
Supported Lipid Bilayer Platform for Characterizing the Membrane-Disruptive Behaviors of Triton X-100 and Potential Detergent Replacements



Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
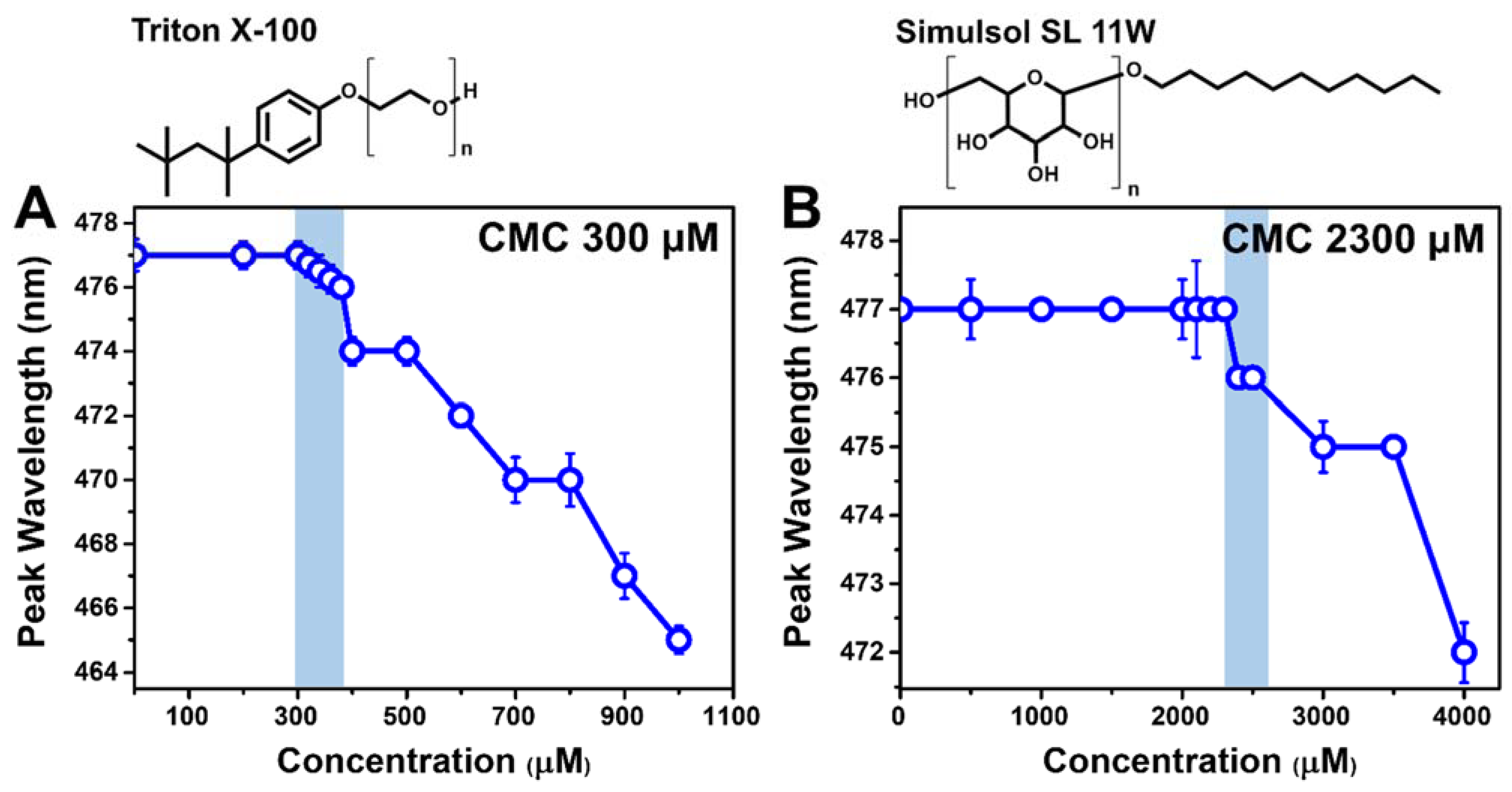
2.1. Micellar Aggregation Properties
2.2. QCM-D Tracking of Detergent-SLB Interactions
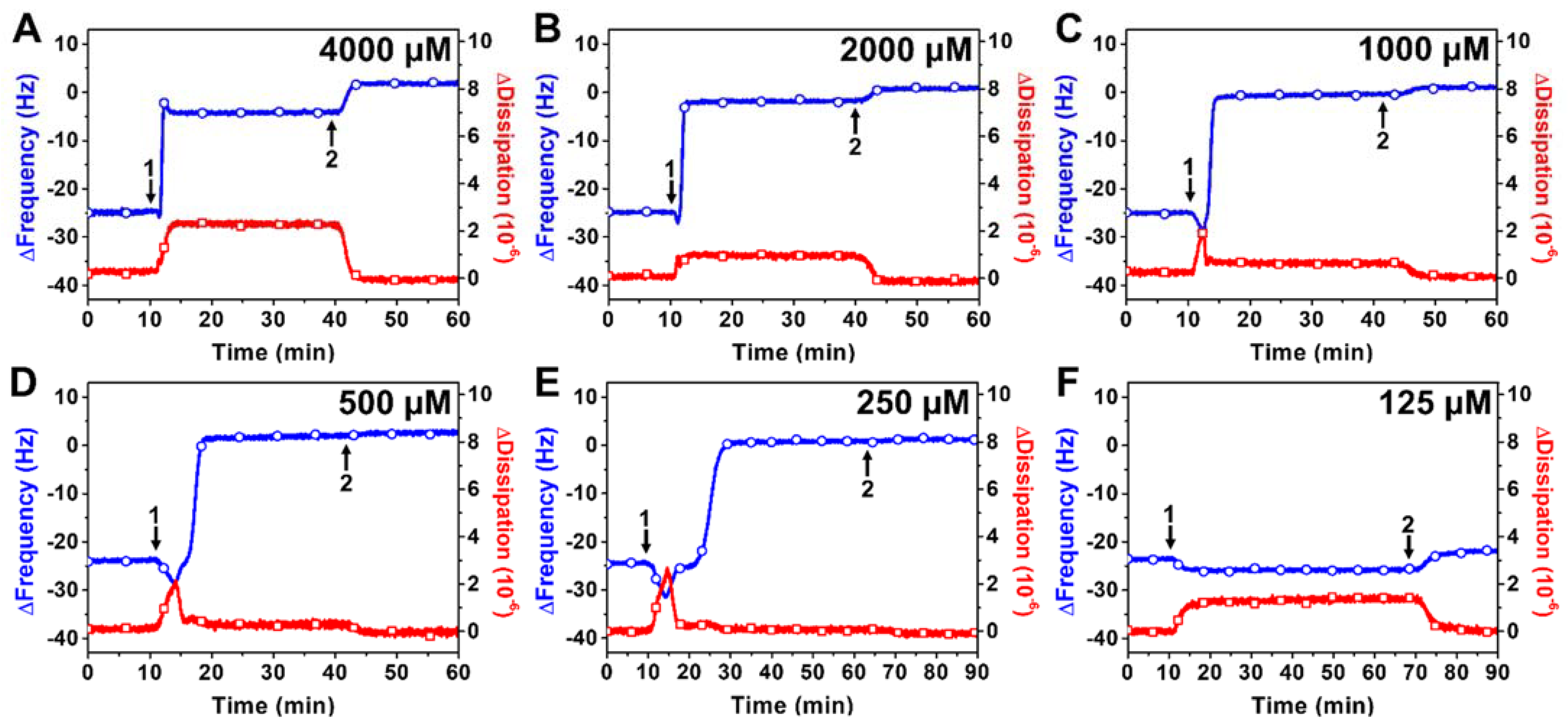
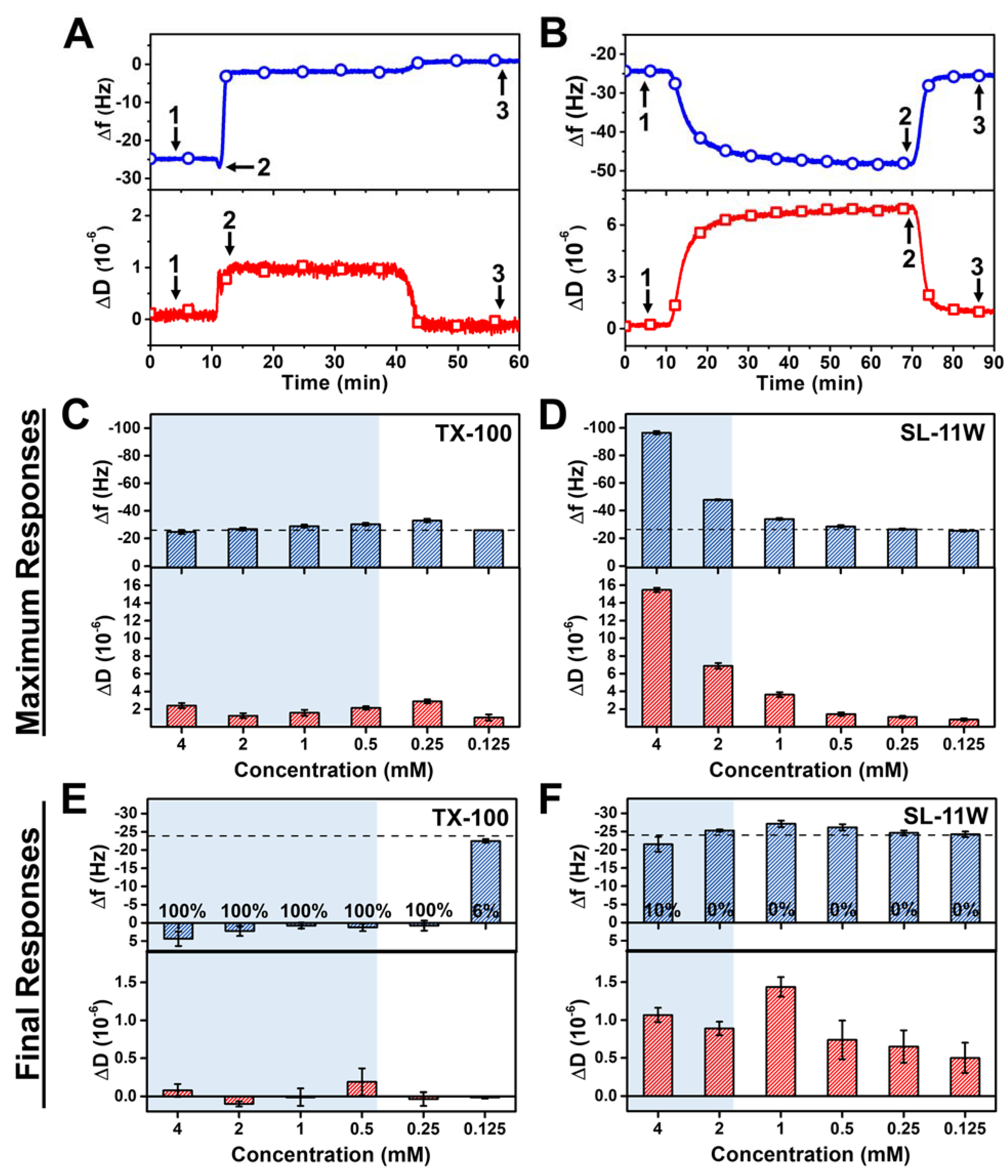
2.2.1. TX-100
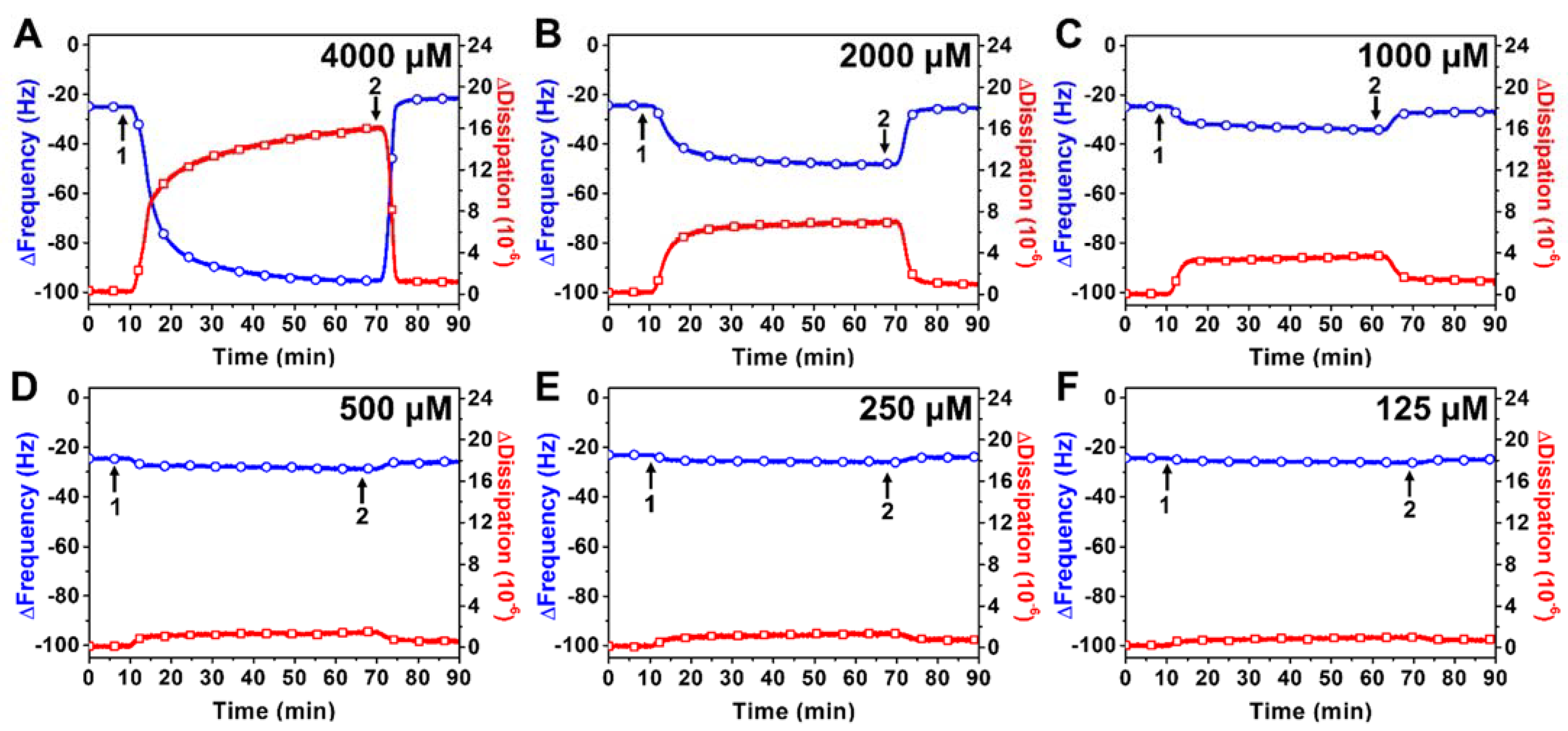
2.2.2. SL-11W
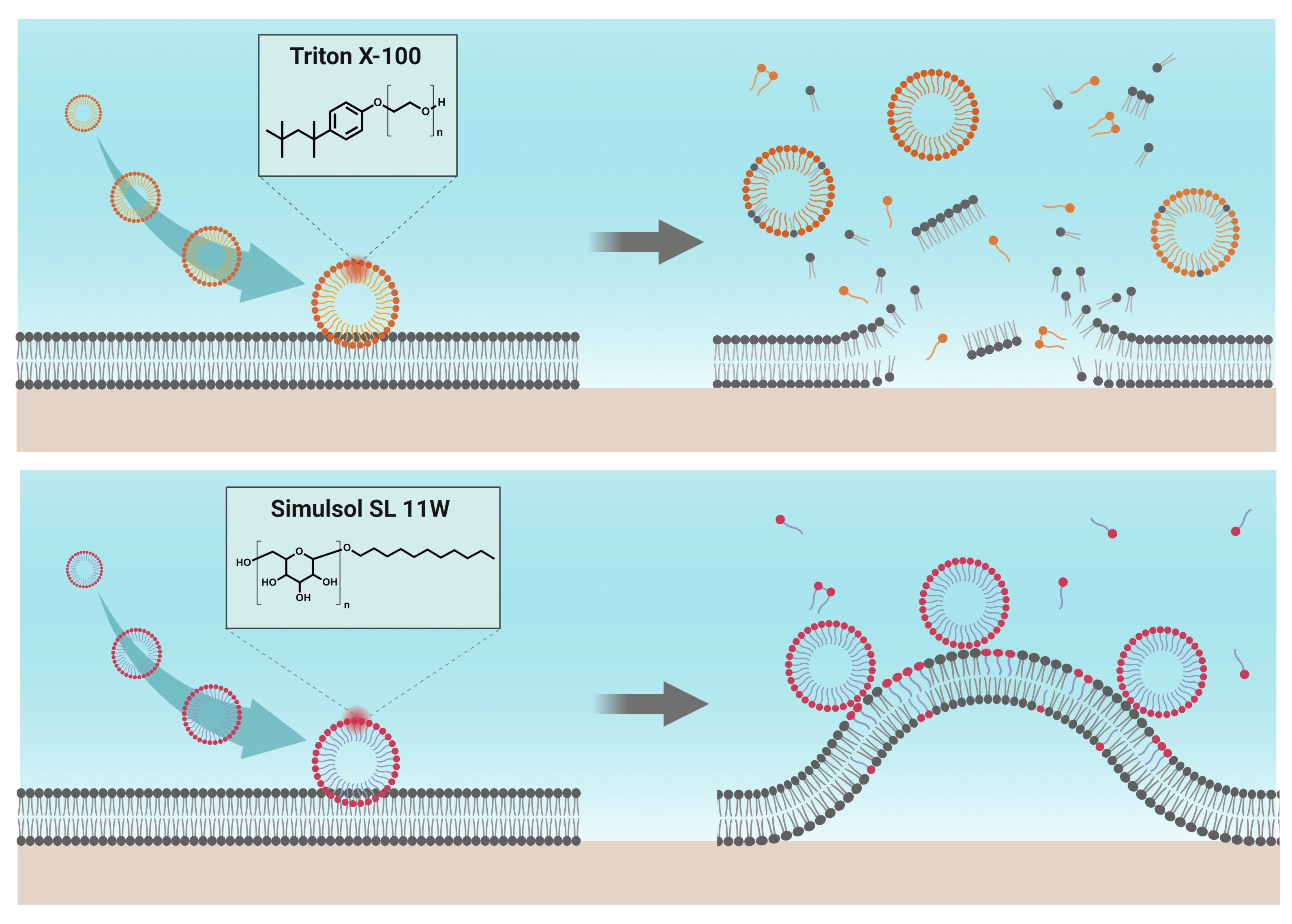
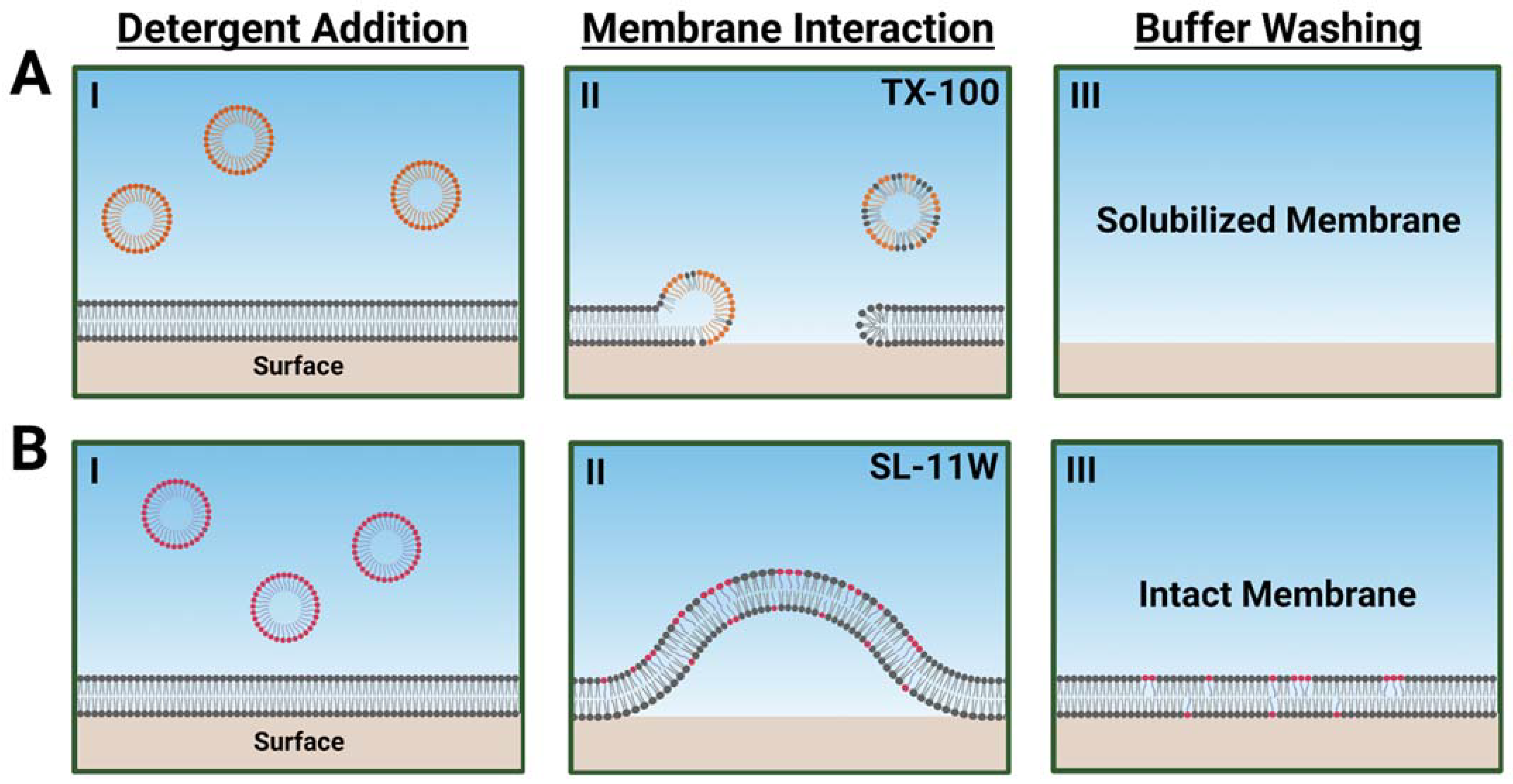
2.3. Comparison of TX-100 and SL-11W Membrane-Disruptive Effects
3. Conclusions
4. Materials and Methods
4.1. Reagents
4.2. Sample Preparation
4.3. Critical Micelle Concentration (CMC) Assay
4.4. Quartz Crystal Microbalance-Dissipation (QCM-D) Measurements
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aranha, H. Current issues in assuring virological safety of biopharmaceuticals. BioProcess Int. 2012, 10, 12–16. [Google Scholar]
- Gilleskie, G.; Rutter, C.; McCuen, B. Biopharmaceutical Manufacturing: Principles, Processes, and Practices; Walter de Gruyter GmbH & Co. KG: Berlin, Germany, 2021. [Google Scholar]
- Cameron, R.; Smith, K. Virus clearance methods applied in bioprocessing operations: An overview of selected inactivation and removal methods. Pharm. Bioprocess. 2014, 2, 75–83. [Google Scholar] [CrossRef]
- Shukla, A.; Aranha, H. Pharmaceutical viral clearance for biopharmaceutical downstream processes. Pharm. Bioprocess. 2015, 3, 127–138. [Google Scholar] [CrossRef]
- Remington, K. Fundamental strategies for viral clearance. Part 1: Exploring the regulatory implications. BioProcess Int. 2015, 13, 10–16. [Google Scholar]
- White, R.; Jobling, S.; Hoare, S.; Sumpter, J.; Parker, M. Environmentally persistent alkylphenolic compounds are estrogenic. Endocrinology 1994, 135, 175–182. [Google Scholar] [CrossRef] [PubMed]
- ECHA 12 New Substances Added to the Authorisation List. Available online: https://echa.europa.eu/en/-/reach-authorisation-list-updated (accessed on 10 December 2021).
- ECHA. SVHC Support Document for Identification of 4-(1,1,3,3,-Tetramethybutyl) Phenol, Ethoxylated. 2012. Available online: https://echa.europa.eu/documents/10162/430c2613-588f-8b08-8a72-df4013727ef8 (accessed on 10 December 2021).
- Conley, L.; Tao, Y.; Henry, A.; Koepf, E.; Cecchini, D.; Pieracci, J.; Ghose, S. Evaluation of eco-friendly zwitterionic detergents for enveloped virus inactivation. Biotechnol. Bioeng. 2017, 114, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Farcet, J.B.; Kindermann, J.; Karbiener, M.; Kreil, T.R. Development of a Triton X-100 replacement for effective virus inactivation in biotechnology processes. Eng. Rep. 2019, 1, e12078. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Luo, W.; Hoffman, J.; Huang, L.; Sandefur, S.; Hall, T.; Murphy, M.; O’Donnell, S. Insights into virus inactivation by polysorbate 80 in the absence of solvent. Biotechnol. Prog. 2020, 36, e2953. [Google Scholar] [CrossRef]
- Luo, W.; Hickman, D.; Keykhosravani, M.; Wilson, J.; Fink, J.; Huang, L.; Chen, D.; O’Donnell, S. Identification and characterization of a Triton X-100 replacement for virus inactivation. Biotechnol. Prog. 2020, 36, e3036. [Google Scholar] [CrossRef] [PubMed]
- Farcet, J.B.; Kindermann, J.; Karbiener, M.; Scheinecker, R.; Kostner, O.; Kreil, T.R. Synthesis of “Nereid”, a new phenol-free detergent to replace Triton X-100 in virus inactivation. J. Med. Virol. 2021, 93, 3880–3889. [Google Scholar] [CrossRef]
- Feroz, H.; Cetnar, D.; Hewlett, R.; Sharma, S.; Holstein, M.; Ghose, S.; Li, Z.J. Surrogate model to screen for inactivation-based clearance of enveloped viruses during biotherapeutics process development. Biotechnol. J. 2021, 16, 2100176. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.; Horzinek, M. Resistance of berne virus to physical and chemical treatment. Vet. Microbiol. 1986, 11, 41–49. [Google Scholar] [CrossRef]
- Ukkonen, P.; Korpela, J.; Suni, J.; Hedman, K. Inactivation of human immunodeficiency virus in serum specimens as a safety measure for diagnostic immunoassays. Eur. J. Clin. Microbiol. Infect. Dis. 1988, 7, 518–523. [Google Scholar] [CrossRef] [PubMed]
- Laethem, V.; Wijngaerden, V.; Ranst, V.; Clercq, D. Long-term stability of human immunodeficiency virus viral load and infectivity in whole blood. Eur. J. Clin. Investig. 1999, 29, 445–452. [Google Scholar] [CrossRef]
- Colavita, F.; Quartu, S.; Lalle, E.; Bordi, L.; Lapa, D.; Meschi, S.; Vulcano, A.; Toffoletti, A.; Bordi, E.; Paglia, M.G. Evaluation of the inactivation effect of Triton X-100 on Ebola virus infectivity. J. Clin. Virol. 2017, 86, 27–30. [Google Scholar] [CrossRef] [PubMed]
- López, O.; de la Maza, A.; Coderch, L.; López-Iglesias, C.; Wehrli, E.; Parra, J.L. Direct formation of mixed micelles in the solubilization of phospholipid liposomes by Triton X-100. FEBS Lett. 1998, 426, 314–318. [Google Scholar] [CrossRef] [Green Version]
- Pizzirusso, A.; De Nicola, A.; Sevink, G.A.; Correa, A.; Cascella, M.; Kawakatsu, T.; Rocco, M.; Zhao, Y.; Celino, M.; Milano, G. Biomembrane solubilization mechanism by Triton X-100: A computational study of the three stage model. Phys. Chem. Chem. Phys. 2017, 19, 29780–29794. [Google Scholar] [CrossRef]
- Yoon, B.K.; Jackman, J.A.; Kim, M.C.; Cho, N.-J. Spectrum of membrane morphological responses to antibacterial fatty acids and related surfactants. Langmuir 2015, 31, 10223–10232. [Google Scholar] [CrossRef]
- Yoon, B.K.; Jackman, J.A.; Valle-González, E.R.; Cho, N.-J. Antibacterial free fatty acids and monoglycerides: Biological activities, experimental testing, and therapeutic applications. Int. J. Mol. Sci. 2018, 19, 1114. [Google Scholar] [CrossRef] [Green Version]
- Jackman, J.A.; Hakobyan, A.; Zakaryan, H.; Elrod, C.C. Inhibition of African Swine Fever virus in liquid and feed by medium-chain fatty acids and glycerol monolaurate. J. Anim. Sci. Biotechnol. 2020, 11, 114. [Google Scholar] [CrossRef]
- Yoon, B.K.; Jeon, W.-Y.; Sut, T.N.; Cho, N.-J.; Jackman, J.A. Stopping membrane-enveloped viruses with nanotechnology strategies: Toward antiviral drug development and pandemic preparedness. ACS Nano 2020, 15, 125–148. [Google Scholar] [CrossRef]
- Heerklotz, H.; Seelig, J. Correlation of membrane/water partition coefficients of detergents with the critical micelle concentration. Biophys. J. 2000, 78, 2435–2440. [Google Scholar] [CrossRef] [Green Version]
- Henriksen, J.R.; Andresen, T.L.; Feldborg, L.N.; Duelund, L.; Ipsen, J.H. Understanding detergent effects on lipid membranes: A model study of lysolipids. Biophys. J. 2010, 98, 2199–2205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goddard, E.; Turro, N.; Kuo, P.; Ananthapadmanabhan, K. Fluorescence probes for critical micelle concentration determination. Langmuir 1985, 1, 352–355. [Google Scholar] [CrossRef] [PubMed]
- Nemethy, G.; Ray, A. Difference spectrophotometric method for the determination of critical micelle concentrations. J. Phys. Chem. 1971, 75, 804–808. [Google Scholar] [CrossRef]
- Haque, M.E.; Das, A.R.; Moulik, S.P. Behaviors of sodium deoxycholate (NaDC) and polyoxyethylene tert-octylphenyl ether (Triton X-100) at the air/water interface and in the bulk. J. Phys. Chem. 1995, 99, 14032–14038. [Google Scholar] [CrossRef]
- Cho, N.-J.; Frank, C.W.; Kasemo, B.; Höök, F. Quartz crystal microbalance with dissipation monitoring of supported lipid bilayers on various substrates. Nat. Protoc. 2010, 5, 1096–1106. [Google Scholar] [CrossRef]
- Kolahdouzan, K.; Jackman, J.A.; Yoon, B.K.; Kim, M.C.; Johal, M.S.; Cho, N.-J. Optimizing the formation of supported lipid bilayers from bicellar mixtures. Langmuir 2017, 33, 5052–5064. [Google Scholar] [CrossRef] [PubMed]
- Jackman, J.A.; Cho, N.-J. Supported lipid bilayer formation: Beyond vesicle fusion. Langmuir 2020, 36, 1387–1400. [Google Scholar] [CrossRef]
- Keller, C.; Kasemo, B. Surface specific kinetics of lipid vesicle adsorption measured with a quartz crystal microbalance. Biophys. J. 1998, 75, 1397–1402. [Google Scholar] [CrossRef] [Green Version]
- Yoon, B.K.; Jackman, J.A.; Kim, M.C.; Sut, T.N.; Cho, N.-J. Correlating membrane morphological responses with micellar aggregation behavior of capric acid and monocaprin. Langmuir 2017, 33, 2750–2759. [Google Scholar] [CrossRef] [PubMed]
- Yoon, B.K.; Park, S.; Ma, G.J.; Kolahdouzan, K.; Zhdanov, V.P.; Jackman, J.A.; Cho, N.-J. Competing interactions of fatty acids and monoglycerides trigger synergistic phospholipid membrane remodeling. J. Phys. Chem. Lett. 2020, 11, 4951–4957. [Google Scholar] [CrossRef] [PubMed]
- Yoon, B.K.; Park, S.; Jackman, J.A.; Cho, N.-J. Supported lipid bilayer platform for characterizing the optimization of mixed monoglyceride nano-micelles. Appl. Mater. Today 2020, 19, 100598. [Google Scholar] [CrossRef]
- Fatisson, J.; Azari, F.; Tufenkji, N. Real-time QCM-D monitoring of cellular responses to different cytomorphic agents. Biosens. Bioelectron. 2011, 26, 3207–3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serro, A.; Carapeto, A.; Paiva, G.; Farinha, J.; Colaço, R.; Saramago, B. Formation of an intact liposome layer adsorbed on oxidized gold confirmed by three complementary techniques: QCM-D, AFM and confocal fluorescence microscopy. Surf. Interface Anal. 2012, 44, 426–433. [Google Scholar] [CrossRef]
- Dalgarno, P.A.; Juan-Colás, J.; Hedley, G.J.; Piñeiro, L.; Novo, M.; Perez-Gonzalez, C.; Samuel, I.D.; Leake, M.C.; Johnson, S.; Al-Soufi, W. Unveiling the multi-step solubilization mechanism of sub-micron size vesicles by detergents. Sci. Rep. 2019, 9, 12897. [Google Scholar] [CrossRef] [Green Version]
- Fujino, Y.; Nakamura, R.; Han, H.-W.; Yamashita, I.; Shimizu, T.; Shingubara, S.; Ito, T. Electrochemical impedance spectroscopy study of liposome adsorption and rupture on self-assembled monolayer: Effect of surface charge. J. Electroanal. Chem. 2020, 878, 114572. [Google Scholar] [CrossRef]
- Asai, N.; Matsumoto, N.; Yamashita, I.; Shimizu, T.; Shingubara, S.; Ito, T. Detailed analysis of liposome adsorption and its rupture on the liquid-solid interface monitored by LSPR and QCM-D integrated sensor. Sens. Bio-Sens. Res. 2021, 32, 100415. [Google Scholar] [CrossRef]
- Sauerbrey, G. Verwendung von schwingquarzen zur wägung dünner schichten und zur mikrowägung. Z. Phys. 1959, 155, 206–222. [Google Scholar] [CrossRef]
- Tang, T.; Savva, A.; Traberg, W.C.; Xu, C.; Thiburce, Q.; Liu, H.-Y.; Pappa, A.-M.; Martinelli, E.; Withers, A.; Cornelius, M.; et al. Functional infectious nanoparticle detector: Finding viruses by detecting their host entry functions using organic bioelectronic devices. ACS Nano 2021, 15, 18142–18152. [Google Scholar] [CrossRef]
- Liu, H.-Y.; Pappa, A.-M.; Pavia, A.; Pitsalidis, C.; Thiburce, Q.; Salleo, A.; Owens, R.M.; Daniel, S.J. Self-assembly of mammalian-cell membranes on bioelectronic devices with functional transmembrane proteins. Langmuir 2020, 36, 7325–7331. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gooran, N.; Yoon, B.K.; Jackman, J.A. Supported Lipid Bilayer Platform for Characterizing the Membrane-Disruptive Behaviors of Triton X-100 and Potential Detergent Replacements. Int. J. Mol. Sci. 2022, 23, 869. https://doi.org/10.3390/ijms23020869
Gooran N, Yoon BK, Jackman JA. Supported Lipid Bilayer Platform for Characterizing the Membrane-Disruptive Behaviors of Triton X-100 and Potential Detergent Replacements. International Journal of Molecular Sciences. 2022; 23(2):869. https://doi.org/10.3390/ijms23020869
Chicago/Turabian StyleGooran, Negin, Bo Kyeong Yoon, and Joshua A. Jackman. 2022. "Supported Lipid Bilayer Platform for Characterizing the Membrane-Disruptive Behaviors of Triton X-100 and Potential Detergent Replacements" International Journal of Molecular Sciences 23, no. 2: 869. https://doi.org/10.3390/ijms23020869