
Soluble CD146 as a Potential Target for Preventing Triple Negative Breast Cancer MDA-MB-231 Cell Growth and Dissemination

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Experimental Results
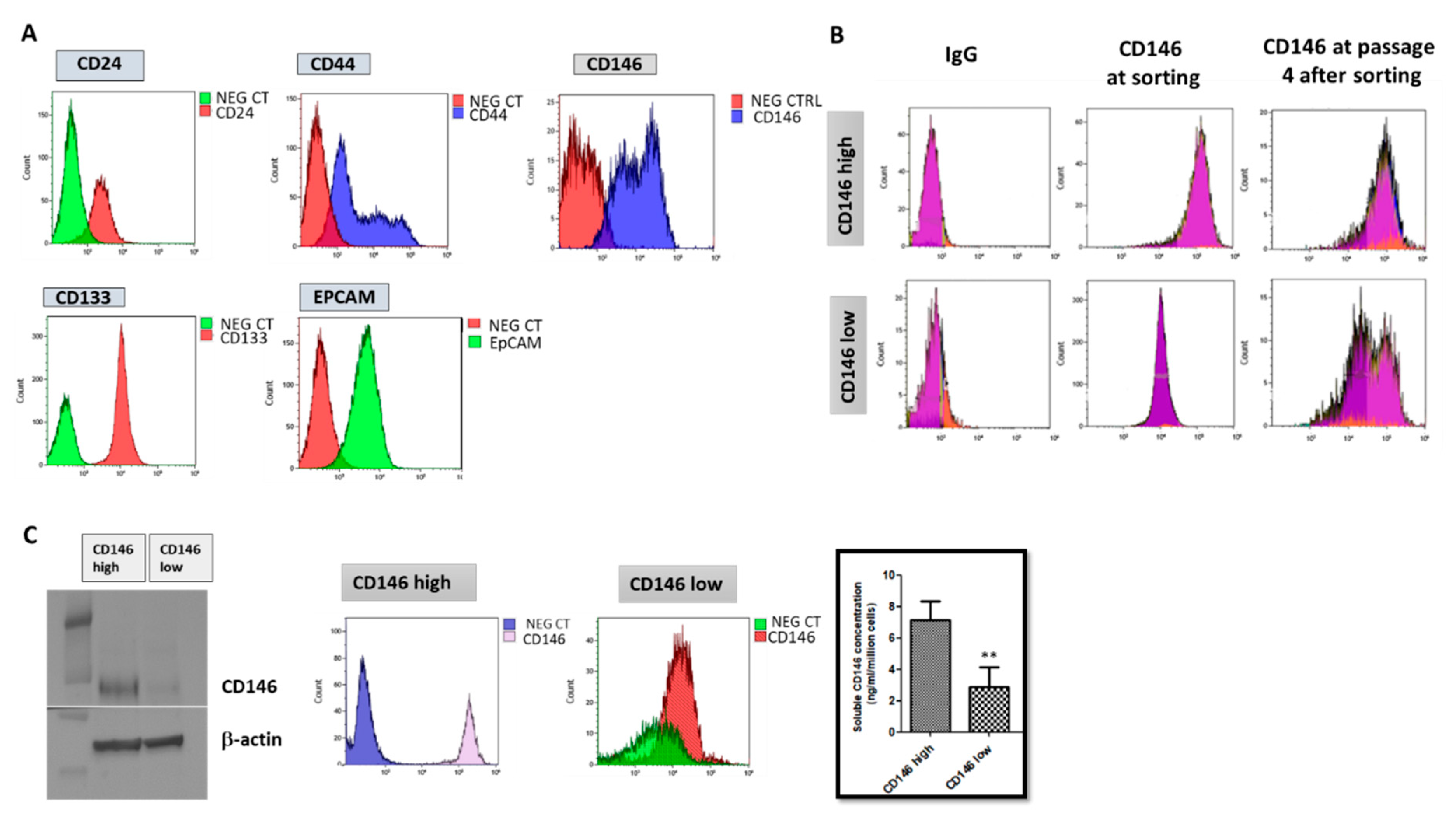
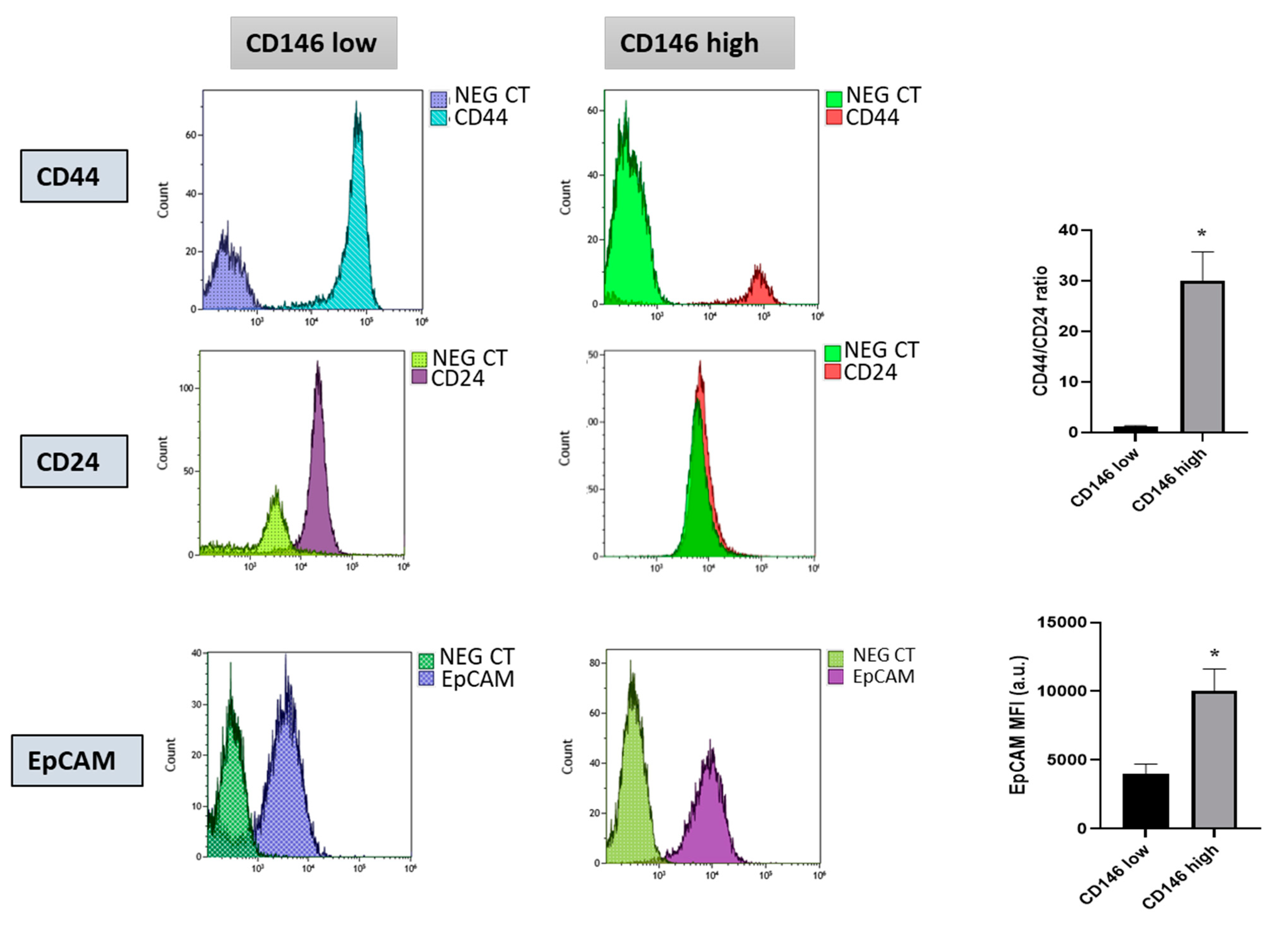
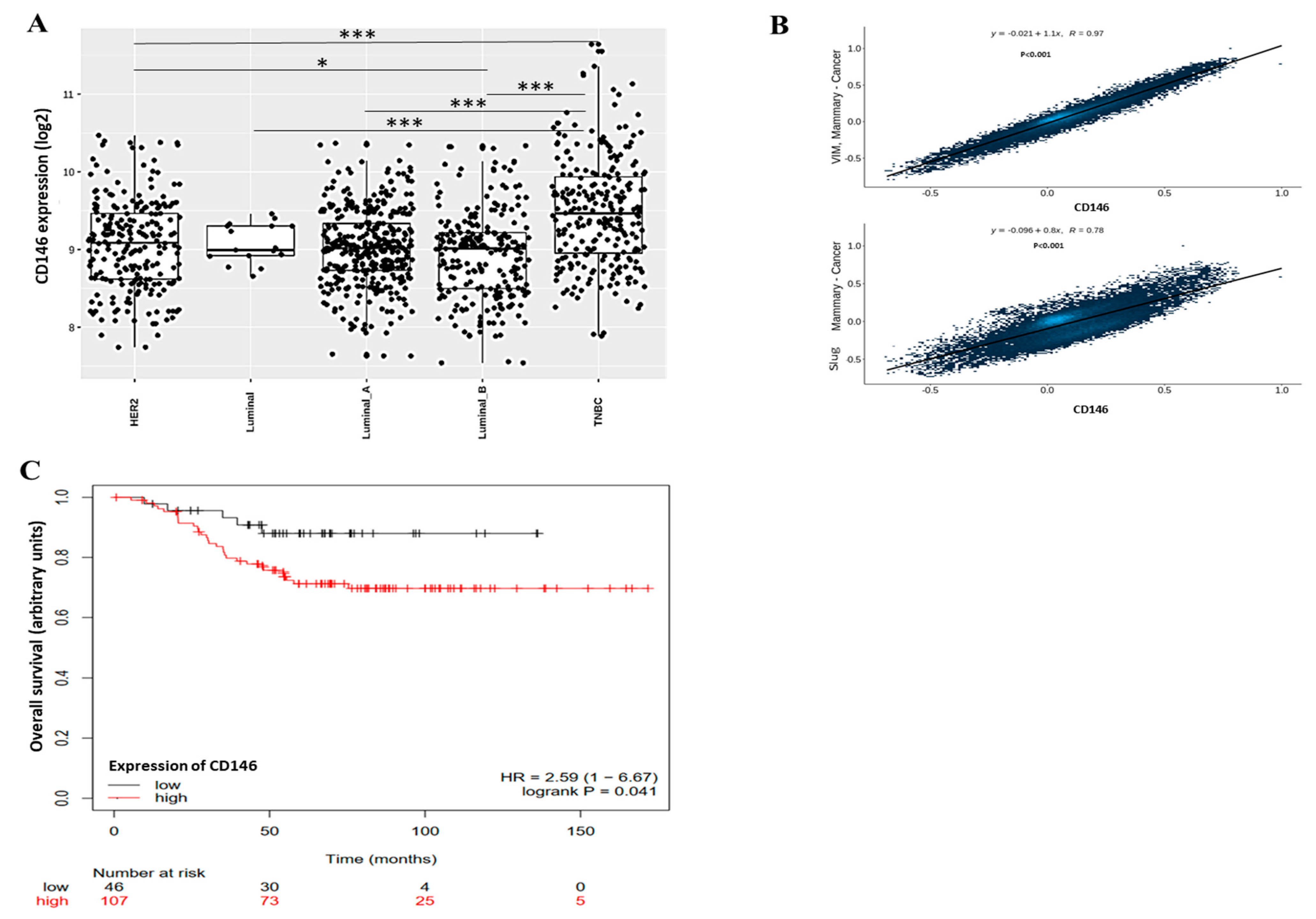
2.1.1. Triple Negative Breast Cancer Cells MDA-MB-231 Express Cancer Stem Cells Markers and Are Essentially Composed of Cells with High CD146 Expression
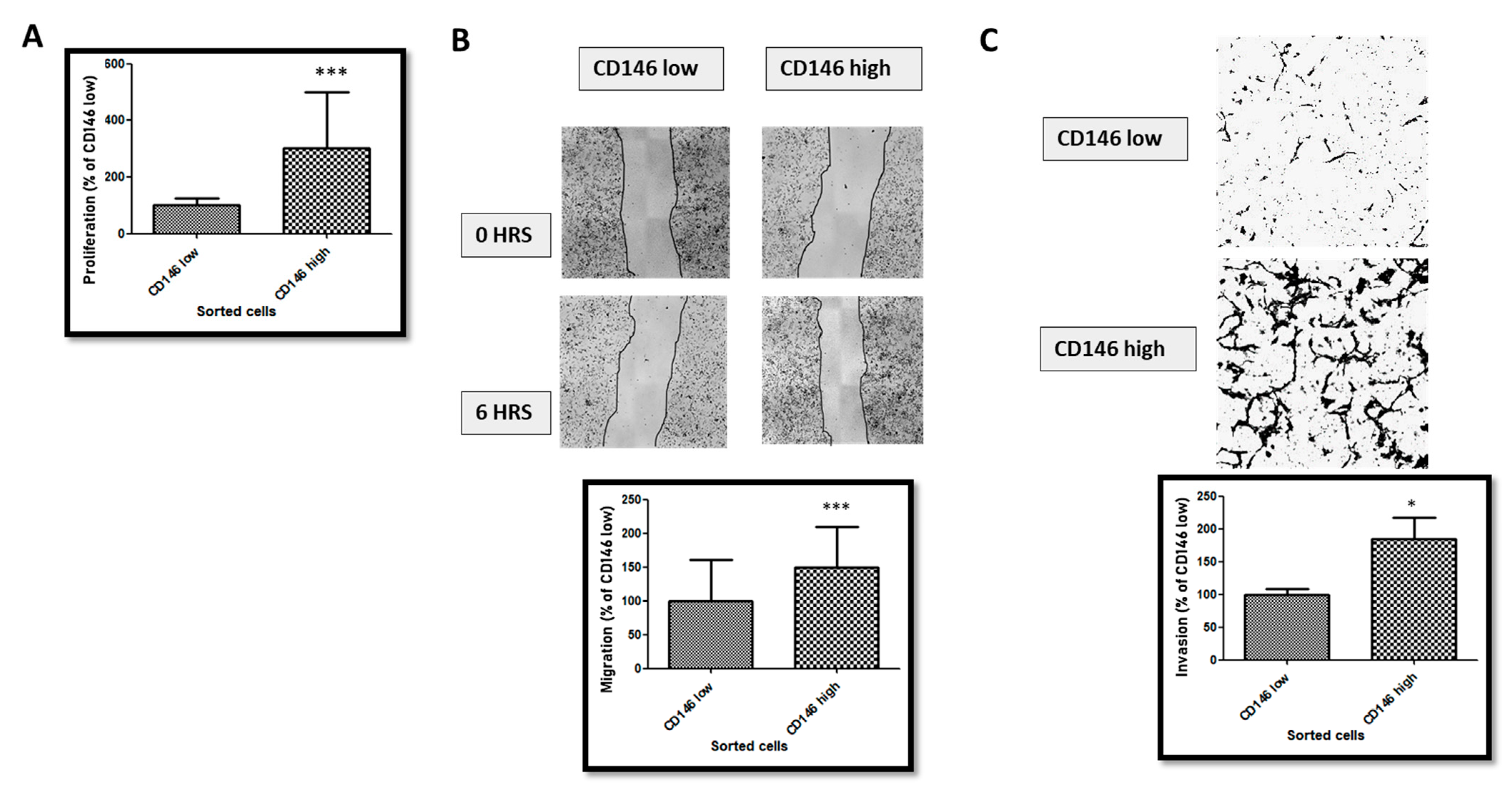
2.1.2. CD146-High MDA-MB-231 Cells Secrete a High Amount of Soluble CD146 and Display Invasive Properties
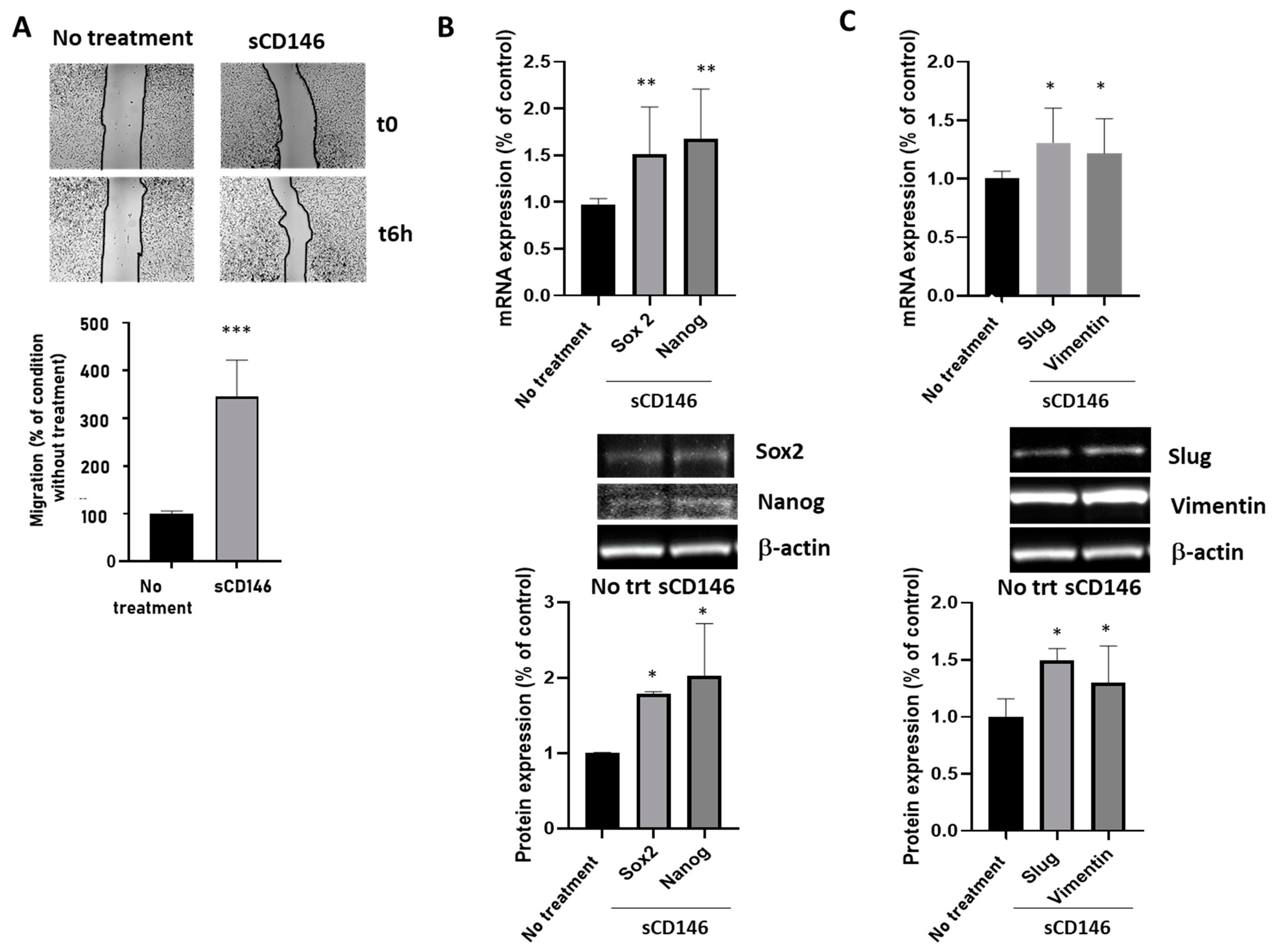
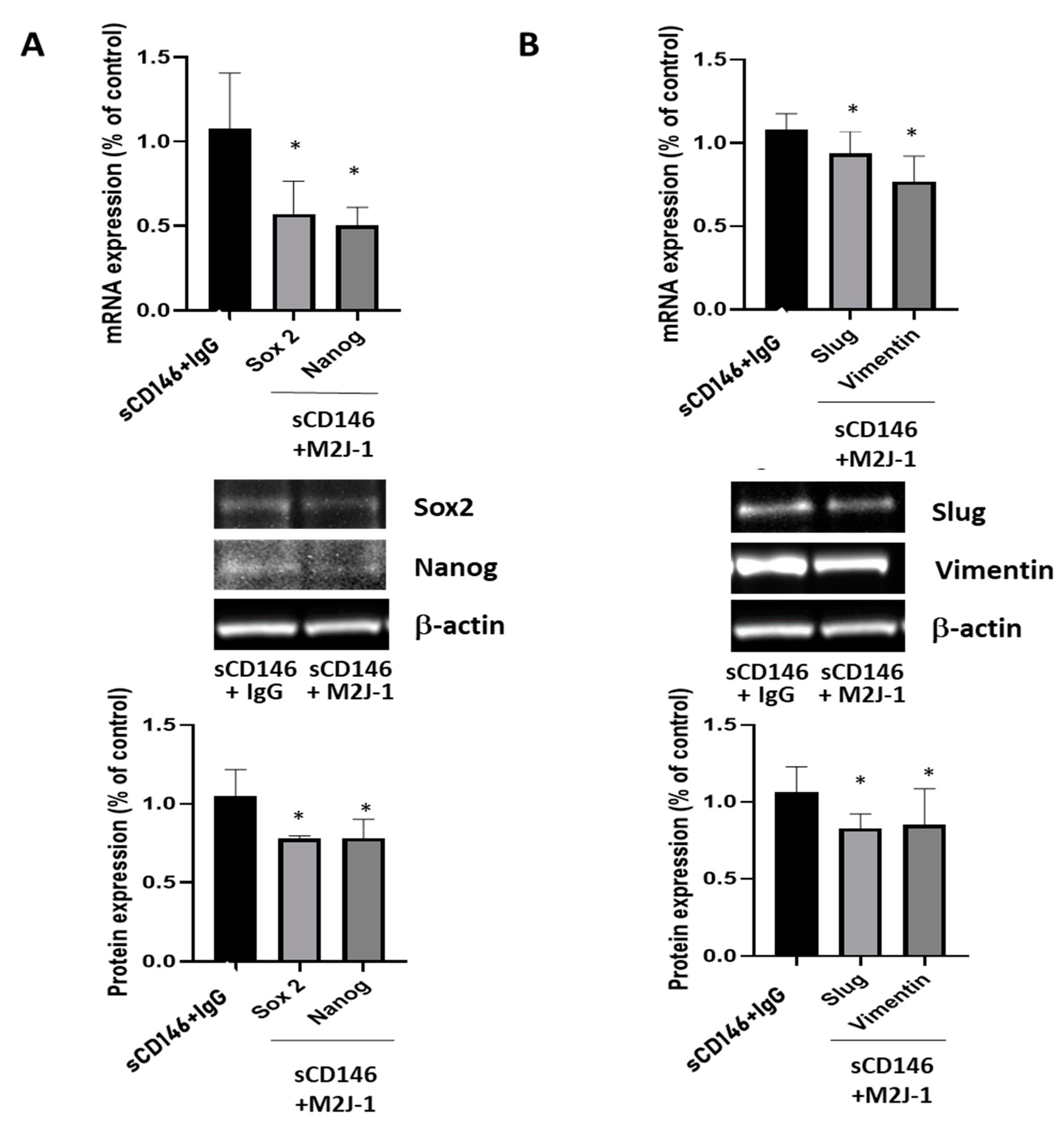
2.1.3. Soluble CD146 Stimulates MDA-MB 231 Cells Migration and Enhances Expression of Stem Cells and Epithelia-Mesenchymal Markers
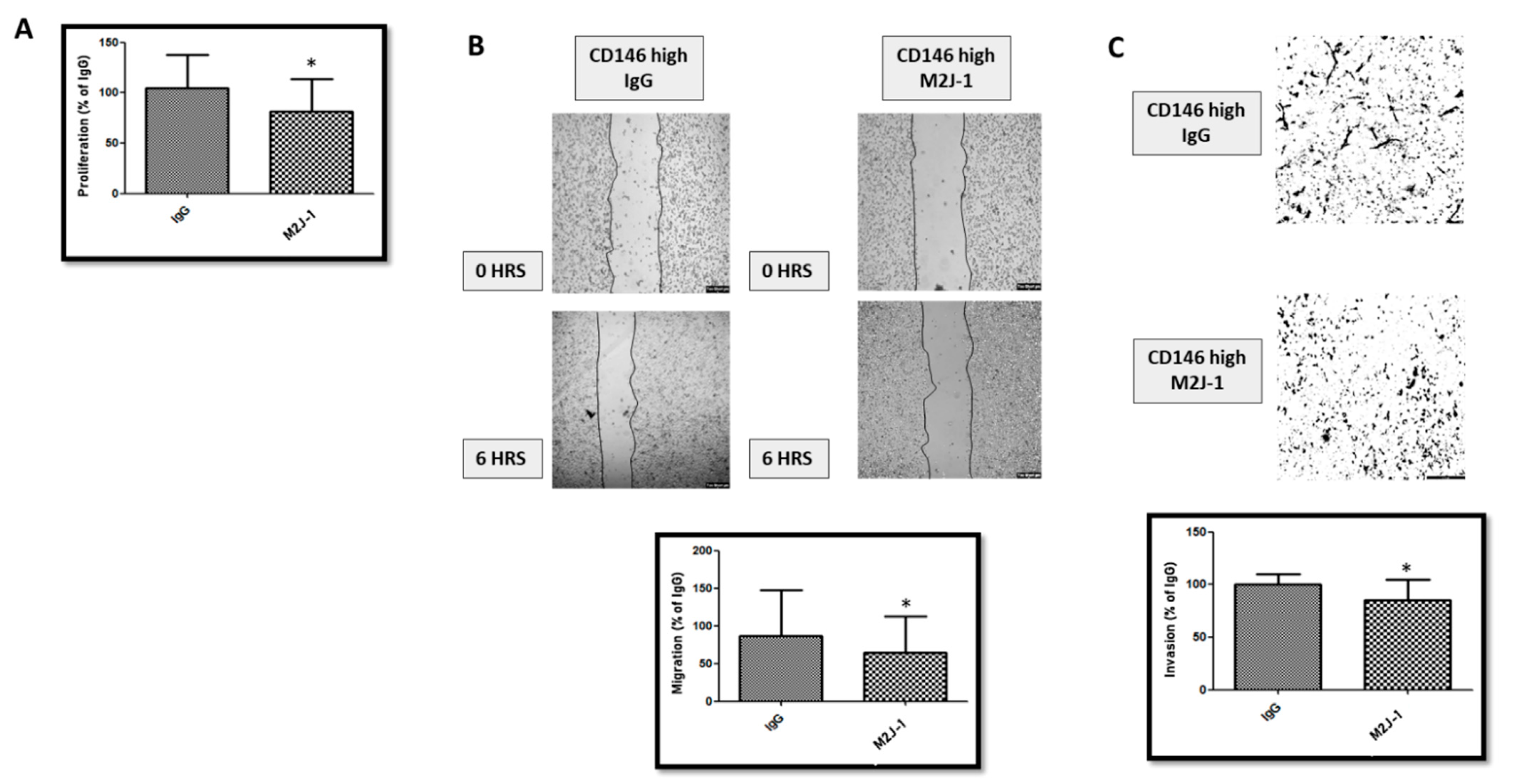
2.1.4. An Anti-sCD146 Monoclonal Antibody Blocks the Effects of sCD146 on MDA-MB 231 In Vitro
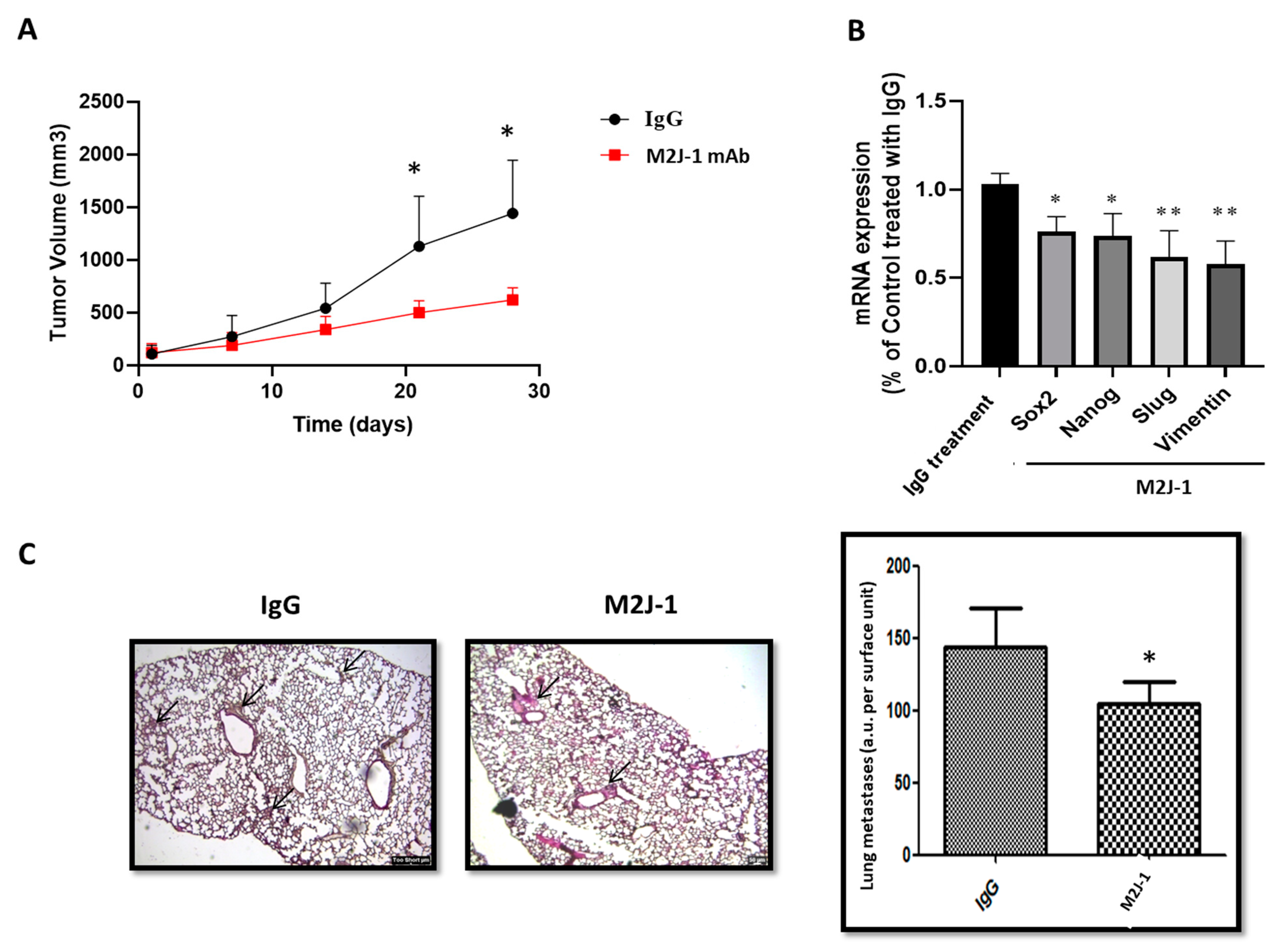
2.1.5. Anti-sCD146 mAb M2J-1 Prevents Tumor Growth and Metastatic Dissemination of MDA-MB-231 Cells in Two In Vivo Experimental Models
3. Discussion
4. Material and Methods
4.1. Cell Culture
4.2. Flow Cytometric Analysis and Cell Sorting
4.3. Western Blotting
4.4. Cell Proliferation Assay
4.5. Migration Assay
4.6. Cell Invasion Assay
4.7. qRT-PCR Experiments
4.8. Animal Studies
4.9. Histological Staining
4.10. Antibodies and ELISA Kit
4.11. Public Databases
4.12. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Lehmann, J.M.; Riethmuller, G.; Johnson, J.P. MUC18, a marker of tumor progression in human melanoma, shows sequence similarity to the neural cell adhesion molecules of the immunoglobulin superfamily. Proc. Natl. Acad. Sci. USA 1989, 86, 9891–9895. [Google Scholar] [CrossRef] [Green Version]
- Leroyer, A.S.; Blin, M.G.; Bachelier, R.; Bardin, N.; Blot-Chabaud, M.; Dignat-George, F. CD146 (Cluster of Differentiation 146). Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1026–1033. [Google Scholar] [CrossRef]
- Shih, I.M. The role of CD146 (Mel-CAM) in biology and pathology. J. Pathol. 1999, 189, 4–11. [Google Scholar] [CrossRef]
- Sacchetti, B.; Funari, A.; Michienzi, S.; Di Cesare, S.; Piersanti, S.; Saggio, I.; Tagliafico, E.; Ferrari, S.; Robey, P.G.; Riminucci, M.; et al. Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoietic microenvironment. Cell 2007, 131, 324–336. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.; Ohuchida, K.; Chijiiwa, Y.; Zhao, M.; Mizuuchi, Y.L.; Horioka, K.; Ohtsuka, T.; Mizumoto, K.; Oda, Y.; Hashizume, M.; et al. CD146 Attenuation in Cancer-Associated Fibroblasts Promotes Pancreatic Cancer Progression. Mol. Carcinog. 2016, 55, 1560–1572. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yang, D.; Wang, S.; Guo, X.; Lang, R.; Fan, Y.; Gu, F.; Zhang, X.; Niu, Y.; Yan, X.; et al. Increased expression of CD146 and microvessel density (MVD) in invasive micropapillary carcinoma of the breast: Comparative study with invasive ductal carcinoma-not otherwise specified. Pathol. Res. Pract. 2011, 207, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.J.; Peng, Q.; Fu, P.; Wang, S.W.; Chiang, C.F.; Dillehay, D.L.; Wu, M.W. Ectopical expression of human MUC18 increases metastasis of human prostate cancer cells. Gene 2004, 327, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Wu, Z.; Li, J.; Yang, X.; Wang, Y.; Yu, Y.; Ye, J.; Xu, C.; Qin, W.; Zhang, Z. MCAM is a novel metastasis marker and regulates spreading, apoptosis and invasion of ovarian cancer cells. Tumour Biol. 2012, 33, 1619–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, G.; Zhang, L.; Zhu, Q.; Bai, D.; Zhang, C.; Wang, X. CD146 promotes metastasis and predicts poor prognosis of hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2016, 35, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halt, K.; Parssinen, H.; Junttila, S.; Sim-Lucas, S.; Koivunen, P.; Myllyharju, J.; Quaggin, S.; Skovorodkin, I.; Vainio, S. CD146 cells are essential for kidney vasculature development. Kidney Int. 2016, 90, 311–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imbert, A.M.; Garulli, C.; Choquet, E.; Koubi, M.; Aurrand-Lions, M.; Chabannon, C. CD146 expression in human breast cancer cell lines induces phenotypic and functional changes observed in epithelial to mesenchymal transition. PLoS ONE 2012, 7, e43752. [Google Scholar] [CrossRef] [PubMed]
- Mani, S.A.; Guo, W.; Liao, M.J.; Eaton, E.N.; Ayyanan, A.; Zhou, A.Y.; Brooks, M.; Reinhard, F.; Zhang, C.C.; Shipitsin, M.; et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell 2008, 133, 704–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Q.; Li, W.; Lu, D.; Wu, Z.; Duan, H.; Luo, Y.; Feng, J.; Yang, D.; Fu, L.; Yan, X. CD146, an epithelial mesenchymal transition inducer, is associated with triple-negative breast cancer. Proc. Natl. Acad. Sci. USA 2012, 109, 1127–1132. [Google Scholar] [CrossRef] [Green Version]
- Zabouo, G.; Imbert, A.M.; Jacquemier, J.; Finetti, P.; Moreau, T.; Esterni, B.; Birnbaum, D.; Bertucci, F.; Chabannon, C. CD146 expression is associated with a poor prognosis in human breast tumors and with enhanced motility in breast cancer cell lines. Breast Cancer Res. 2009, 11, R1. [Google Scholar] [CrossRef] [Green Version]
- De Kruijff, I.; Timmermans, A.; den Bakker, M.; Trapman-Jansen, A.; Foekens, R.; Meijer-Van Gelder, M.; Oomen-de Hoop, E.; Smid, M.; Hollestelle, A.; van Deurzen, C.; et al. The Prevalence of CD146 Expression in Breast Cancer Subtypes and Its Relation to Outcome. Cancers 2018, 10, 134. [Google Scholar] [CrossRef] [Green Version]
- So, J.H.; Hong, S.K.; Kim, H.T.; Jung, S.H.; Lee, M.S.; Choi, J.H.; Bae, Y.K.; Kudoh, T.; Kim, J.H.; Kim, C.H. Gicerin/CD146 is involved in zebrafish cardiovascular development and tumor angiogenesis. Genes Cells 2010, 15, 1099–1110. [Google Scholar] [CrossRef]
- Stalin, J.; Traboulsi, W.; Vivancos-Stalin, L.; Nollet, M.; Joshkon, A.; Bachelier, R.; Guillet, B.; Lacroix, R.; Foucault-Bertaud, A.; Leroyer, A.; et al. Therapeutic targeting of soluble CD146/MCAM with the M2J-1 monoclonal antibody prevents metastasis development and procoagulant activity in CD146-positive invasive tumors. Int. J. Cancer 2020, 147, 1666–1679. [Google Scholar] [CrossRef]
- Schlagbauer-Wadl, H.; Jansen, B.; Muller, M.; Polterauer, P.; Wolff, K.; Eichler, H.G.; Pehamberger, H.; Konak, E.; Johnson, J.P. Influence of MUC18/MCAM/CD146 expression on human melanoma growth and metastasis in SCID mice. Int. J. Cancer 1999, 81, 951–955. [Google Scholar] [CrossRef]
- Tsuchiya, S.; Tsukamoto, Y.; Furuya, M.; Hiroi, S.; Miki, N.; Sasaki, F.; Taira, E. Gicerin, a cell adhesion molecule, promotes the metastasis of lymphoma cells of the chicken. Cell Tissue Res. 2003, 314, 389–397. [Google Scholar] [CrossRef]
- Tsukamoto, Y.; Sakaiuchi, T.; Hiroi, S.; Furuya, M.; Tsuchiya, S.; Sasaki, F.; Miki, N.; Taira, E. Expression of gicerin enhances the invasive and metastatic activities of a mouse mammary carcinoma cell line. Int. J. Oncol. 2003, 23, 1671–1677. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.J.; Fu, P.; Chiang, C.F.; Huss, W.J.; Greenberg, N.M.; Wu, M.W. Increased expression of MUC18 correlates with the metastatic progression of mouse prostate adenocarcinoma in the TRAMP model. J. Urol. 2005, 173, 1778–1783. [Google Scholar] [CrossRef]
- Aldovini, D.; Demichelis, F.; Doglioni, C.; Di Vizio, D.; Galligioni, E.; Brugnara, S.; Zeni, B.; Griso, C.; Pegoraro, C.; Zannoni, M.; et al. M-CAM expression as marker of poor prognosis in epithelial ovarian cancer. Int. J. Cancer 2006, 119, 1920–1926. [Google Scholar] [CrossRef]
- Kristiansen, G.; Yu, Y.; Schluns, K.; Sers, C.; Dietel, M.; Petersen, I. Expression of the cell adhesion molecule CD146/MCAM in non-small cell lung cancer. Anal. Cell Pathol. 2003, 25, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Oka, S.; Uramoto, H.; Chikaishi, Y.; Tanaka, F. The Expression of CD146 Predicts a Poor Overall Survival in Patients with Adenocarcinoma of the Lung. Anticancer Res. 2012, 32, 861–864. [Google Scholar]
- Ouellette, D.; Desbiens, G.; Emond, C.; Beauchamp, G. Lung cancer in women compared with men: Stage, treatment, and survival. Ann. Thorac. Surg. 1998, 66, 1140–1143. [Google Scholar] [CrossRef]
- Zeng, P.; Li, H.; Lu, P.; Zhou, L.; Tang, M.; Liu, C.; Chen, M. Prognostic value of CD146 in solid tumor: A Systematic Review and Meta-analysis. Sci. Rep. 2017, 7, 4223. [Google Scholar] [CrossRef]
- Shih, L.M.; Hsu, M.Y.; Palazzo, J.P.; Herlyn, M. The cell–cell adhesion receptor Mel-CAM acts as a tumor suppressor in breast carcinoma. Am. J. Pathol. 1997, 151, 745–751. [Google Scholar] [PubMed]
- Tripathi, S.C. MCAM mediates chemoresistance in small cell lung cancer via the PI3K/AKT/SOX2 signaling pathway. Cancer Res. 2017, 77, 4414–4425. [Google Scholar] [CrossRef] [Green Version]
- Stalin, J.; Nollet, M.; Garigue, P.; Fernandez, S.; Vivancos, L.; Essaadi, A.; Muller, A.; Bachelier, R.; Foucault-Bertaud, A.; Fugazza, L.; et al. Targeting soluble CD146 with a neutralizing antibody inhibits vascularization, growth and survival of CD146-positive tumors. Oncogene 2016, 35, 5489–5500. [Google Scholar] [CrossRef]
- Reinhold Arnold, C.; Mangesius, J.; Skvortsova, I.-I.; Ganswindt, U. The role of cancer stem cell in radiation resistance. Front. Oncol. 2020, 10, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostert, B.; Kraan, J.; Bolt-de Vries, J.; van der Spoel, P.; Sieuwerts, A.M.; Schutte, M.; Timmermans, A.M.; Foekens, R.; Martens, J.W.M.; Gratama, J.-W.; et al. Detection of circulating tumor cells in breast cancer may improve through enrichment with anti-CD146. Breast Cancer Res. Treat. 2010, 127, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaman, S.; Jadid, H.; Denson, A.; Gray, J. Targeting Trop-2 in solid tumors: Future prospects. OncoTargets Ther. 2019, 12, 1781–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Wambeke, S.; Gyawali, B. Atezolizumab in Metastatic Triple-Negative Breast Cancer—No Contradiction in the Eyes of a Dispassionate Observer. JAMA Oncol. 2021, 7, 1285–1286. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, A.; Joshkon, A.; Ladjimi, A.; Traboulsi, W.; Bachelier, R.; Robert, S.; Foucault-Bertaud, A.; Leroyer, A.S.; Bardin, N.; Somasundaram, I.; et al. Soluble CD146 as a Potential Target for Preventing Triple Negative Breast Cancer MDA-MB-231 Cell Growth and Dissemination. Int. J. Mol. Sci. 2022, 23, 974. https://doi.org/10.3390/ijms23020974
Sharma A, Joshkon A, Ladjimi A, Traboulsi W, Bachelier R, Robert S, Foucault-Bertaud A, Leroyer AS, Bardin N, Somasundaram I, et al. Soluble CD146 as a Potential Target for Preventing Triple Negative Breast Cancer MDA-MB-231 Cell Growth and Dissemination. International Journal of Molecular Sciences. 2022; 23(2):974. https://doi.org/10.3390/ijms23020974
Chicago/Turabian StyleSharma, Akshita, Ahmad Joshkon, Aymen Ladjimi, Waël Traboulsi, Richard Bachelier, Stéphane Robert, Alexandrine Foucault-Bertaud, Aurélie S. Leroyer, Nathalie Bardin, Indumathi Somasundaram, and et al. 2022. "Soluble CD146 as a Potential Target for Preventing Triple Negative Breast Cancer MDA-MB-231 Cell Growth and Dissemination" International Journal of Molecular Sciences 23, no. 2: 974. https://doi.org/10.3390/ijms23020974