-tumor tissues were collected from The First Affiliated Hospital of Hunan Normal University, Changsha, China. The tissues were all immediately frozen in liquid nitrogen and stored at −80 °C. The tissues were cut into 5 μm sections before 30 min at RT. The sections were then fixed with 4% PFA for 15 min at RT and washed with PBS. Then, 3% hydrogen peroxide (H2O2) was used to block the endogenous peroxidase activity for 30 min at RT, and the sections were pre-treated with sodium citrate buffer (pH = 6) for 30 min. The specific primary antibody of EXOSC5 (Proteintech group, Wuhan, China) was used to incubate the sections (1:200) at 4 °C overnight. 3,3-diaminobenzidine tetrahydrochloride (DAB) solution was used to stain the sections following with incubation by HRP-conjugated secondary antibody at 37 °C for 45 min. The sections were counterstained with hematoxylin and observed under a microscope.
4.9. Cell Culture
The normal hepatocytes (L02) were a gift from Nanjing Medical University and the HCC cell lines (HepG2, SMMC7721, Huh7, LM3) were purchased from the American Type Culture Collection (Manassas, VA, USA). L02 hepatocytes were cultured by RPMI1640 medium with 10% fetal bovine serum (FBS; Biological Industries, BeitHaemek, Israel) and 100 units/mL penicillin and streptomycin (Invitrogen, Carlsbad, CA, USA). HepG2, SMMC7721, Huh7 and LM3 cells were all cultured in Dulbecco’s modified Eagle’s medium (DMEM) with 10% FBS and 100 units/mL penicillin and streptomycin. The STAT3 inhibitor S31-201 dissolved in DMSO (Selleck Chemicals, Houston, TX, USA) was utilized to suppress STAT3 signaling. These cells were kept at 37 °C in a humidified % CO2 incubator.
4.10. Transfection of EXOSC5 siRNA in SMMC772 Cells
The SMMC7721 cells were seeded into 6-well cell culture tissue for 6~8 h. Then, 100 nm EXOSC5 siRNA and a negative control (NC) were transferred into SMMC7721 cells using Lipofectamine 2000 (Life Technologies, Carlsbad, CA, USA) according to the manufacturer’s protocol. The sequence of EXOSC5 siRNAs was synthesized by RIBIOBIO (Guangzhou, China). These sequences are as follow: siRNA-1, 5′-GCCAAAATCCGTG CTGAAAATGG-3′, siRNA-2,5′-CAGCAAAGAGATTTTCAACAAGG-3′, siRNA-3, 5′-GGAATGGATGAATCAATAAATT-3′. The knockdown expression of EXOSC5 was determined by qRT-PCR and western blot.
4.11. MTT Assay
SMMC7721 cells transfected with EXOSC5 siRNA or treated with S31-201were seeded into 96-well cell culture plates at density of 2 × 103. Then, 20 μL MTT (5 mg/mL) were added into the well of 0 h after 2 h. Subsequently, DMSO was added into each well every 24 h. After 120 h of culture, the absorbance was measured by microplate reader at a wavelength of 490 nm.
4.12. Colony Formation Assay
SMMC7721 cells transfected with EXOSC5 siRNA, or treated with S31-201, were seeded into 6-well culture plates at a density of 1 × 103 cells/well. The cells were cultured in an incubator with 5% CO2 at 37 °C for 2 weeks. The cells were then fixed with methanol for 20 min at room temperature followed by PBS washing. One percent crystal violet solution was utilized to stain the fixed cells. Photomicrographs of each well were obtained using a camera.
4.13. RNA Extraction and qRT-PCR Assay
The TRIzol regent (Invitrogen, Carlsbad, CA, USA) was used to extract the total RNA of tissues and cells according to the manufacturer’s protocol. Then primeScript RT Reagent Kit (Takara, Tokyo, Japan) was used to synthesize cDNA. The expression of EXOSC5 and STAT3 mRNA were determined by SYBR Green PCR Master Mix (Takara, Tokyo, Japan) on real-time PCR system (Roche, Indianapolis, IN, USA) at 95 °C for 30 s, followed by 40 cycles of 95 °C for 5 s and 60 °C for 34 s. ΔCT values were normalized by ACTB levels, and the relative expressions of mRNA to the control were calculated by 2−ΔΔCT. The sequence of related genes primers as follow: EXOSC5-F, 5′-ACTTTGCCTGCGAACAGAACC-3′, EXOSC5-R, 5′-CTCTTTGCTGACCTTCACCTC-3′; STAT3-F, 5′-GGCATTCGGAAAGTATTG-3′, STAT3-R, 5′-TCACCCACATTCACTCATT3′; ACTB-F, 5′-CACCAGGGCGTGATGGT-3′, ACTB-R, 5′-CTCAAACATGATCTGGGTCAT-3′.
4.14. Western Blot
Cells were seeded into 60 mm cell culture plates at a density of 4 × 105 for about 12~16 h. The cells were lysed using RIPA lysis buffer (Beyotime Institute of Biotechnology, Shanghai, China) containing protease inhibitor cocktail and phenylmethylsulfonyl fluoride (PMSF). The lyses was centrifuged at 1.5 × 104 cells/well for 15 min at 4 °C and the concentration of these supernatants were assayed by BCA Protein assay kit (Beyotime Institute of Biotechnology, Shanghai, China). To separate these proteins, 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was used, and they were then transferred into nitrocellulose (NC) membranes (PALL, Stevenage, UK). Then, the NC membranes were incubated with specific primary antibodies, including Ki67 (ABclonal Technology, Wuhan, China), STAT3 (ABclonal Technology, Wuhan, China), p-STAT3(Cell Signaling Technology, MA, USA), EXOSC5, GAPDH (Cell Signaling Technology, MA, USA) at 1:1000 overnight at 4 °C. In addition, horseradish peroxidase-conjugated immunoglobulin G (IgG) (Cell Signaling Technology, MA, USA) was used to incubate the membranes at 1:5000 for 1 h at room temperature. Immunoreactive protein bands were detected with an Enhanced Chemiluminescence (ECL) Detection Kit (Amersham, Piscataway, NJ, USA). Image J software was used to analyze the relative levels of proteins normalized to the expression of GAPDH.
4.15. Statistical Analysis
The data from experiments were presented as the mean ± SD from 3 independent experiments except for the MTT assay. Statistical comparisons between two groups were conducted using Student’s t-test, whereas an ANOVA was used to test the significance of differences between multiple groups. p < 0.05 was considered significant.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}










