
Role of STAR and SCP2/SCPx in the Transport of Cholesterol and Other Lipids


{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Synthesis
1.2. Distribution
1.3. Intracellular Trafficking: Vesicular and Non-Vesicular Cholesterol Transport
1.4. Cholesterol Transport in Steroid Biosynthesis
2. Model Cholesterol-Transport Proteins
2.1. STAR
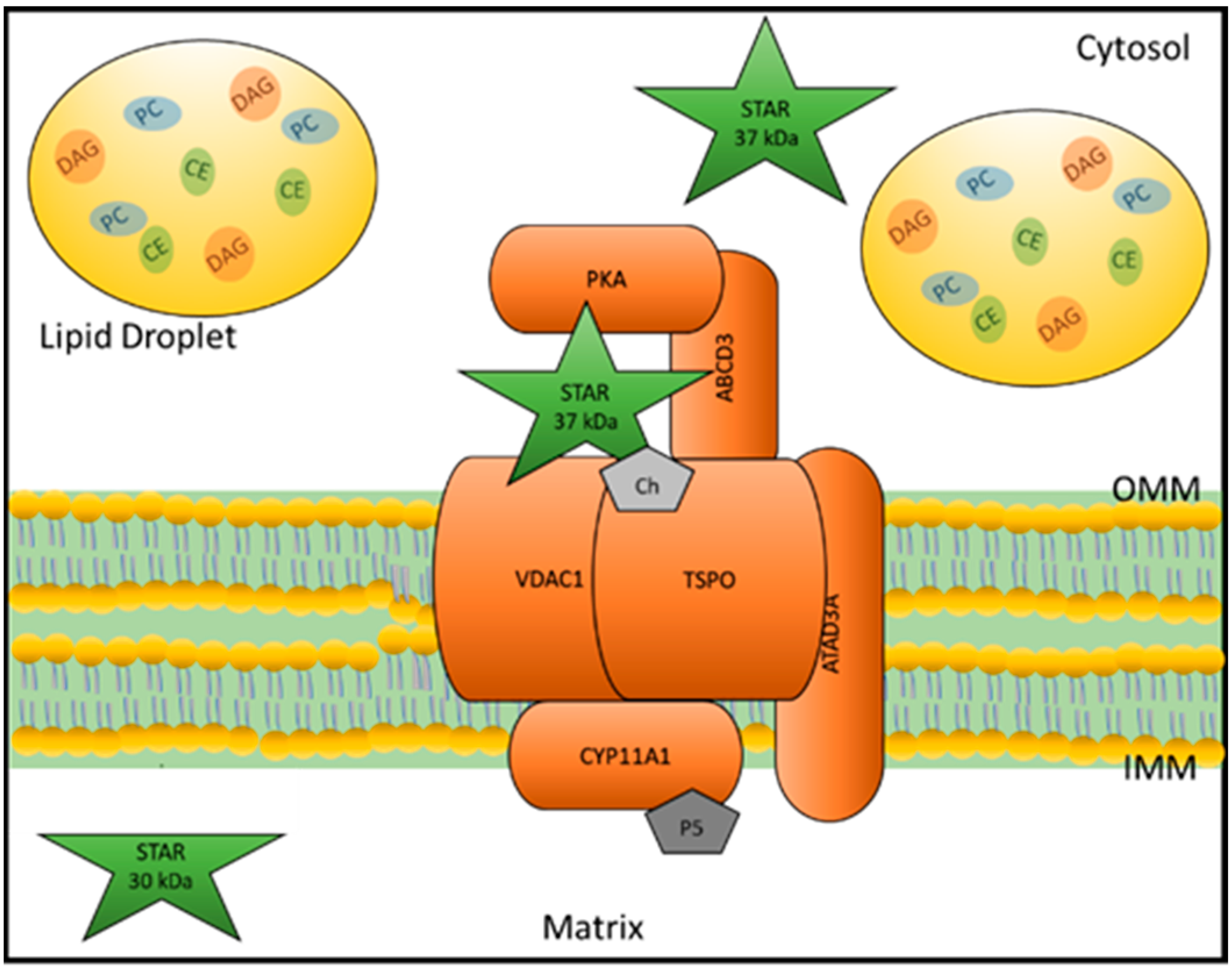
2.1.1. Role of STAR in Steroidogenesis
2.1.2. STAR Protein Activity
2.1.3. STAR Function in Cholesterol Transport and the START Domain
2.1.4. Other Roles of STAR
2.2. SCP2/SCPx
2.2.1. SCP2/SCPx Gene and Protein Products
2.2.2. Role of SCP2/SCPx in Cholesterol Transport
2.2.3. Role of SCP2 in Steroidogenesis
2.2.4. Role of SCP2/SCPx in the Transport of Other Lipids
2.2.5. Role of SCPx in Peroxisomal β-Oxidation
3. Future Directions and Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Luo, J.; Yang, H.; Song, B.L. Mechanisms and regulation of cholesterol homeostasis. Nat. Rev. Mol. Cell Biol. 2020, 21, 225–245. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, Y.; Chang, C.C.; Chang, T.Y. ACAT1/SOAT1 as a therapeutic target for Alzheimer’s disease. Future. Med. Chem. 2015, 7, 2451–2467. [Google Scholar] [CrossRef] [Green Version]
- Kuzu, O.F.; Noory, M.A.; Robertson, G.P. The Role of Cholesterol in Cancer. Cancer Res. 2016, 76, 2063–2070. [Google Scholar] [CrossRef] [Green Version]
- Cortes, V.A.; Busso, D.; Maiz, A.; Arteaga, A.; Nervi, F.; Rigotti, A. Physiological and pathological implications of cholesterol. Front. Biosci. 2014, 19, 416–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maxfield, F.R.; Tabas, I. Role of cholesterol and lipid organization in disease. Nature 2005, 438, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Luu, W.; Sharpe, L.J.; Capell-Hattam, I.; Gelissen, I.C.; Brown, A.J. Oxysterols: Old Tale, New Twists. Annu. Rev. Pharmacol. Toxicol. 2016, 56, 447–467. [Google Scholar] [CrossRef] [PubMed]
- Wong, L.H.; Gatta, A.T.; Levine, T.P. Lipid transfer proteins: The lipid commute via shuttles, bridges and tubes. Nat. Rev. Mol. Cell Biol. 2019, 20, 85–101. [Google Scholar] [CrossRef]
- Midzak, A.; Rone, M.; Aghazadeh, Y.; Culty, M.; Papadopoulos, V. Mitochondrial protein import and the genesis of steroidogenic mitochondria. Mol. Cell. Endocrinol. 2011, 336, 70–79. [Google Scholar] [CrossRef] [Green Version]
- Kriska, T.; Pilat, A.; Schmitt, J.C.; Girotti, A.W. Sterol carrier protein-2 (SCP-2) involvement in cholesterol hydroperoxide cytotoxicity as revealed by SCP-2 inhibitor effects. J. Lipid Res. 2010, 51, 3174–3184. [Google Scholar] [CrossRef] [Green Version]
- Galano, M.; Li, Y.; Li, L.; Sottas, C.; Papadopoulos, V. Role of Constitutive STAR in Leydig Cells. Int. J. Mol. Sci. 2021, 22, 2021. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, S.M.; Furth, E.E.; Ohba, T.; Chang, Y.J.; Rennert, H.; Sakuragi, N.; Billheimer, J.T.; Strauss, J.F. Sterol carrier protein 2: A role in steroid hormone synthesis? J. Steroid Biochem. Mol. Biol. 1993, 47, 167–172. [Google Scholar] [CrossRef]
- Petrescu, A.D.; Gallegos, A.M.; Okamura, Y.; Strauss, J.F.; Schroeder, F. Steroidogenic acute regulatory protein binds cholesterol and modulates mitochondrial membrane sterol domain dynamics. J. Biol. Chem. 2001, 276, 36970–36982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kallen, C.B.; Billheimer, J.T.; Summers, S.A.; Stayrook, S.E.; Lewis, M.; Strauss, J.F. Steroidogenic acute regulatory protein (StAR) is a sterol transfer protein. J. Biol. Chem. 1998, 273, 26285–26288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charreau, E.H.; Calvo, J.C.; Nozu, K.; Pignataro, O.; Catt, K.J.; Dufau, M.L. Hormonal modulation of 3-hydroxy-3-methylglutaryl coenzyme A reductase activity in gonadotropin-stimulated and -desensitized testicular Leydig cells. J. Biol. Chem. 1981, 256, 12719–12724. [Google Scholar] [CrossRef]
- Rainey, W.E.; Shay, J.W.; Mason, J.I. ACTH induction of 3-hydroxy-3-methylglutaryl coenzyme A reductase, cholesterol biosynthesis, and steroidogenesis in primary cultures of bovine adrenocortical cells. J. Biol. Chem. 1986, 261, 7322–7326. [Google Scholar] [CrossRef]
- Goldstein, J.L.; Brown, M.S. Regulation of the mevalonate pathway. Nature 1990, 343, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Bloch, K. Biological synthesis of cholesterol. Harvey Lect. 1952, 48, 68–88. [Google Scholar] [CrossRef] [PubMed]
- Cartwright, I.J.; Plonné, D.; Higgins, J.A. Intracellular events in the assembly of chylomicrons in rabbit enterocytes. J. Lipid Res. 2000, 41, 1728–1739. [Google Scholar] [CrossRef]
- Chang, T.Y.; Chang, C.C.; Ohgami, N.; Yamauchi, Y. Cholesterol sensing, trafficking, and esterification. Annu. Rev. Cell. Dev. Biol. 2006, 22, 129–157. [Google Scholar] [CrossRef]
- Hu, J.; Zhang, Z.; Shen, W.J.; Azhar, S. Cellular cholesterol delivery, intracellular processing and utilization for biosynthesis of steroid hormones. Nutr. Metab. 2010, 7, 47. [Google Scholar] [CrossRef]
- Rigotti, A.; Miettinen, H.E.; Krieger, M. The role of the high-density lipoprotein receptor SR-BI in the lipid metabolism of endocrine and other tissues. Endocr. Rev. 2003, 24, 357–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leiva, A.; Verdejo, H.; Benítez, M.L.; Martínez, A.; Busso, D.; Rigotti, A. Mechanisms regulating hepatic SR-BI expression and their impact on HDL metabolism. Atherosclerosis 2011, 217, 299–307. [Google Scholar] [CrossRef]
- Lange, Y.; Steck, T.L. Active membrane cholesterol as a physiological effector. Chem. Phys. Lipids 2016, 199, 74–93. [Google Scholar] [CrossRef]
- Steck, T.L.; Lange, Y. Cell cholesterol homeostasis: Mediation by active cholesterol. Trends Cell Biol. 2010, 20, 680–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epand, R.M. Proteins and cholesterol-rich domains. Biochim. Biophys. Acta 2008, 1778, 1576–1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liscum, L.; Munn, N.J. Intracellular cholesterol transport. Biochim. Biophys. Acta 1999, 1438, 19–37. [Google Scholar] [CrossRef]
- Soccio, R.E.; Breslow, J.L. Intracellular cholesterol transport. Arter. Thromb. Vasc. Biol. 2004, 24, 1150–1160. [Google Scholar] [CrossRef]
- Mukherjee, S.; Zha, X.; Tabas, I.; Maxfield, F.R. Cholesterol distribution in living cells: Fluorescence imaging using dehydroergosterol as a fluorescent cholesterol analog. Biophys. J. 1998, 75, 1915–1925. [Google Scholar] [CrossRef] [Green Version]
- Lange, Y. Disposition of intracellular cholesterol in human fibroblasts. J. Lipid Res. 1991, 32, 329–339. [Google Scholar] [CrossRef]
- Rone, M.B.; Fan, J.; Papadopoulos, V. Cholesterol transport in steroid biosynthesis: Role of protein-protein interactions and implications in disease states. Biochim. Biophys. Acta 2009, 1791, 646–658. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, V.; Miller, W.L. Role of mitochondria in steroidogenesis. Best Pract. Res. Clin. Endocrinol. Metab. 2012, 26, 771–790. [Google Scholar] [CrossRef] [PubMed]
- Lev, S. Non-vesicular lipid transport by lipid-transfer proteins and beyond. Nat. Rev. Mol. Cell Biol. 2010, 11, 739–750. [Google Scholar] [CrossRef]
- Baumann, N.A.; Sullivan, D.P.; Ohvo-Rekilä, H.; Simonot, C.; Pottekat, A.; Klaassen, Z.; Beh, C.T.; Menon, A.K. Transport of newly synthesized sterol to the sterol-enriched plasma membrane occurs via nonvesicular equilibration. Biochemistry 2005, 44, 5816–5826. [Google Scholar] [CrossRef] [PubMed]
- Urbani, L.; Simoni, R.D. Cholesterol and vesicular stomatitis virus G protein take separate routes from the endoplasmic reticulum to the plasma membrane. J. Biol. Chem. 1990, 265, 1919–1923. [Google Scholar] [CrossRef]
- Kaplan, M.R.; Simoni, R.D. Transport of cholesterol from the endoplasmic reticulum to the plasma membrane. J. Cell Biol. 1985, 101, 446–453. [Google Scholar] [CrossRef] [Green Version]
- DeGrella, R.F.; Simoni, R.D. Intracellular transport of cholesterol to the plasma membrane. J. Biol. Chem. 1982, 257, 14256–14262. [Google Scholar] [CrossRef]
- Lev, S. Nonvesicular lipid transfer from the endoplasmic reticulum. Cold Spring Harb. Perspect. Biol. 2012, 4, a013300. [Google Scholar] [CrossRef] [PubMed]
- Issop, L.; Rone, M.B.; Papadopoulos, V. Organelle plasticity and interactions in cholesterol transport and steroid biosynthesis. Mol. Cell. Endocrinol. 2013, 371, 34–46. [Google Scholar] [CrossRef]
- Simpson, E.R.; Boyd, G.S. The cholesterol side-chain cleavage system of the adrenal cortex: A mixed-function oxidase. Biochem. Biophys. Res. Commun. 1966, 24, 10–17. [Google Scholar] [CrossRef]
- Simpson, E.R.; Boyd, G.S. The cholesterol side-chain cleavage system of bovine adrenal cortex. Eur. J. Biochem. 1967, 2, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Jefcoate, C. High-flux mitochondrial cholesterol trafficking, a specialized function of the adrenal cortex. J. Clin. Investig. 2002, 110, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Rone, M.B.; Papadopoulos, V. Protein-protein interactions mediate mitochondrial cholesterol transport and steroid biosynthesis. J. Biol. Chem. 2006, 281, 38879–38893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauet, T.; Yao, Z.X.; Bose, H.S.; Wall, C.T.; Han, Z.; Li, W.; Hales, D.B.; Miller, W.L.; Culty, M.; Papadopoulos, V. Peripheral-type benzodiazepine receptor-mediated action of steroidogenic acute regulatory protein on cholesterol entry into leydig cell mitochondria. Mol. Endocrinol. 2005, 19, 540–554. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Degenhardt, B.; Tobin, D.; Yao, Z.X.; Tasken, K.; Papadopoulos, V. Identification, localization, and function in steroidogenesis of PAP7: A peripheral-type benzodiazepine receptor- and PKA (RIalpha)-associated protein. Mol. Endocrinol. 2001, 15, 2211–2228. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulos, V.; Amri, H.; Li, H.; Boujrad, N.; Vidic, B.; Garnier, M. Targeted disruption of the peripheral-type benzodiazepine receptor gene inhibits steroidogenesis in the R2C Leydig tumor cell line. J. Biol. Chem. 1997, 272, 32129–32135. [Google Scholar] [CrossRef] [Green Version]
- Rone, M.B.; Midzak, A.S.; Issop, L.; Rammouz, G.; Jagannathan, S.; Fan, J.; Ye, X.; Blonder, J.; Veenstra, T.; Papadopoulos, V. Identification of a dynamic mitochondrial protein complex driving cholesterol import, trafficking, and metabolism to steroid hormones. Mol. Endocrinol. 2012, 26, 1868–1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, W.L. Steroidogenic acute regulatory protein (StAR), a novel mitochondrial cholesterol transporter. Biochim. Biophys. Acta 2007, 1771, 663–676. [Google Scholar] [CrossRef]
- Bose, M.; Whittal, R.M.; Miller, W.L.; Bose, H.S. Steroidogenic activity of StAR requires contact with mitochondrial VDAC1 and phosphate carrier protein. J. Biol. Chem. 2008, 283, 8837–8845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stocco, D.M.; Clark, B.J. Regulation of the acute production of steroids in steroidogenic cells. Endocr. Rev. 1996, 17, 221–244. [Google Scholar] [CrossRef] [Green Version]
- Epstein, L.F.; Orme-Johnson, N.R. Regulation of steroid hormone biosynthesis. Identification of precursors of a phosphoprotein targeted to the mitochondrion in stimulated rat adrenal cortex cells. J. Biol. Chem. 1991, 266, 19739–19745. [Google Scholar] [CrossRef]
- Stocco, D.M.; Sodeman, T.C. The 30-kDa mitochondrial proteins induced by hormone stimulation in MA-10 mouse Leydig tumor cells are processed from larger precursors. J. Biol. Chem. 1991, 266, 19731–19738. [Google Scholar] [CrossRef]
- Clark, B.J.; Wells, J.; King, S.R.; Stocco, D.M. The purification, cloning, and expression of a novel luteinizing hormone-induced mitochondrial protein in MA-10 mouse Leydig tumor cells. Characterization of the steroidogenic acute regulatory protein (StAR). J. Biol. Chem. 1994, 269, 28314–28322. [Google Scholar] [CrossRef]
- Lin, D.; Sugawara, T.; Strauss, J.F.; Clark, B.J.; Stocco, D.M.; Saenger, P.; Rogol, A.; Miller, W.L. Role of steroidogenic acute regulatory protein in adrenal and gonadal steroidogenesis. Science 1995, 267, 1828–1831. [Google Scholar] [CrossRef]
- Sugawara, T.; Holt, J.A.; Driscoll, D.; Strauss, J.F., 3rd; Lin, D.; Miller, W.L.; Patterson, D.; Clancy, K.P.; Hart, I.M.; Clark, B.J.; et al. Human steroidogenic acute regulatory protein: Functional activity in COS-1 cells, tissue-specific expression, and mapping of the structural gene to 8p11.2 and a pseudogene to chromosome 13. Proc. Natl. Acad. Sci. USA 1995, 92, 4778–4782. [Google Scholar] [CrossRef] [Green Version]
- Tee, M.K.; Lin, D.; Sugawara, T.; Holt, J.A.; Guiguen, Y.; Buckingham, B.; Strauss, J.F., 3rd; Miller, W.L. T-->A transversion 11 bp from a splice acceptor site in the human gene for steroidogenic acute regulatory protein causes congenital lipoid adrenal hyperplasia. Hum. Mol. Genet. 1995, 4, 2299–2305. [Google Scholar] [CrossRef] [PubMed]
- Stocco, D.M. Clinical disorders associated with abnormal cholesterol transport: Mutations in the steroidogenic acute regulatory protein. Mol. Cell. Endocrinol. 2002, 191, 19–25. [Google Scholar] [CrossRef]
- Caron, K.M.; Soo, S.C.; Wetsel, W.C.; Stocco, D.M.; Clark, B.J.; Parker, K.L. Targeted disruption of the mouse gene encoding steroidogenic acute regulatory protein provides insights into congenital lipoid adrenal hyperplasia. Proc. Natl. Acad. Sci. USA 1997, 94, 11540–11545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strauss, J.F.; Kishida, T.; Christenson, L.K.; Fujimoto, T.; Hiroi, H. START domain proteins and the intracellular trafficking of cholesterol in steroidogenic cells. Mol. Cell. Endocrinol. 2003, 202, 59–65. [Google Scholar] [CrossRef]
- Tsujishita, Y.; Hurley, J.H. Structure and lipid transport mechanism of a StAR-related domain. Nat. Struct Biol. 2000, 7, 408–414. [Google Scholar] [CrossRef] [Green Version]
- Aghazadeh, Y.; Rone, M.B.; Blonder, J.; Ye, X.; Veenstra, T.D.; Hales, D.B.; Culty, M.; Papadopoulos, V. Hormone-induced 14-3-3γ adaptor protein regulates steroidogenic acute regulatory protein activity and steroid biosynthesis in MA-10 Leydig cells. J. Biol. Chem. 2012, 287, 15380–15394. [Google Scholar] [CrossRef] [PubMed]
- Artemenko, I.P.; Zhao, D.; Hales, D.B.; Hales, K.H.; Jefcoate, C.R. Mitochondrial processing of newly synthesized steroidogenic acute regulatory protein (StAR), but not total StAR, mediates cholesterol transfer to cytochrome P450 side chain cleavage enzyme in adrenal cells. J. Biol. Chem. 2001, 276, 46583–46596. [Google Scholar] [CrossRef] [Green Version]
- Clark, B.J.; Soo, S.C.; Caron, K.M.; Ikeda, Y.; Parker, K.L.; Stocco, D.M. Hormonal and developmental regulation of the steroidogenic acute regulatory protein. Mol. Endocrinol. 1995, 9, 1346–1355. [Google Scholar] [CrossRef]
- Clark, B.J.; Hudson, E.A. StAR Protein Stability in Y1 and Kin-8 Mouse Adrenocortical Cells. Biology 2015, 4, 200–215. [Google Scholar] [CrossRef] [Green Version]
- Granot, Z.; Geiss-Friedlander, R.; Melamed-Book, N.; Eimerl, S.; Timberg, R.; Weiss, A.M.; Hales, K.H.; Hales, D.B.; Stocco, D.M.; Orly, J. Proteolysis of normal and mutated steroidogenic acute regulatory proteins in the mitochondria: The fate of unwanted proteins. Mol. Endocrinol. 2003, 17, 2461–2476. [Google Scholar] [CrossRef] [Green Version]
- Arakane, F.; Sugawara, T.; Nishino, H.; Liu, Z.; Holt, J.A.; Pain, D.; Stocco, D.M.; Miller, W.L.; Strauss, J.F. Steroidogenic acute regulatory protein (StAR) retains activity in the absence of its mitochondrial import sequence: Implications for the mechanism of StAR action. Proc. Natl. Acad. Sci. USA 1996, 93, 13731–13736. [Google Scholar] [CrossRef] [Green Version]
- Bose, H.S.; Lingappa, V.R.; Miller, W.L. Rapid regulation of steroidogenesis by mitochondrial protein import. Nature 2002, 417, 87–91. [Google Scholar] [CrossRef]
- Arakane, F.; Kallen, C.B.; Watari, H.; Stayrook, S.E.; Lewis, M.; Strauss, J.F. Steroidogenic acute regulatory protein (StAR) acts on the outside of mitochondria to stimulate steroidogenesis. Endocr. Res. 1998, 24, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Bose, H.S.; Lingappa, V.R.; Miller, W.L. The steroidogenic acute regulatory protein, StAR, works only at the outer mitochondrial membrane. Endocr. Res. 2002, 28, 295–308. [Google Scholar] [CrossRef]
- Arakane, F.; King, S.R.; Du, Y.; Kallen, C.B.; Walsh, L.P.; Watari, H.; Stocco, D.M.; Strauss, J.F. Phosphorylation of steroidogenic acute regulatory protein (StAR) modulates its steroidogenic activity. J. Biol. Chem. 1997, 272, 32656–32662. [Google Scholar] [CrossRef] [Green Version]
- Jo, Y.; King, S.R.; Khan, S.A.; Stocco, D.M. Involvement of protein kinase C and cyclic adenosine 3′,5′-monophosphate-dependent kinase in steroidogenic acute regulatory protein expression and steroid biosynthesis in Leydig cells. Biol. Reprod. 2005, 73, 244–255. [Google Scholar] [CrossRef]
- Dyson, M.T.; Jones, J.K.; Kowalewski, M.P.; Manna, P.R.; Alonso, M.; Gottesman, M.E.; Stocco, D.M. Mitochondrial A-kinase anchoring protein 121 binds type II protein kinase A and enhances steroidogenic acute regulatory protein-mediated steroidogenesis in MA-10 mouse leydig tumor cells. Biol. Reprod. 2008, 78, 267–277. [Google Scholar] [CrossRef] [Green Version]
- Baker, B.Y.; Epand, R.F.; Epand, R.M.; Miller, W.L. Cholesterol binding does not predict activity of the steroidogenic acute regulatory protein, StAR. J. Biol. Chem. 2007, 282, 10223–10232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romanowski, M.J.; Soccio, R.E.; Breslow, J.L.; Burley, S.K. Crystal structure of the Mus musculus cholesterol-regulated START protein 4 (StarD4) containing a StAR-related lipid transfer domain. Proc. Natl. Acad. Sci. USA 2002, 99, 6949–6954. [Google Scholar] [CrossRef] [Green Version]
- Mathieu, A.P.; Fleury, A.; Ducharme, L.; Lavigne, P.; LeHoux, J.G. Insights into steroidogenic acute regulatory protein (StAR)-dependent cholesterol transfer in mitochondria: Evidence from molecular modeling and structure-based thermodynamics supporting the existence of partially unfolded states of StAR. J. Mol. Endocrinol. 2002, 29, 327–345. [Google Scholar] [CrossRef] [Green Version]
- Clark, B.J. The mammalian START domain protein family in lipid transport in health and disease. J. Endocrinol. 2012, 212, 257–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, W.L. StAR search--what we know about how the steroidogenic acute regulatory protein mediates mitochondrial cholesterol import. Mol. Endocrinol. 2007, 21, 589–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bose, H.S.; Whittal, R.M.; Baldwin, M.A.; Miller, W.L. The active form of the steroidogenic acute regulatory protein, StAR, appears to be a molten globule. Proc. Natl. Acad. Sci. USA 1999, 96, 7250–7255. [Google Scholar] [CrossRef] [Green Version]
- Galano, M.; Papadopoulos, V. Role of Constitutive STAR in Mitochondrial Structure and Function in MA-10 Leydig Cells. Endocrinology 2022, 163, bqac091. [Google Scholar] [CrossRef] [PubMed]
- Gallegos, A.M.; Atshaves, B.P.; Storey, S.M.; McIntosh, A.L.; Petrescu, A.D.; Schroeder, F. Sterol carrier protein-2 expression alters plasma membrane lipid distribution and cholesterol dynamics. Biochemistry 2001, 40, 6493–6506. [Google Scholar] [CrossRef]
- Puglielli, L.; Rigotti, A.; Greco, A.V.; Santos, M.J.; Nervi, F. Sterol carrier protein-2 is involved in cholesterol transfer from the endoplasmic reticulum to the plasma membrane in human fibroblasts. J. Biol. Chem. 1995, 270, 18723–18726. [Google Scholar] [CrossRef]
- Li, N.C.; Fan, J.J.; Papadopoulos, V. Sterol Carrier Protein-2, a Nonspecific Lipid-Transfer Protein, in Intracellular Cholesterol Trafficking in Testicular Leydig Cells. PLoS ONE 2016, 11, e0149728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohba, T.; Holt, J.A.; Billheimer, J.T.; Strauss, J.F. Human sterol carrier protein x/sterol carrier protein 2 gene has two promoters. Biochemistry 1995, 34, 10660–10668. [Google Scholar] [CrossRef]
- Atshaves, B.P.; Petrescu, A.D.; Starodub, O.; Roths, J.B.; Kier, A.B.; Schroeder, F. Expression and intracellular processing of the 58 kDa sterol carrier protein-2/3-oxoacyl-CoA thiolase in transfected mouse L-cell fibroblasts. J. Lipid Res. 1999, 40, 610–622. [Google Scholar] [CrossRef]
- Moncecchi, D.; Murphy, E.J.; Prows, D.R.; Schroeder, F. Sterol carrier protein-2 expression in mouse L-cell fibroblasts alters cholesterol uptake. Biochim. Biophys. Acta 1996, 1302, 110–116. [Google Scholar] [CrossRef]
- Vahouny, G.V.; Chanderbhan, R.; Kharroubi, A.; Noland, B.J.; Pastuszyn, A.; Scallen, T.J. Sterol carrier and lipid transfer proteins. Adv. Lipid Res. 1987, 22, 83–113. [Google Scholar] [CrossRef]
- Wanders, R.J.; Denis, S.; Wouters, F.; Wirtz, K.W.; Seedorf, U. Sterol carrier protein X (SCPx) is a peroxisomal branched-chain beta-ketothiolase specifically reacting with 3-oxo-pristanoyl-CoA: A new, unique role for SCPx in branched-chain fatty acid metabolism in peroxisomes. Biochem. Biophys. Res. Commun. 1997, 236, 565–569. [Google Scholar] [CrossRef] [Green Version]
- Mendis-Handagama, S.M.; Watkins, P.A.; Gelber, S.J.; Scallen, T.J. Leydig cell peroxisomes and sterol carrier protein-2 in luteinizing hormone-deprived rats. Endocrinology 1992, 131, 2839–2845. [Google Scholar] [CrossRef]
- Keller, G.A.; Scallen, T.J.; Clarke, D.; Maher, P.A.; Krisans, S.K.; Singer, S.J. Subcellular localization of sterol carrier protein-2 in rat hepatocytes: Its primary localization to peroxisomes. J. Cell Biol. 1989, 108, 1353–1361. [Google Scholar] [CrossRef] [Green Version]
- Mori, T.; Tsukamoto, T.; Mori, H.; Tashiro, Y.; Fujiki, Y. Molecular cloning and deduced amino acid sequence of nonspecific lipid transfer protein (sterol carrier protein 2) of rat liver: A higher molecular mass (60 kDa) protein contains the primary sequence of nonspecific lipid transfer protein as its C-terminal part. Proc. Natl. Acad. Sci. USA 1991, 88, 4338–4342. [Google Scholar] [CrossRef] [Green Version]
- Westerman, J.; Wirtz, K.W. The primary structure of the nonspecific lipid transfer protein (sterol carrier protein 2) from bovine liver. Biochem. Biophys. Res. Commun. 1985, 127, 333–338. [Google Scholar] [CrossRef]
- Stolowich, N.; Frolov, A.; Petrescu, A.D.; Scott, A.I.; Billheimer, J.T.; Schroeder, F. Holo-sterol carrier protein-2. (13)C NMR investigation of cholesterol and fatty acid binding sites. J. Biol. Chem. 1999, 274, 35425–35433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colles, S.M.; Woodford, J.K.; Moncecchi, D.; Myers-Payne, S.C.; McLean, L.R.; Billheimer, J.T.; Schroeder, F. Cholesterol interaction with recombinant human sterol carrier protein-2. Lipids 1995, 30, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Frolov, A.; Woodford, J.K.; Murphy, E.J.; Billheimer, J.T.; Schroeder, F. Spontaneous and protein-mediated sterol transfer between intracellular membranes. J. Biol. Chem. 1996, 271, 16075–16083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallegos, A.M.; Atshaves, B.P.; Storey, S.M.; Starodub, O.; Petrescu, A.D.; Huang, H.; McIntosh, A.L.; Martin, G.G.; Chao, H.; Kier, A.B.; et al. Gene structure, intracellular localization, and functional roles of sterol carrier protein-2. Prog. Lipid Res. 2001, 40, 498–563. [Google Scholar] [CrossRef]
- Atshaves, B.P.; Starodub, O.; McIntosh, A.; Petrescu, A.; Roths, J.B.; Kier, A.B.; Schroeder, F. Sterol carrier protein-2 alters high density lipoprotein-mediated cholesterol efflux. J. Biol. Chem. 2000, 275, 36852–36861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, E.J.; Schroeder, F. Sterol carrier protein-2 mediated cholesterol esterification in transfected L-cell fibroblasts. Biochim. Biophys. Acta 1997, 1345, 283–292. [Google Scholar] [CrossRef]
- Baum, C.L.; Reschly, E.J.; Gayen, A.K.; Groh, M.E.; Schadick, K. Sterol carrier protein-2 overexpression enhances sterol cycling and inhibits cholesterol ester synthesis and high density lipoprotein cholesterol secretion. J. Biol. Chem. 1997, 272, 6490–6498. [Google Scholar] [CrossRef] [Green Version]
- Zanlungo, S.; Amigo, L.; Mendoza, H.; Miquel, J.F.; Vío, C.; Glick, J.M.; Rodríguez, A.; Kozarsky, K.; Quiñones, V.; Rigotti, A.; et al. Sterol carrier protein 2 gene transfer changes lipid metabolism and enterohepatic sterol circulation in mice. Gastroenterology 2000, 119, 1708–1719. [Google Scholar] [CrossRef] [Green Version]
- Seedorf, U.; Raabe, M.; Ellinghaus, P.; Kannenberg, F.; Fobker, M.; Engel, T.; Denis, S.; Wouters, F.; Wirtz, K.W.A.; Wanders, R.J.A.; et al. Defective peroxisomal catabolism of branched fatty acyl coenzyme A in mice lacking the sterol carrier protein-2 sterol carrier protein-x gene function. Genes Dev. 1998, 12, 1189–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puglielli, L.; Rigotti, A.; Amigo, L.; Nuñez, L.; Greco, A.V.; Santos, M.J.; Nervi, F. Modulation of intrahepatic cholesterol trafficking: Evidence by in vivo antisense treatment for the involvement of sterol carrier protein-2 in newly synthesized cholesterol transport into rat bile. Biochem. J. 1996, 317 Pt 3, 681–687. [Google Scholar] [CrossRef]
- Chanderbhan, R.; Noland, B.J.; Scallen, T.J.; Vahouny, G.V. Sterol carrier protein2. Delivery of cholesterol from adrenal lipid droplets to mitochondria for pregnenolone synthesis. J. Biol. Chem. 1982, 257, 8928–8934. [Google Scholar] [CrossRef]
- Yamamoto, R.; Kallen, C.B.; Babalola, G.O.; Rennert, H.; Billheimer, J.T.; Strauss, J.F. Cloning and expression of a cDNA encoding human sterol carrier protein 2. Proc. Natl. Acad. Sci. USA 1991, 88, 463–467. [Google Scholar] [CrossRef] [Green Version]
- Rennert, H.; Amsterdam, A.; Billheimer, J.T.; Strauss, J.F. Regulated expression of sterol carrier protein 2 in the ovary: A key role for cyclic AMP. Biochemistry 1991, 30, 11280–11285. [Google Scholar] [CrossRef]
- Trzeciak, W.H.; Simpson, E.R.; Scallen, T.J.; Vahouny, G.V.; Waterman, M.R. Studies on the synthesis of sterol carrier protein-2 in rat adrenocortical cells in monolayer culture. Regulation by ACTH and dibutyryl cyclic 3′,5′-AMP. J. Biol. Chem. 1987, 262, 3713–3717. [Google Scholar] [CrossRef]
- McLean, M.P.; Puryear, T.K.; Khan, I.; Azhar, S.; Billheimer, J.T.; Orly, J.; Gibori, G. Estradiol regulation of sterol carrier protein-2 independent of cytochrome P450 side-chain cleavage expression in the rat corpus luteum. Endocrinology 1989, 125, 1337–1344. [Google Scholar] [CrossRef] [PubMed]
- van Noort, M.; Rommerts, F.F.; van Amerongen, A.; Wirtz, K.W. Localization and hormonal regulation of the non-specific lipid transfer protein (sterol carrier protein2) in the rat testis. J. Endocrinol. 1986, 109, R13–R16. [Google Scholar] [CrossRef]
- Gallegos, A.M.; Schoer, J.K.; Starodub, O.; Kier, A.B.; Billheimer, J.T.; Schroeder, F. A potential role for sterol carrier protein-2 in cholesterol transfer to mitochondria. Chem. Phys. Lipids 2000, 105, 9–29. [Google Scholar] [CrossRef]
- Schroeder, F.; Myerspayne, S.C.; Billheimer, J.T.; Wood, W.G. Probing the ligand-binding sites of fatty acid and sterol carrier proteins-effects of ethanol. Biochemistry 1995, 34, 11919–11927. [Google Scholar] [CrossRef]
- McArthur, M.J.; Atshaves, B.P.; Frolov, A.; Foxworth, W.D.; Kier, A.B.; Schroeder, F. Cellular uptake and intracellular trafficking of long chain fatty acids. J. Lipid Res. 1999, 40, 1371–1383. [Google Scholar] [CrossRef]
- Murphy, E.J. Sterol carrier protein-2 expression increases NBD-stearate uptake and cytoplasmic diffusion in L cells. Am. J. Physiol. 1998, 275, G237–G243. [Google Scholar] [CrossRef] [PubMed]
- Atshaves, B.P.; Storey, S.M.; Schroeder, F. Sterol carrier protein-2/sterol carrier protein-x expression differentially alters fatty acid metabolism in L cell fibroblasts. J. Lipid Res. 2003, 44, 1751–1762. [Google Scholar] [CrossRef] [Green Version]
- Murphy, E.J.; Stiles, T.; Schroeder, F. Sterol carrier protein-2 expression alters phospholipid content and fatty acyl composition in L-cell fibroblasts. J. Lipid Res. 2000, 41, 788–796. [Google Scholar] [CrossRef]
- Gossett, R.E.; Frolov, A.A.; Roths, J.B.; Behnke, W.D.; Kier, A.B.; Schroeder, F. Acyl-CoA binding proteins: Multiplicity and function. Lipids 1996, 31, 895–918. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, J.; Jensen, M.V.; Hansen, J.K.; Faergeman, N.J.; Neergaard, T.B.; Gaigg, B. Role of acylCoA binding protein in acylCoA transport, metabolism and cell signaling. Mol. Cell. Biochem. 1999, 192, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Starodub, O.; Jolly, C.A.; Atshaves, B.P.; Roths, J.B.; Murphy, E.J.; Kier, A.B.; Schroeder, F. Sterol carrier protein-2 localization in endoplasmic reticulum and role in phospholipid formation. Am. J. Physiol. Cell. Physiol. 2000, 279, C1259–C1269. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, F.; Zhou, M.; Swaggerty, C.L.; Atshaves, B.P.; Petrescu, A.D.; Storey, S.M.; Martin, G.G.; Huang, H.; Helmkamp, G.M.; Ball, J.M. Sterol carrier protein-2 functions in phosphatidylinositol transfer and signaling. Biochemistry 2003, 42, 3189–3202. [Google Scholar] [CrossRef] [PubMed]
- Atshaves, B.P.; Jefferson, J.R.; McIntosh, A.L.; Gallegos, A.; McCann, B.M.; Landrock, K.K.; Kier, A.B.; Schroeder, F. Effect of sterol carrier protein-2 expression on sphingolipid distribution in plasma membrane lipid rafts/caveolae. Lipids 2007, 42, 871–884. [Google Scholar] [CrossRef]
- Bloj, B.; Zilversmit, D.B. Accelerated transfer of neutral glycosphingolipids and ganglioside GM1 by a purified lipid transfer protein. J. Biol. Chem. 1981, 256, 5988–5991. [Google Scholar] [CrossRef]
- Fong, T.H.; Wang, S.M.; Lin, H.S. Immunocytochemical demonstration of a lipid droplet-specific capsule in cultured Leydig cells of the golden hamsters. J. Cell. Biochem. 1996, 63, 366–373. [Google Scholar] [CrossRef]
- Seedorf, U.; Brysch, P.; Engel, T.; Schrage, K.; Assmann, G. Sterol carrier protein X is peroxisomal 3-oxoacyl coenzyme A thiolase with intrinsic sterol carrier and lipid transfer activity. J. Biol. Chem. 1994, 269, 21277–21283. [Google Scholar] [CrossRef]
- Ferdinandusse, S.; Denis, S.; Faust, P.L.; Wanders, R.J. Bile acids: The role of peroxisomes. J. Lipid Res. 2009, 50, 2139–2147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atshaves, B.P.; McIntosh, A.L.; Landrock, D.; Payne, H.R.; Mackie, J.T.; Maeda, N.; Ball, J.; Schroeder, F.; Kier, A.B. Effect of SCP-x gene ablation on branched-chain fatty acid metabolism. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G939–G951. [Google Scholar] [CrossRef] [PubMed]
- Ferdinandusse, S.; Kostopoulos, P.; Denis, S.; Rusch, H.; Overmars, H.; Dillmann, U.; Reith, W.; Haas, D.; Wanders, R.J.A.; Duran, M.; et al. Mutations in the gene encoding peroxisomal sterol carrier protein X (SCPx) cause leukencephalopathy with dystonia and motor neuropathy. Am. J. Hum. Genet. 2006, 78, 1046–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, R.; Lewis-Smith, D.; Douroudis, K.; Duff, J.; Keogh, M.; Pyle, A.; Fletcher, N.; Chinnery, P.F. SCP2 Mutations and neurodegeneration with brain iron accumulation. Neurology 2015, 85, 1909–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galano, M.; Ezzat, S.; Papadopoulos, V. SCP2 variant is associated with alterations in lipid metabolism, brainstem neurodegeneration, and testicular defects. Hum. Genom. 2022, 16, 32. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galano, M.; Venugopal, S.; Papadopoulos, V. Role of STAR and SCP2/SCPx in the Transport of Cholesterol and Other Lipids. Int. J. Mol. Sci. 2022, 23, 12115. https://doi.org/10.3390/ijms232012115
Galano M, Venugopal S, Papadopoulos V. Role of STAR and SCP2/SCPx in the Transport of Cholesterol and Other Lipids. International Journal of Molecular Sciences. 2022; 23(20):12115. https://doi.org/10.3390/ijms232012115
Chicago/Turabian StyleGalano, Melanie, Sathvika Venugopal, and Vassilios Papadopoulos. 2022. "Role of STAR and SCP2/SCPx in the Transport of Cholesterol and Other Lipids" International Journal of Molecular Sciences 23, no. 20: 12115. https://doi.org/10.3390/ijms232012115