
The Broad-Spectrum Antiviral Potential of the Amphibian Peptide AR-23

 , , , and
, , , and
Abstract
:
1. Introduction
2. Results
2.1. Cytotoxicity evaluation
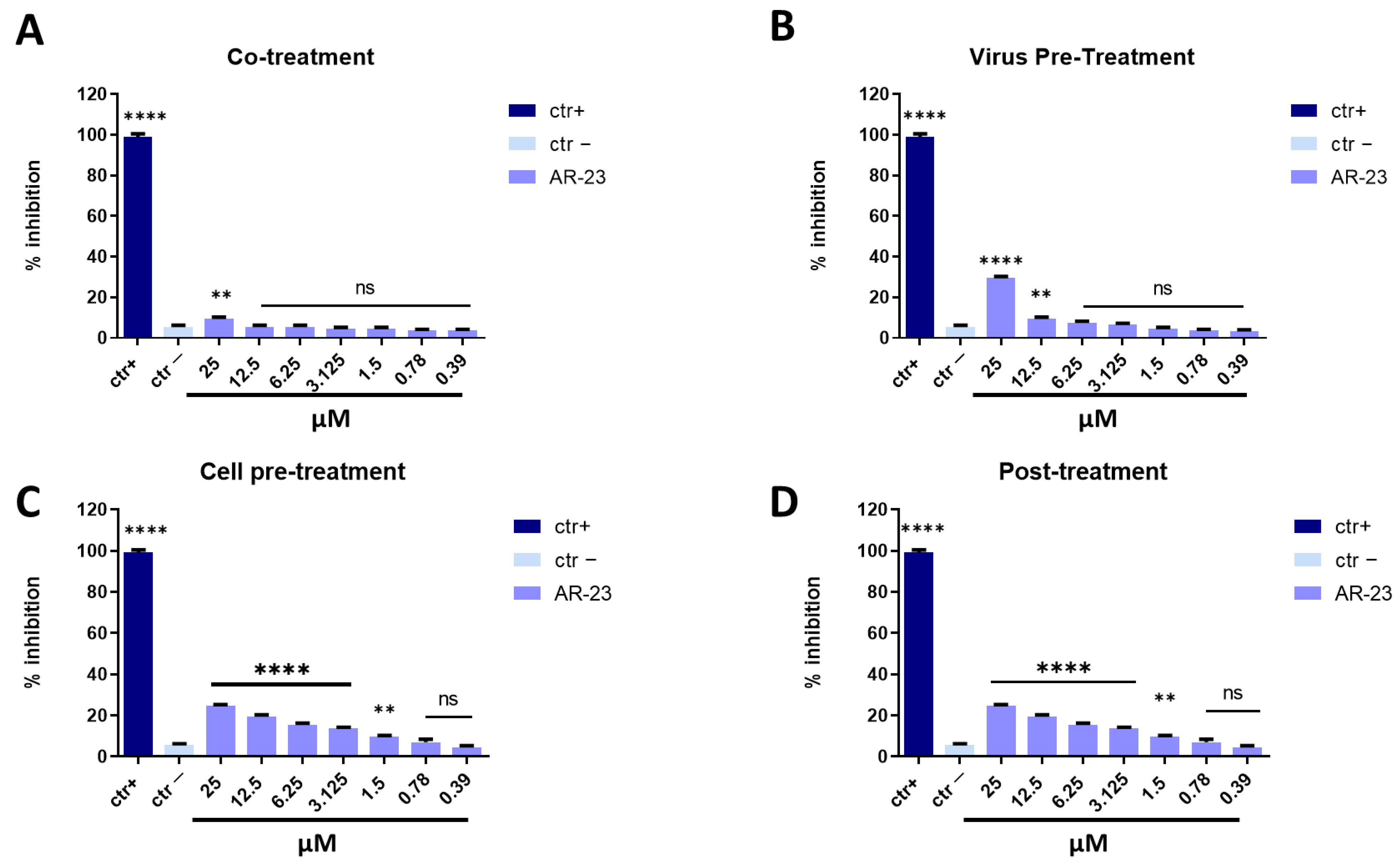
2.2. Evaluation of Antiviral Activity through Plaque Assay
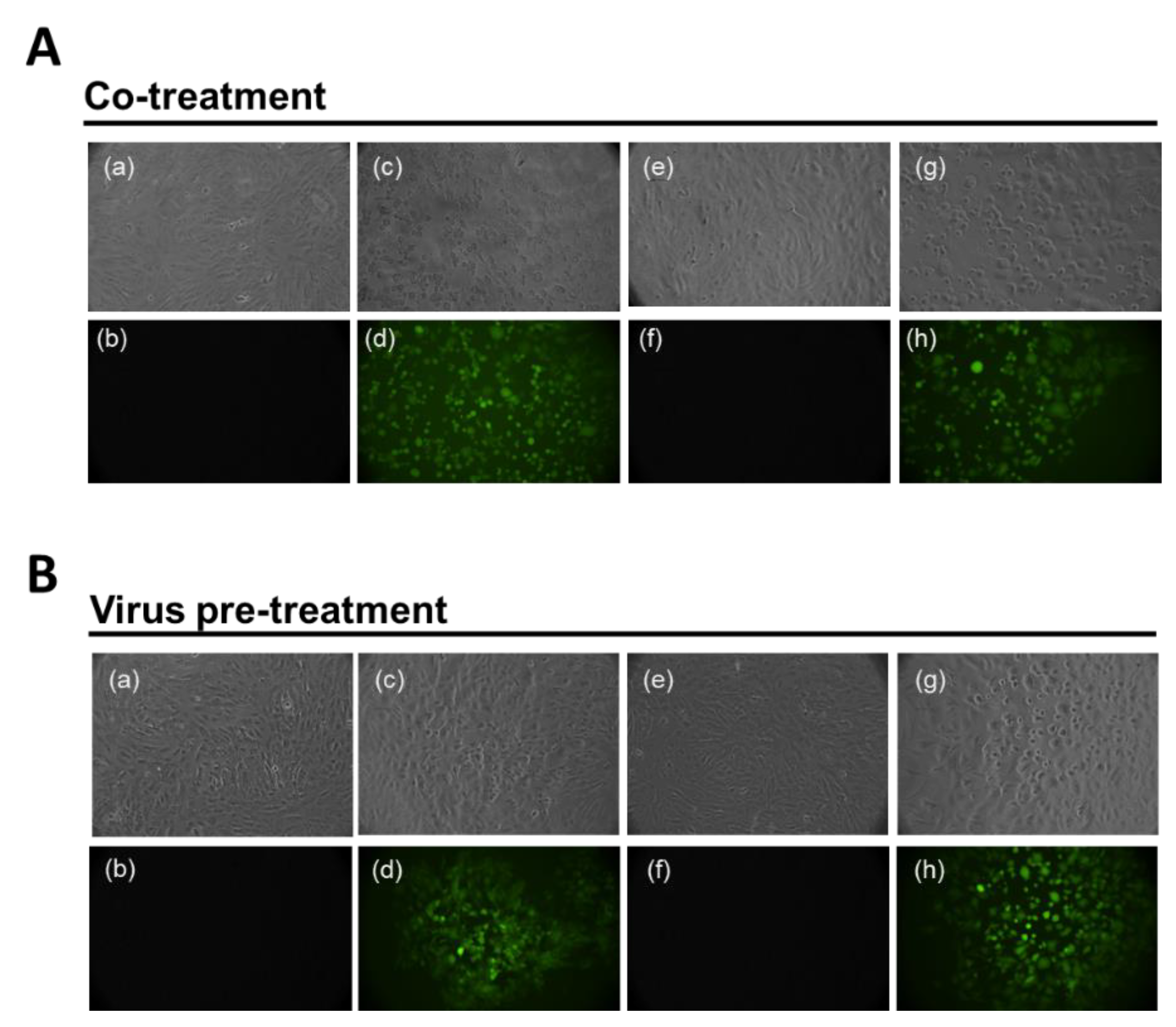
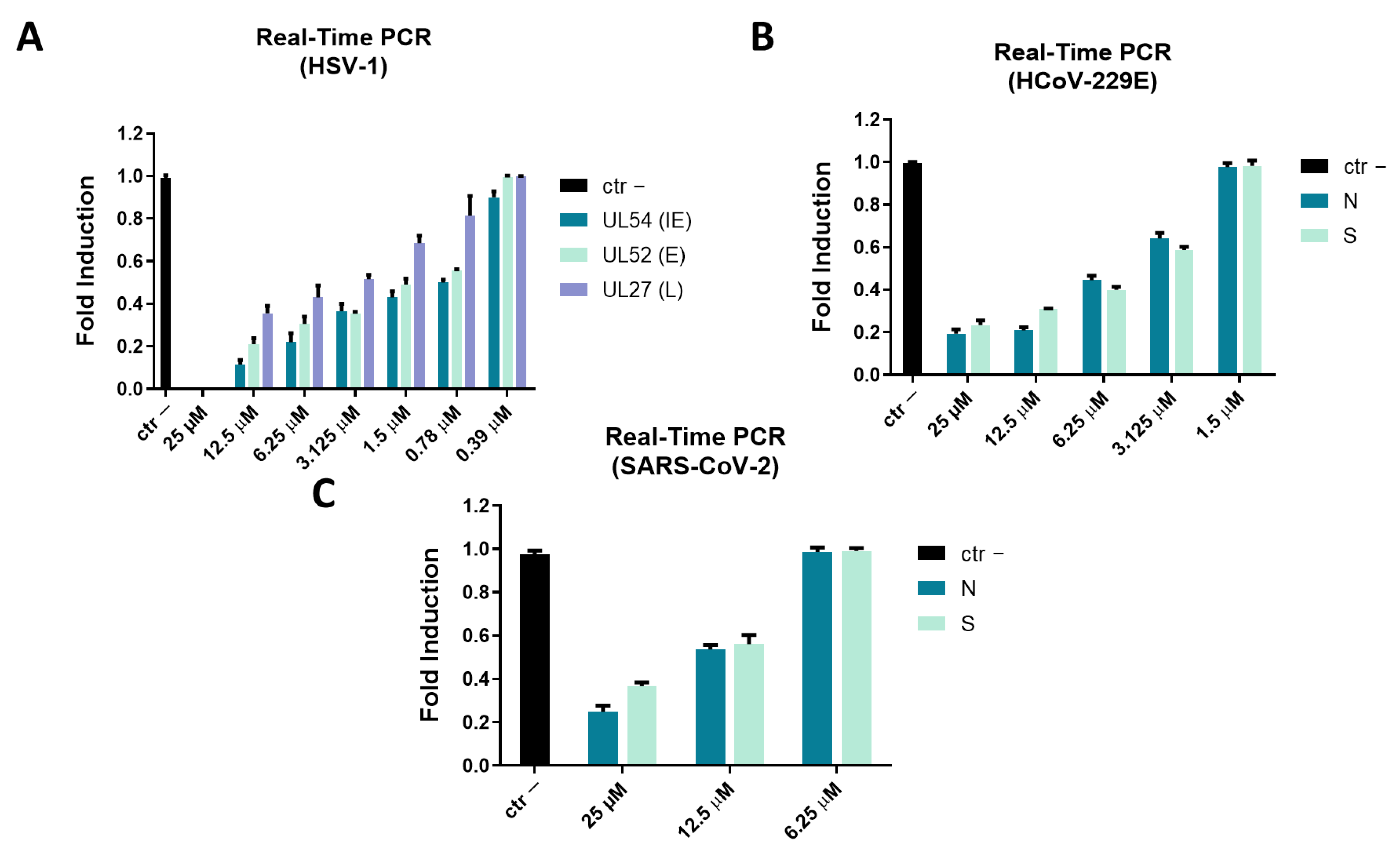
2.2.1. Inhibitory Activity against DNA Virus: HSV-1
2.2.2. Inhibitory Activity against RNA Virus:
Paramyxoviridae
Coronaviridae
Picornaviridae
2.3. Molecular Assay: Real-Time PCR
2.4. Morphological Analysis of AR-23 by Transmission Electron Microscope (TEM)
3. Discussion
4. Materials and Methods
4.1. Peptide Synthesis and Characterization
4.2. Cell Culture and Viral Strain
4.3. Cytotoxic Activity
4.4. Antiviral Activity
- (a)
- Co-treatment assay: cells were inoculated with several concentrations of peptide (from 100 to 0.78 μM) and virus at MOI 0.01 for 1 h at 37 °C/5% CO2.
- (b)
- Virus pre-treatment assay: peptide was incubated together with virus at MOI 0.1, for 1 h at 37 °C. After that, the mixture was inoculated on the cells for 1 h at 37 °C/5% CO2.
- (c)
- Cell pre-treatment assay: cells were first treated with peptide for 1 h, after that AR-23 was removed and cells were infected with virus (MOI 0.01) for another hour at 37 °C/5% CO2.
- (d)
- Post-treatment assay: cells were infected with virus (MOI 0.01) for 1 h and, subsequently, peptide was added for another hour at 37 °C/5% CO2.
- (e)
- Attachment assay: Vero cells were seeded in 24-well plates at the density of 2.5 × 105 cells/mL for 24 h. Then, they were infected with HSV-1 (MOI 0.01) and at same time treated with AR-23 from 25 to 0.39 μM for 1 h at 4 °C. Subsequently, the cell monolayer was washed and incubated for 1 h at 37 °C with a culture medium. Then, cells were washed with PBS and DMEM/CMC was added for 48 h at 37 °C/5% CO2.
- (f)
- Entry assay: Vero cells were plated in 24-well plates as indicated above and then infected with HSV-1 (MOI 0.01) for 1 h at 4 °C. Subsequently, AR-23 was put on cells for 1 h at 37 °C and finally cells were washed with citrate buffer (pH 3) and overlaid with DMEM/CMC.
4.5. Analysis of Gene Expression
4.6. Virus Purification and TEM Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lai, C.C.; Shih, T.P.; Ko, W.C.; Tang, H.J.; Hsueh, P.R. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and coronavirus disease-2019 (COVID-19): The epidemic and the challenges. Int. J. Antimicrob. Agents 2020, 55, 105924. [Google Scholar] [CrossRef] [PubMed]
- WHO. Coronavirus Disease (COVID-19) Pandemic. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019 (accessed on 9 December 2021).
- WHO. Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 9 December 2021).
- De Angelis, M.; Casciaro, B.; Genovese, A.; Brancaccio, D.; Marcocci, M.E.; Novellino, E.; Carotenuto, A.; Palamara, A.T.; Mangoni, M.L.; Nencioni, L. Temporin G, an amphibian antimicrobial peptide against influenza and parainfluenza respiratory viruses: Insights into biological activity and mechanism of action. FASEB J. 2021, 35, e21358. [Google Scholar] [CrossRef] [PubMed]
- WHO. Influenza (Seasonal). Available online: https://www.who.int/news-room/fact-sheets/detail/influenza-(seasonal) (accessed on 9 December 2021).
- CDC. Parainfluenza Types 1–3 National Trends. Available online: https://www.cdc.gov/surveillance/nrevss/human-paraflu/natl-trend.html (accessed on 9 December 2021).
- Saxena, S.K.; Mishra, N.; Saxena, R. Advances in antiviral drug discovery and development: Part II: Advancements in antiviral drug development. Future Virol. 2009, 43. [Google Scholar] [CrossRef]
- Memariani, H.; Memariani, M.; Moravvej, H.; Shahidi-Dadras, M. Melittin: A venom-derived peptide with promising anti-viral properties. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 5–17. [Google Scholar] [CrossRef]
- Jiang, Y.C.; Feng, H.; Lin, Y.C.; Guo, X.R. New strategies against drug resistance to herpes simplex virus. Int. J. Oral Sci. 2016, 8, 1–6. [Google Scholar] [CrossRef]
- Marcocci, M.E.; Amatore, D.; Villa, S.; Casciaro, B.; Aimola, P.; Franci, G.; Grieco, P.; Galdiero, M.; Palamara, A.T.; Mangoni, M.L.; et al. The Amphibian Antimicrobial Peptide Temporin B Inhibits In Vitro Herpes Simplex Virus 1 Infection. Antimicrob. Agents Chemother. 2018, 62, e02367-17. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.S.; Prodhan, Z.H.; Biswas, S.K.; Le, C.F.; Sekaran, S.D. Antimicrobial peptides from different plant sources: Isolation, characterisation, and purification. Phytochemistry 2018, 154, 94–105. [Google Scholar] [CrossRef]
- Bulet, P.; Hetru, C.; Dimarcq, J.L.; Hoffmann, D. Antimicrobial peptides in insects; structure and function. Dev. Comp. Immunol. 1999, 23, 329–344. [Google Scholar] [CrossRef]
- Memariani, H.; Memariani, M.; Pourmand, M.R. Venom-derived peptide Mastoparan-1 eradicates planktonic and biofilm-embedded methicillin-resistant Staphylococcus aureus isolates. Microb. Pathog. 2018, 119, 72–80. [Google Scholar] [CrossRef]
- Carvalho Ade, O.; Gomes, V.M. Plant defensins—Prospects for the biological functions and biotechnological properties. Peptides 2009, 30, 1007–1020. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef]
- Conlon, J.M.; Mechkarska, M. Host-defense peptides with therapeutic potential from skin secretions of frogs from the family pipidae. Pharmaceuticals 2014, 7, 58–77. [Google Scholar] [CrossRef] [Green Version]
- Albiol Matanic, V.C.; Castilla, V. Antiviral activity of antimicrobial cationic peptides against Junin virus and herpes simplex virus. Int. J. Antimicrob. Agents 2004, 23, 382–389. [Google Scholar] [CrossRef]
- Ohsaki, Y.; Gazdar, A.F.; Chen, H.C.; Johnson, B.E. Antitumor activity of magainin analogues against human lung cancer cell lines. Cancer Res. 1992, 52, 3534–3538. [Google Scholar]
- Lehmann, J.; Retz, M.; Sidhu, S.S.; Suttmann, H.; Sell, M.; Paulsen, F.; Harder, J.; Unteregger, G.; Stockle, M. Antitumor activity of the antimicrobial peptide magainin II against bladder cancer cell lines. Eur. Urol. 2006, 50, 141–147. [Google Scholar] [CrossRef]
- Kim, M.K.; Kang, N.; Ko, S.J.; Park, J.; Park, E.; Shin, D.W.; Kim, S.H.; Lee, S.A.; Lee, J.I.; Lee, S.H.; et al. Antibacterial and Antibiofilm Activity and Mode of Action of Magainin 2 against Drug-Resistant Acinetobacter baumannii. Int. J. Mol. Sci. 2018, 19, 3041. [Google Scholar] [CrossRef] [Green Version]
- VanCompernolle, S.E.; Taylor, R.J.; Oswald-Richter, K.; Jiang, J.; Youree, B.E.; Bowie, J.H.; Tyler, M.J.; Conlon, J.M.; Wade, D.; Aiken, C.; et al. Antimicrobial peptides from amphibian skin potently inhibit human immunodeficiency virus infection and transfer of virus from dendritic cells to T cells. J. Virol. 2005, 79, 11598–11606. [Google Scholar] [CrossRef] [Green Version]
- Bartels, E.J.H.; Dekker, D.; Amiche, M. Dermaseptins, Multifunctional Antimicrobial Peptides: A Review of Their Pharmacology, Effectivity, Mechanism of Action, and Possible Future Directions. Front. Pharmacol. 2019, 10, 1421. [Google Scholar] [CrossRef] [Green Version]
- Stelitano, D.; Franci, G.; Chianese, A.; Galdiero, S.; Morelli, G.; Galdiero, M. HSV membrane glycoproteins, their function in viral entry and their use in vaccine studies. In Amino Acids, Peptides and Proteins; Royal Society of Chemistry: London, UK, 2019; pp. 14–43. [Google Scholar]
- Tulassay, T.; Rascher, W. Atrial natriuretic peptide. New aspects in the regulation of blood volume and electrolyte homeostasis. Monatsschr. Kinderheilkd. 1986, 134, 710–715. [Google Scholar]
- Roy, M.; Lebeau, L.; Chessa, C.; Damour, A.; Ladram, A.; Oury, B.; Boutolleau, D.; Bodet, C.; Leveque, N. Comparison of Anti-Viral Activity of Frog Skin Anti-Microbial Peptides Temporin-Sha and [K(3)]SHa to LL-37 and Temporin-Tb against Herpes Simplex Virus Type 1. Viruses 2019, 11, 77. [Google Scholar] [CrossRef] [Green Version]
- Conlon, J.M.; Sonnevend, A.; Patel, M.; Camasamudram, V.; Nowotny, N.; Zilahi, E.; Iwamuro, S.; Nielsen, P.F.; Pal, T. A melittin-related peptide from the skin of the Japanese frog, Rana tagoi, with antimicrobial and cytolytic properties. Biochem. Biophys. Res. Commun. 2003, 306, 496–500. [Google Scholar] [CrossRef]
- Urban, E.; Nagy, E.; Pal, T.; Sonnevend, A.; Conlon, J.M. Activities of four frog skin-derived antimicrobial peptides (temporin-1DRa, temporin-1Va and the melittin-related peptides AR-23 and RV-23) against anaerobic bacteria. Int. J. Antimicrob. Agents 2007, 29, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.K.; Song, J.W.; Gong, F.; Li, S.B.; Chang, H.Y.; Xie, H.M.; Gao, H.W.; Tan, Y.X.; Ji, S.P. Design of an alpha-helical antimicrobial peptide with improved cell-selective and potent anti-biofilm activity. Sci. Rep. 2016, 6, 27394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galdiero, E.; Siciliano, A.; Gesuele, R.; Di Onofrio, V.; Falanga, A.; Maione, A.; Liguori, R.; Libralato, G.; Guida, M. Melittin Inhibition and Eradication Activity for Resistant Polymicrobial Biofilm Isolated from a Dairy Industry after Disinfection. Int. J. Microbiol. 2019, 2019, 4012394. [Google Scholar] [CrossRef]
- Baghian, A.; Kousoulas, K.G. Role of the Na+, K+ pump in herpes simplex type 1-induced cell fusion: Melittin causes specific reversion of syncytial mutants with the syn1 mutation to Syn+ (wild-type) phenotype. Virology 1993, 196, 548–556. [Google Scholar] [CrossRef]
- Uddin, M.B.; Lee, B.H.; Nikapitiya, C.; Kim, J.H.; Kim, T.H.; Lee, H.C.; Kim, C.G.; Lee, J.S.; Kim, C.J. Inhibitory effects of bee venom and its components against viruses in vitro and in vivo. J. Microbiol. 2016, 54, 853–866. [Google Scholar] [CrossRef]
- Wachinger, M.; Saermark, T.; Erfle, V. Influence of amphipathic peptides on the HIV-1 production in persistently infected T lymphoma cells. FEBS Lett. 1992, 309, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.H.; Jang, A.Y.; Lin, S.; Lim, S.; Kim, D.; Park, K.; Han, S.M.; Yeo, J.H.; Seo, H.S. Melittin, a honeybee venomderived antimicrobial peptide, may target methicillinresistant Staphylococcus aureus. Mol. Med. Rep. 2015, 12, 6483–6490. [Google Scholar] [CrossRef] [Green Version]
- Dosler, S.; Karaaslan, E.; Alev Gerceker, A. Antibacterial and anti-biofilm activities of melittin and colistin, alone and in combination with antibiotics against Gram-negative bacteria. J. Chemother. 2016, 28, 95–103. [Google Scholar] [CrossRef]
- Falanga, A.; Del Genio, V.; Kaufman, E.A.; Zannella, C.; Franci, G.; Weck, M.; Galdiero, S. Engineering of Janus-Like Dendrimers with Peptides Derived from Glycoproteins of Herpes Simplex Virus Type 1: Toward a Versatile and Novel Antiviral Platform. Int. J. Mol. Sci. 2021, 22, 6488. [Google Scholar] [CrossRef]
- Tai, C.J.; Li, C.L.; Tai, C.J.; Wang, C.K.; Lin, L.T. Early Viral Entry Assays for the Identification and Evaluation of Antiviral Compounds. J. Vis. Exp. 2015, 104, e53124. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Zannella, C.; Folliero, V.; Di Girolamo, R.; Bajardi, F.; Chianese, A.; Altucci, L.; Damasco, A.; Del Sorbo, M.R.; Imperatore, C.; et al. Combating Actions of Green 2D-Materials on Gram Positive and Negative Bacteria and Enveloped Viruses. Front. Bioeng. Biotechnol. 2020, 8, 569967. [Google Scholar] [CrossRef]
- Laksono, B.M.; de Vries, R.D.; McQuaid, S.; Duprex, W.P.; de Swart, R.L. Measles Virus Host Invasion and Pathogenesis. Viruses 2016, 8, 210. [Google Scholar] [CrossRef] [Green Version]
- Branche, A.R.; Falsey, A.R. Parainfluenza Virus Infection. Semin. Respir. Crit. Care Med. 2016, 37, 538–554. [Google Scholar] [CrossRef]
- Li, Q.; Zhao, Z.; Zhou, D.; Chen, Y.; Hong, W.; Cao, L.; Yang, J.; Zhang, Y.; Shi, W.; Cao, Z.; et al. Virucidal activity of a scorpion venom peptide variant mucroporin-M1 against measles, SARS-CoV and influenza H5N1 viruses. Peptides 2011, 32, 1518–1525. [Google Scholar] [CrossRef]
- Ha, M.N.; Delpeut, S.; Noyce, R.S.; Sisson, G.; Black, K.M.; Lin, L.T.; Bilimoria, D.; Plemper, R.K.; Prive, G.G.; Richardson, C.D. Mutations in the Fusion Protein of Measles Virus That Confer Resistance to the Membrane Fusion Inhibitors Carbobenzoxy-d-Phe-l-Phe-Gly and 4-Nitro-2-Phenylacetyl Amino-Benzamide. J. Virol. 2017, 91, e01026-17. [Google Scholar] [CrossRef] [Green Version]
- Barnard, D.L.; Stowell, V.D.; Seley, K.L.; Hegde, V.R.; Das, S.R.; Rajappan, V.P.; Schneller, S.W.; Smee, D.F.; Sidwell, R.W. Inhibition of measles virus replication by 5′-nor carbocyclic adenosine analogues. Antivir. Chem. Chemother. 2001, 12, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Giugliano, R.; Buonocore, C.; Zannella, C.; Chianese, A.; Palma Esposito, F.; Tedesco, P.; De Filippis, A.; Galdiero, M.; Franci, G.; de Pascale, D. Antiviral Activity of the Rhamnolipids Mixture from the Antarctic Bacterium Pseudomonas gessardii M15 against Herpes Simplex Viruses and Coronaviruses. Pharmaceutics 2021, 13, 2121. [Google Scholar] [CrossRef]
- Cheng, P.W.; Ng, L.T.; Chiang, L.C.; Lin, C.C. Antiviral effects of saikosaponins on human coronavirus 229E in vitro. Clin. Exp. Pharmacol. Physiol. 2006, 33, 612–616. [Google Scholar] [CrossRef]
- Tan, Y.L.; Tan, K.S.W.; Chu, J.J.H.; Chow, V.T. Combination Treatment with Remdesivir and Ivermectin Exerts Highly Synergistic and Potent Antiviral Activity Against Murine Coronavirus Infection. Front. Cell Infect. Microbiol. 2021, 11, 700502. [Google Scholar] [CrossRef]
- Sanna, G.; Madeddu, S.; Serra, A.; Collu, D.; Efferth, T.; Hakkim, F.L.; Rashan, L. Anti-poliovirus activity of Nerium oleander aqueous extract. Nat. Prod. Res. 2021, 35, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Dove, A.W.; Racaniello, V.R. An antiviral compound that blocks structural transitions of poliovirus prevents receptor binding at low temperatures. J. Virol. 2000, 74, 3929–3931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.Y.; Lyu, Y.N.; Xin, H.Y.; Cheng, J.T.; Liu, X.Q.; Wang, X.W.; Peng, X.C.; Xiang, Y.; Xin, V.W.; Lu, C.B.; et al. Identification of Putative UL54 (ICP27) Transcription Regulatory Sequences Binding to Oct-1, v-Myb, Pax-6 and Hairy in Herpes Simplex Viruses. J. Cancer 2019, 10, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Cubuk, J.; Alston, J.J.; Incicco, J.J.; Singh, S.; Stuchell-Brereton, M.D.; Ward, M.D.; Zimmerman, M.I.; Vithani, N.; Griffith, D.; Wagoner, J.A.; et al. The SARS-CoV-2 nucleocapsid protein is dynamic, disordered, and phase separates with RNA. Nat. Commun. 2021, 12, 1936. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.L.; Hodges, R.S. Structure-activity relationships of de novo designed cyclic antimicrobial peptides based on gramicidin S. Biopolymers 2003, 71, 28–48. [Google Scholar] [CrossRef] [PubMed]
- Duffalo, M.L.; James, C.W. Enfuvirtide: A novel agent for the treatment of HIV-1 infection. Ann. Pharmacother. 2003, 37, 1448–1456. [Google Scholar] [CrossRef]
- Lalezari, J.P.; Luber, A.D. Enfuvirtide. Drugs Today 2004, 40, 259–269. [Google Scholar] [CrossRef]
- Caporale, A.; Doti, N.; Monti, A.; Sandomenico, A.; Ruvo, M. Automatic procedures for the synthesis of difficult peptides using oxyma as activating reagent: A comparative study on the use of bases and on different deprotection and agitation conditions. Peptides 2018, 102, 38–46. [Google Scholar] [CrossRef]
- Brignati, M.J.; Loomis, J.S.; Wills, J.W.; Courtney, R.J. Membrane association of VP22, a herpes simplex virus type 1 tegument protein. J. Virol. 2003, 77, 4888–4898. [Google Scholar] [CrossRef] [Green Version]
- Petrillo, F.; Chianese, A.; De Bernardo, M.; Zannella, C.; Galdiero, M.; Reibaldi, M.; Avitabile, T.; Boccia, G.; Galdiero, M.; Rosa, N.; et al. Inhibitory Effect of Ophthalmic Solutions against SARS-CoV-2: A Preventive Action to Block the Viral Transmission? Microorganisms 2021, 9, 1550. [Google Scholar] [CrossRef]
- Szelechowski, M.; Bergeron, C.; Gonzalez-Dunia, D.; Klonjkowski, B. Production and purification of non replicative canine adenovirus type 2 derived vectors. J. Vis. Exp. 2013, 82, 50833. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Forward Sequence | Reverse Sequence |
|---|---|---|
| HSV-1 UL54 | TGGCGGACATTAAGGACATTG | TGGCCGTCAACTCGCAG |
| HSV-1 UL52 | GACCGACGGGTGCGTTATT | GAAGGAGTCGCCATTTAGCC |
| HSV-1 UL27 | GCCTTCTTCGCCTTTCGC | CGCTCGTGCCCTTCTTCTT |
| HCoV-229E S | CGTTGAACTTCAAACCTCAGA | ACCAACATTGGCATAAACAG |
| HCoV-229E N | GTCGTCAGGGGTAGAATACCTTA | CCCGTTTGCCCTTTCTAGT |
| SARS-CoV-2 S | AGGTTGATCACAGGCAGACT | GCTGACTGAGGGAAGGAC |
| SARS-CoV-2 N | GGGGAACTTCTCCTGCTAGAAT | CAGACATTTTGCTCTCAAGCTG |
| GAPDH | CCTTTCATTGAGCTCCAT | CGTACATGGGAGCGTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chianese, A.; Zannella, C.; Monti, A.; De Filippis, A.; Doti, N.; Franci, G.; Galdiero, M. The Broad-Spectrum Antiviral Potential of the Amphibian Peptide AR-23. Int. J. Mol. Sci. 2022, 23, 883. https://doi.org/10.3390/ijms23020883
Chianese A, Zannella C, Monti A, De Filippis A, Doti N, Franci G, Galdiero M. The Broad-Spectrum Antiviral Potential of the Amphibian Peptide AR-23. International Journal of Molecular Sciences. 2022; 23(2):883. https://doi.org/10.3390/ijms23020883
Chicago/Turabian StyleChianese, Annalisa, Carla Zannella, Alessandra Monti, Anna De Filippis, Nunzianna Doti, Gianluigi Franci, and Massimiliano Galdiero. 2022. "The Broad-Spectrum Antiviral Potential of the Amphibian Peptide AR-23" International Journal of Molecular Sciences 23, no. 2: 883. https://doi.org/10.3390/ijms23020883