1. Introduction
Red fluorescent proteins (red FPs, RFPs) are genetically encoded molecular probes that find use in fluorescence microscopy of biological tissues, such as the brain [
1,
2,
3,
4,
5]. Their red-shifted absorption and fluorescence provide structural and dynamic information from deeper layers of tissue, compared to their green counterparts, because red light penetrates better in scattering media. RFPs are particularly attractive for use in two-photon laser microscopy (TPLM) [
6,
7,
8,
9], where two near-infrared photons can effectively excite the RFP chromophore [
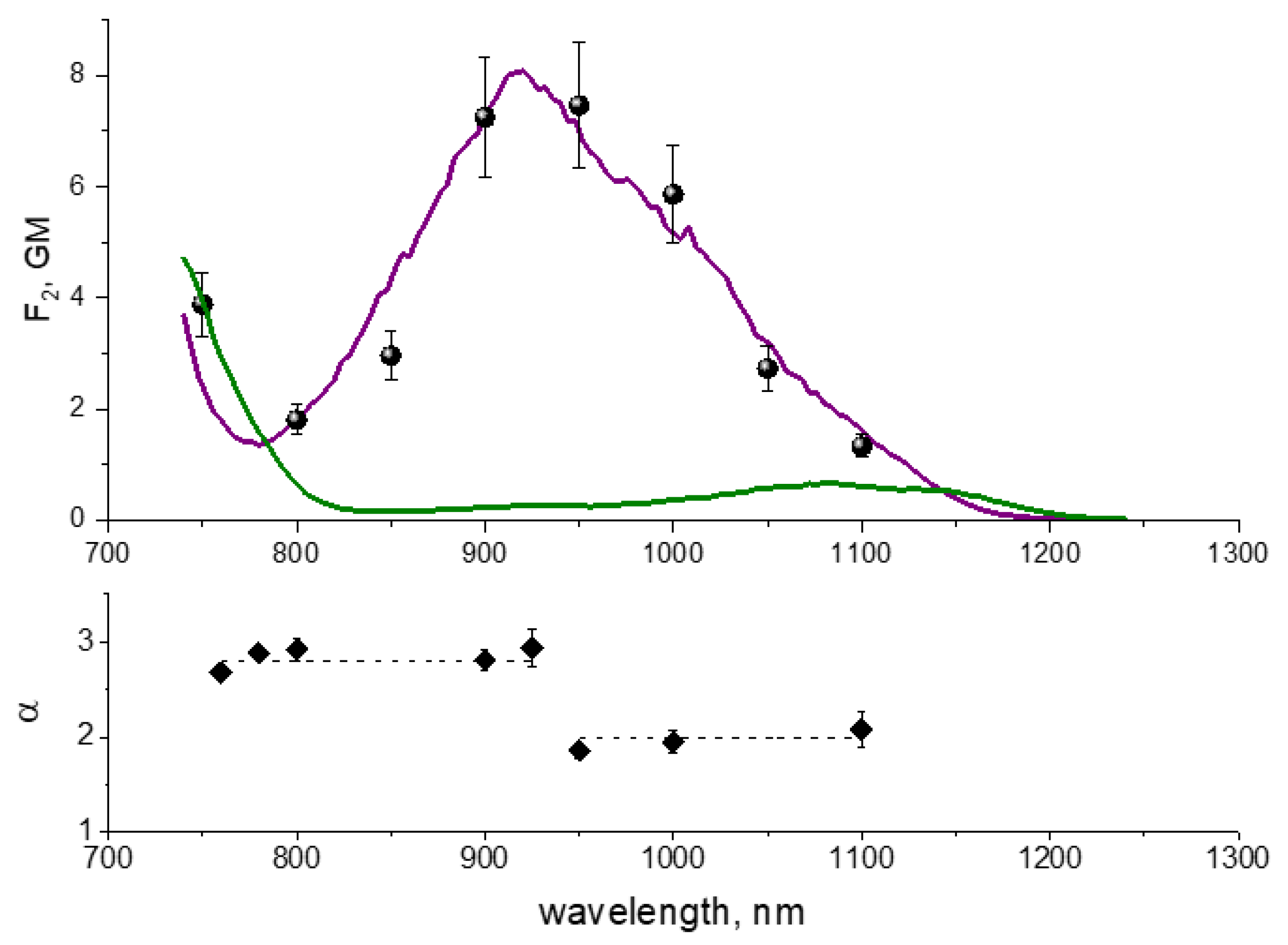
10]. Two-photon excitation (2PE) spectra of RFPs fall in the range from 700 to 1200 nm [
10,
11] that matches the so-called tissue transparency window [
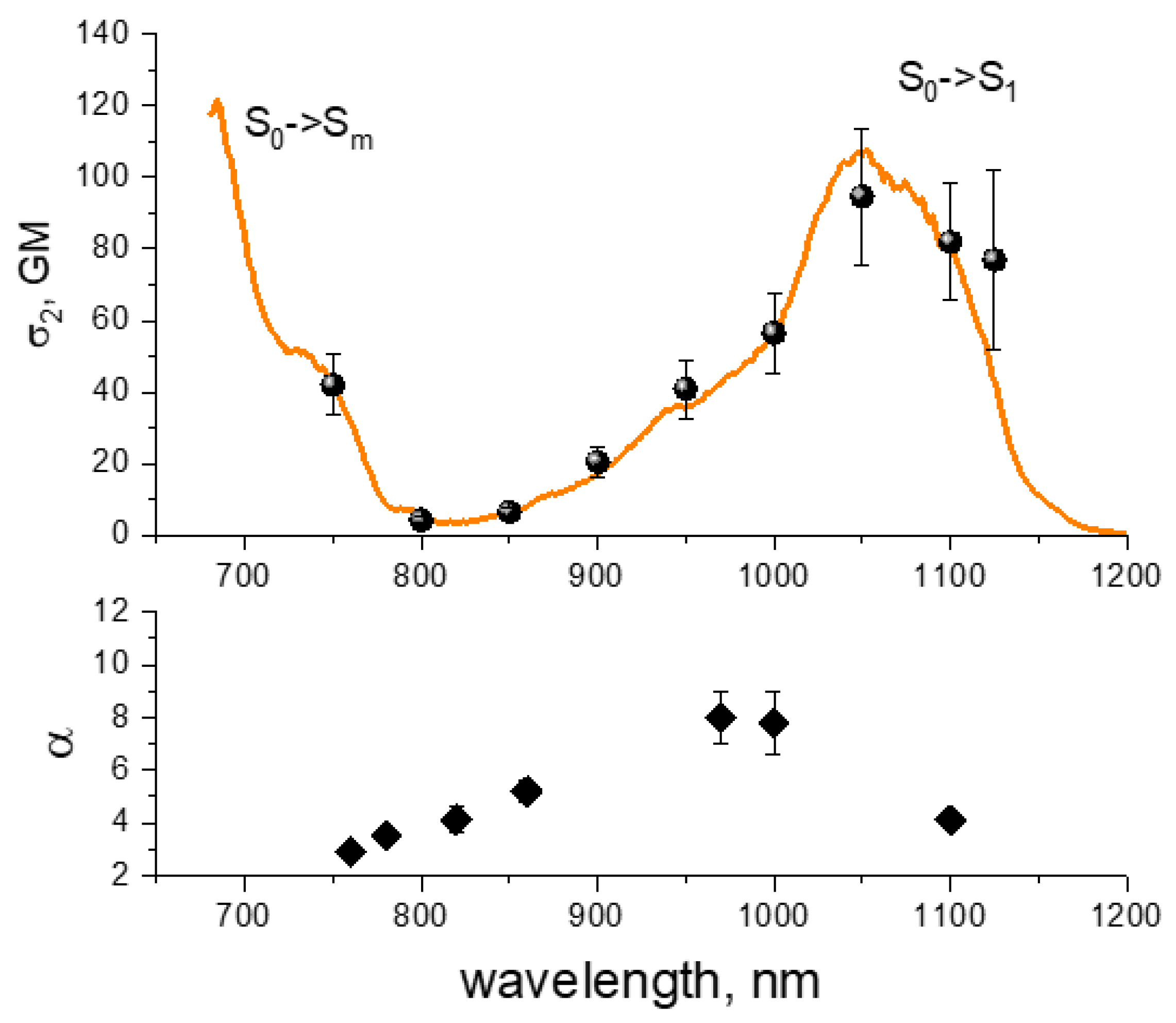
12]. They show two main peaks in this range. The first one, at 1050–1200 nm, typically has moderate two-photon absorption (2PA) cross section,
σ2 = 10–100 GM (1 GM = 10
−50 cm
4 s). The second one, at 700–780 nm, is usually stronger, with
σ2 reaching 100–300 GM in some RFPs. This latter region corresponds to a higher energy, S
0 → S
m, transition that is also present in one-photon absorption spectra as a weak peak near 350–380 nm. When excited with two photons, it gains its intensity due to the nonlinear optical effect of pre-resonance enhancement that occurs when the laser photon energy approaches the lowest S
0 → S
1 transition energy from below, but still does not populate it [
13,
14]. Therefore, excitation at 700–780 nm still corresponds to a simultaneous two-photon absorption with all advantages offered by TPLM, including high spatial (3D) resolution, low out-of-focus damage of tissue, and low autofluorescence background. Given the availability of commercial femtosecond Ti:sapphire lasers, with the high output power and stable operation in this spectral region and the possibility of dual-color two-photon imaging with simultaneous excitation of red and blue FPs [
15], excitation at 700–780 nm provides a very attractive opportunity for TPLM with RFPs. However, researchers quickly discovered that RFPs bleach extremely fast when using femtosecond lasers operating in this wavelength range, with typical pulse-peak photon fluxes of 10
27–10
29 photon/cm
2/s at the sample.
There are a number of reports investigating multiphoton bleaching of fluorescent dyes and FPs, but the molecular mechanisms remain almost completely unexplored. In their seminal work, Patterson and Piston reported on multiphoton bleaching of fluorescein dextran, aminocoumarin dextran, NADH, and Indo-1 [
16]. Using a femtosecond 710 nm excitation, they found that although the initial fluorescence intensity increased quadratically with laser power
P, the photobleaching rate
K increased much faster (i.e., according to a power law,
K~Pα with the exponents
α ranging from 3 to 5). This super-quadratic dependence suggested involvement of multiphoton processes with number of photons
n > 2. It is clear that after direct two-photon excitation, that requires very high photon flux, a large organic molecule can readily undergo several stepwise one-photon absorption events, through a ladder of electronic/vibrational excited states, that make a quasi-continuum above the S
1 level. The most general photochemical outcome of this process is multiphoton ionization or electron detachment (for anionic chromophores) [
17]. For example, fluorescein dextran required absorption of a total of three photons (power exponent 3.1 ± 0.1) for photobleaching. Photodetachment energy of fluorescein lies between 5.1 and 6.3 eV (depending on protonation state) [
18]. For the 1.75 eV photon energy (used in the Patterson and Piston experiment) the above numbers translate into the number of photons,
n ~3, thus supporting the photodetachment mechanism. Photobleaching of aminocoumarin requires five photons (power exponent 5.1 ± 0.2 at 710 nm) [
16]. Given photoionization energy of aminocoumarin of 8.05 eV [
19], a minimum of five photons are required to ionize this molecule, which supports our assumption that the photobleaching is caused by multiphoton ionization.
Marchant and co-authors [
20] demonstrated that the wild-type red FP, DsRed, bleaches in CHO cells according to a third power law (
α = 2.98 ± 0.10) when excited at 760 nm, in the range of average power densities 5–40 μW/μm
2 (approximately corresponding to peak photon fluxes of
I0 = 3 × 10
26–2.5 × 10
27 photon/cm
2/s). This suggests a three-photon initiated process. The authors also found that, upon excitation at 950 nm, DsRed bleaches much slower than at 750 nm excitation, with the laser power adjusted to produce similar initial fluorescence signals at these two wavelengths. Later, Robinson and Marchant [
21] extended this study to several DsRed mutants expressed in HEK-293 cells, and have shown that all of them bleach much faster at 750 nm compared to 950 nm. The power exponents observed for bleaching at 750 mn varied from 2.69 (for DsRed2) to 3.05 (for DsRed [N42Q]). A similar trend of slowing down the bleaching rate upon shifting excitation of DsRed2 to longer wavelengths, from 760 to 880 and further to 1100 nm, was found in [
22]. Higher than quadratic power dependence (
α = 2.9 ± 0.10) was also observed in another DsRed mutant, tdRFP, expressed in T cells and excited at 1100 nm with
I0 = (2–3.5) × 10
29 photon/cm
2/s [
23]. Interestingly, the same excitation fluxes resulted in much faster bleaching of enhanced green fluorescent protein (EGFP) at 850 nm, with
α = 2.57 ± 0.25. Other groups also reported a super-quadratic dependence of photobleaching for EGFP and cyan FP (CFP) [
24,
25,
26,
27]. Kalies et al. correlated the power exponent of the EGFP bleaching rate with the multiphoton electron photodetachment from the chromophore or a nearby tryptophan residue [
26].
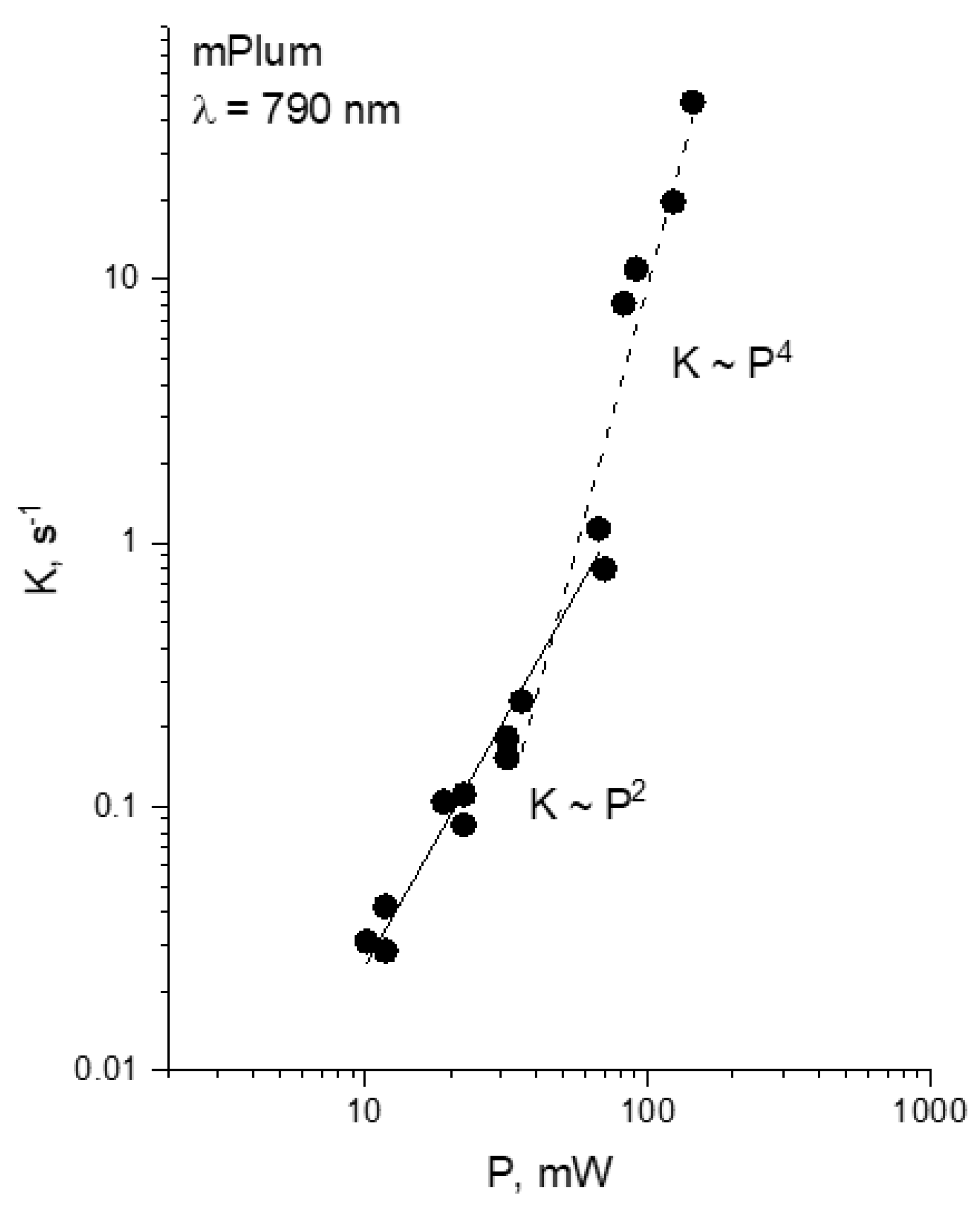
Drobizhev and co-authors characterized multiphoton bleaching of monomeric RFPs, mFruits (mCherry, mPlum, and mStrawberry) and DsRed2 in buffer solution and
E.
coli cells, with the emphasis on possible photochemical mechanisms [
28]. The rate of the first step of a chain of photochemical reactions leading to photobleaching was measured as a function of laser intensity at 790 nm. The DsRed2 rate followed a power law with
α = 2.84 ± 0.01, at
I0 = (0.3–1.1) × 10
29 photon/cm
2/s, in agreement with previous observations [
20,
21]. In DsRed2, photobleaching occurs through the three-photon absorption process with a slight saturation of the one-photon S
m → S
n transition. Vertical detachment energy (VDE) of the anionic chromophore inside DsRed2 lies at ~7.24 eV [
28], suggesting that at least five photons (of 790 nm) are needed to reach this threshold. A three-photon-induced electron transfer reaction starting from the bound S
n state of DsRed2 is probably a first step of phototransformation [
28]. mFruits bleach according to a power law with
α = 3.2–3.5, suggesting a four-photon mechanism with strong saturation of one or two one-photon transitions, following initial two-photon absorption step. Electron photodetachment energies of mFruits are found in the range 5.52 (mCherry)–5.61 eV (mPlum) [
28]. Absorption of four photons (at 790 nm) can easily provide enough energy to overcome this threshold and, therefore, the four-photon photodetachment is suggested as a first step of photobleaching in mFruits [
28]. The chromophore environment of DsRed2 contains more positive amino acid residues compared to mFruits, and this can be a reason of its higher photodetachment energy.
Since the fluorescence signal increases slower (quadratically), with the laser peak intensity compared to photobleaching rate, any reduction of the laser pulse-peak intensity should be beneficial until the moment where fluorescence signal becomes undetectable against the noise. Following this idea, several approaches to reduce photobleaching were suggested. Drummond et al. proposed to (1) block the laser with the programmed Pockels cell during the “flyback” time (when fluorescence is not collected but the laser still irradiates the sample); and (2) increase the sensitivity of detection by using an additional widefield detector in the transmission position, which made it possible to use less excitation power [
25]. The use of passive pulse splitters, providing less peak intensity but higher repetition rate, resulted in faster reduction of photobleaching relatively to fluorescence signal [
29]. On the other hand, if multiphoton absorption involves triplet manifold, lower pulse repetition rate can be beneficial [
30]. Phase modulation or dispersion compensation of femtosecond pulses also helped reducing multiphoton bleaching [
27,
31,
32,
33]. Although these methods provide some improvement on the fluorescence/bleaching ratio, they require technical upgrades of commercial 2P microscopes.
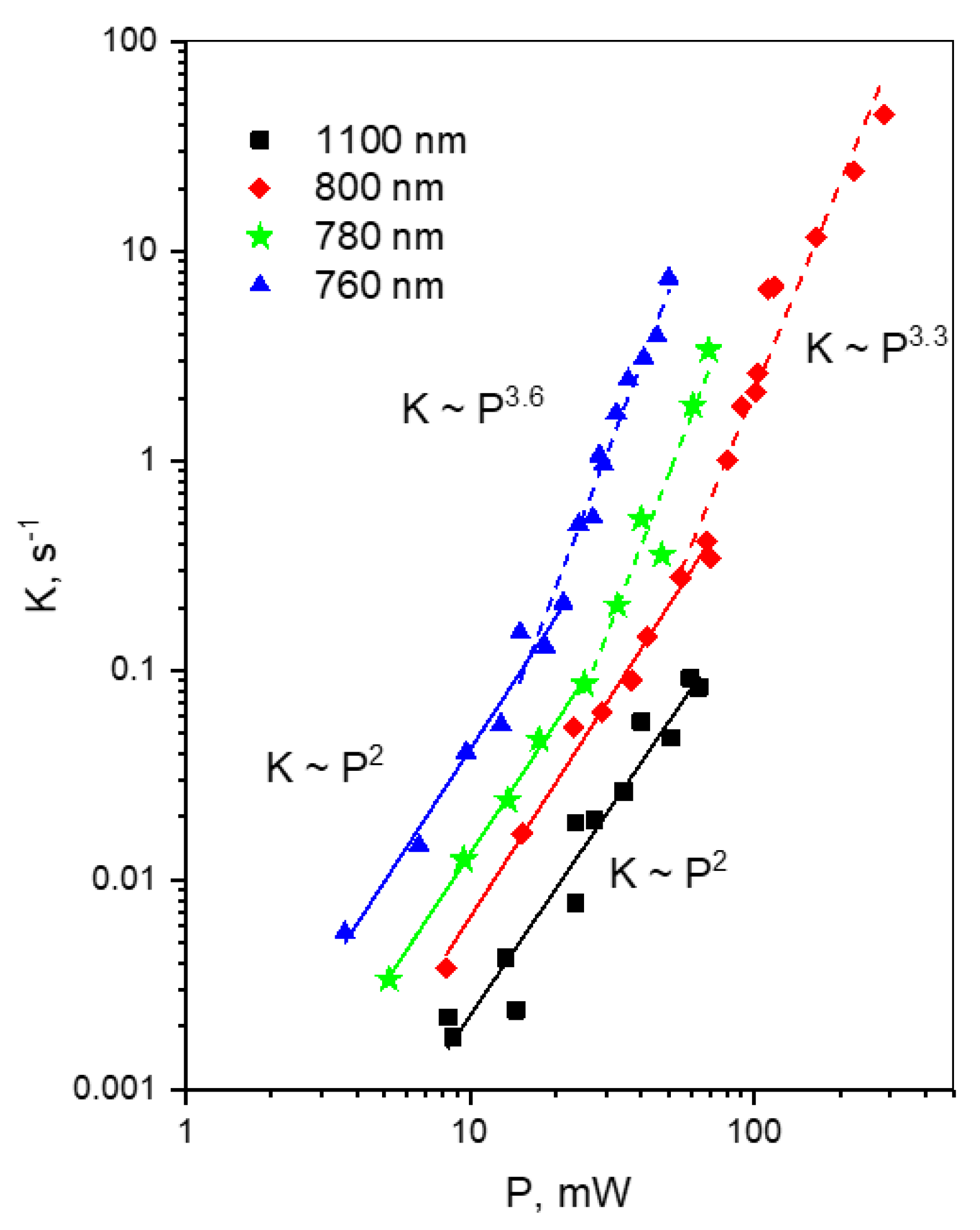
Here we aim at finding simple solutions to the problem of rapid multiphoton bleaching in terms of optimization of laser intensity and excitation wavelength. They require the (phenomenological) knowledge of photochemical mechanisms and their dependence on laser intensity and wavelength. First, one can expect that at small laser intensities the bleaching rate should show a quadratic dependence on intensity, with the rate equal to that observed with one-photon excitation (with the 2P and 1P excitation rates being equal). A transition between quadratic and higher-order dependence defines a particular threshold for laser power, P*. An optimum intensity for TPLM should be chosen close to the P* value because it provides the highest signal-to-background ratio (SBR), and any further reduction of intensity would not improve the fluorescence/bleaching ratio. A particular value of threshold intensity is a function of wavelength because the corresponding laser and molecular parameters depend on wavelength. Second, if the bleaching is due to multiphoton ionization (electron detachment) with n photons involved (like in mFruits), one can look for a threshold photon energy (wavelength) where this process in not allowed energetically: nhν < VDE.
We check these ideas for a series of RFPs, including popular monomeric variants mCherry [
1] and mPlum [
34], one of the two-photon brightest RFP, tdTomato [
1,
10], and a new red Ca
2+ sensor with the long Stokes shift in the Ca
2+-bound state, jREX-GECO1 [
35]. Compared to the previous work [
28], we study photobleaching rates in a broader range of excitation intensities and at different excitation wavelengths across the 2PE spectrum. We found some important threshold values for the laser photon fluxes and wavelengths for these RFPs and show that in some cases one can predict these values if the molecular and laser parameters are known.
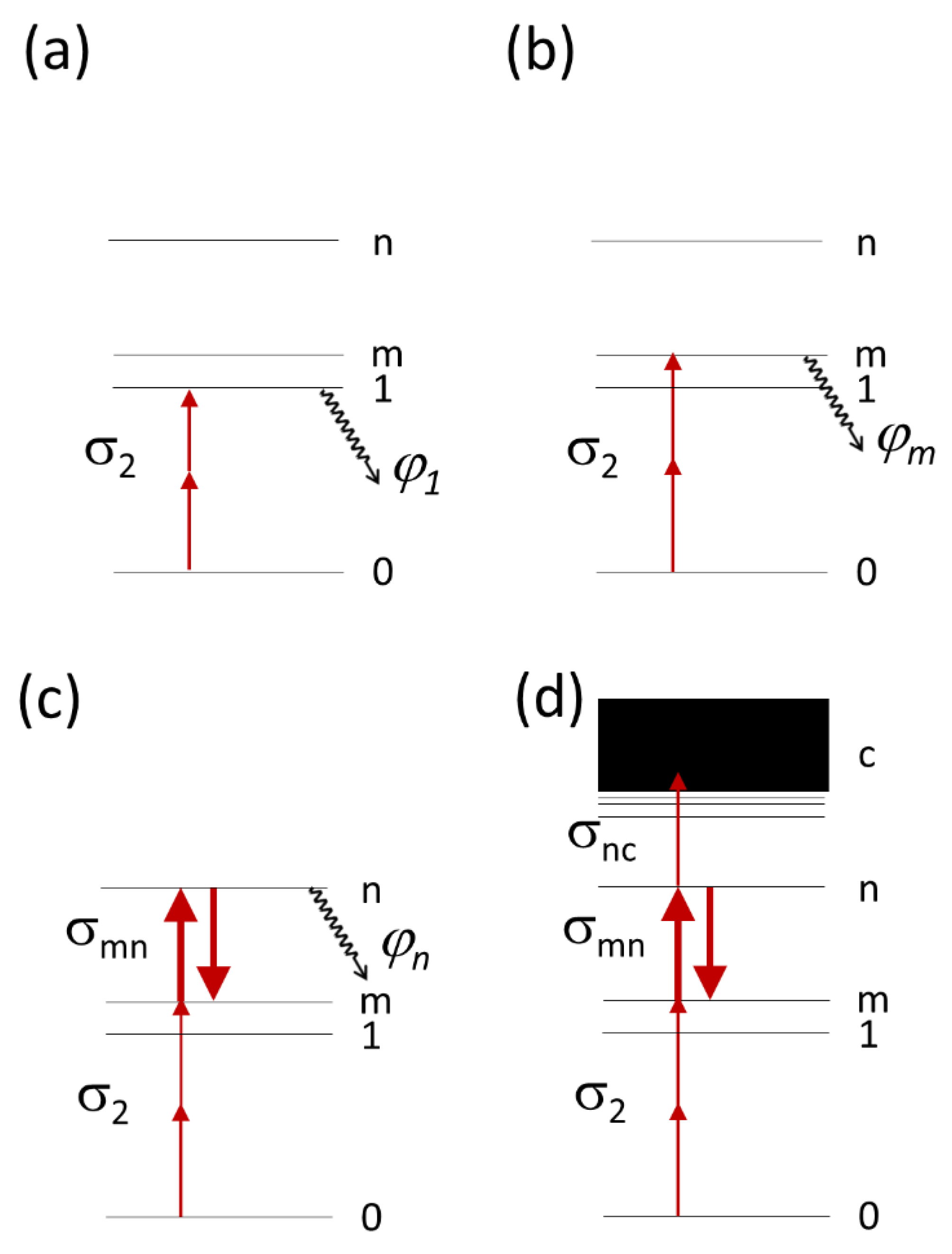
3. Discussion
Our results show that even within a set of red FPs with the same chemical structure of the chromophore, the mechanisms of multiphoton bleaching are quite different. This includes different numbers of photons involved in the process and different power dependences observed at various excitation wavelengths even for a particular protein. Nevertheless, we try to compare here the performance of one protein at different wavelengths or different proteins at similar wavelength using a specific “figure of merit” (FOM) that reflects the maximum possible number of fluorescence photons collected from an FP during a given pixel dwell time in a TPLM experiment. The FOM parameter involves the molecular fluorescence brightness and photobleaching parameters (see
Supplementary Information for derivation of equations). In the case of quadratic power dependence of photobleaching rate, the FOM is equal to the ratio of fluorescence quantum yield and photochemical reaction quantum yield starting either from state 1 or m (see
Figure 1a,b):
For the third-order process without saturation,
Figure 1c, the FOM reads
In the fourth-order process without saturation, we have
Using these equations, we estimated the FOM values of the fourth-order process observed experimentally for mCherry, mPlum, and tdTomato at several wavelengths, see
Table 5. It should be kept in mind that the comparison between different wavelengths and different proteins using these numbers is possible only for the same laser peak photon flux and the same mechanism of the process.
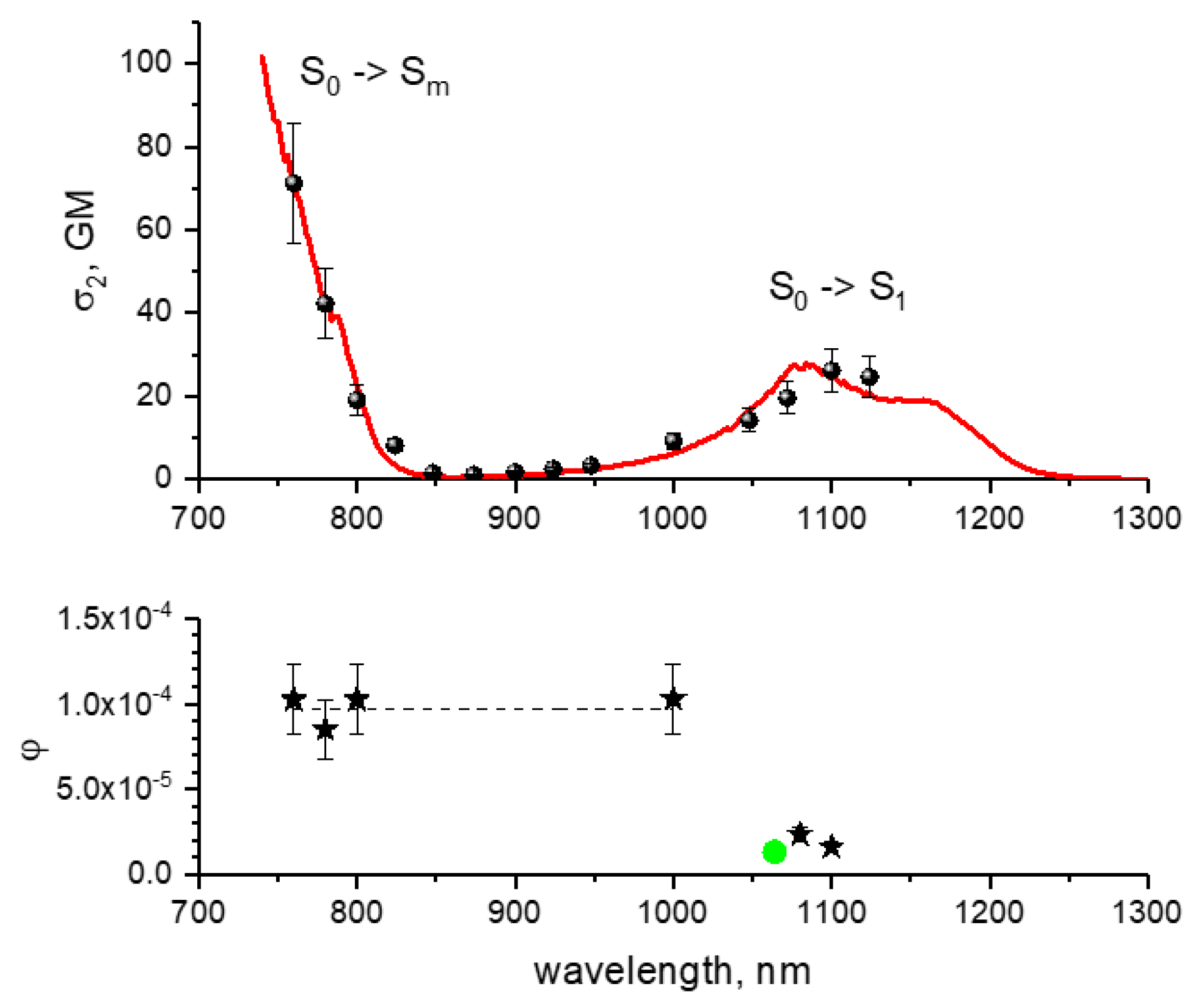
For the S0 → Sm transition, excitation of mCherry at 800 nm is slightly beneficial, compared to 780 nm, because of smaller σmnσnc parameter for the former. At 790 nm, mPlum shows about two-times smaller FOM (4) compared to mCherry because of proportionally smaller fluorescence quantum yield. tdTomato has the FOM (4) similar to mCherry at 780–820 nm, but much larger at 1100 nm. This strong enhancement is due to much higher 2PA cross section of tdTomato compared to mCherry at this wavelength.
Table 6 presents the FOM
(2) values obtained for the RFPs under study in the power ranges where the bleaching rate follows quadratic power dependence.
In this regime, mCherry has much higher FOM
(2) at 1100 nm than at 760–1000 nm. This is explained by much lower
φ1 for the reaction starting from the S
1 level relatively to S
m level. Compared to mCherry, mPlum shows lower FOM
(2) at 790 nm, and jREX-GECO1—higher at 950–1000 nm. Excitation of tdTomato with low powers at 970–1000 nm (
Figure 9a), provides extremely large relative FOM
(2) values thanks to high fluorescence quantum yield and relatively low photobleaching yield.
In summary, we developed a method of quantitative characterization of multiphoton bleaching of FPs expressed in
E. coli colonies that one can use to compare bleaching rates of different mutants in standardized experimental conditions. Our physical model proposed in [
28] and further developed here, makes it possible to obtain key molecular parameters involved in photobleaching process. Knowing laser parameters, such as wavelength, pulse duration and peak photon flux in focal plane in addition to molecular bleaching parameters, makes it possible to predict the bleaching rate in a specific experiment. In addition to quantitative estimations of FOM provided above, we can further suggest some general, qualitative, approaches to reduce the bleaching rates. We have shown here, for a first time to the best of our knowledge, that there is often a range of laser photon fluxes where the power dependence of bleaching follows quadratic law. After a critical threshold,
P =
P*, is passed, the dependence turns to a much faster, super-quadratic one that was observed in several previous studies. We suggest that in the TPLM experiments the working laser intensity should be selected close to this threshold to provide the best SBR. We also found that, in agreement with previous observations [
20,
21,
22], it is more advantageous (in terms of brightness/photobleaching ratio) to excite RFPs at their longer wavelength transition, S
0 → S
1 (near 1050–1100 nm). In particular, in the case of mCherry, excitation at 1100 nm does not activate a detrimental four-photon electron photodetachment process, and the power dependence follows a quadratic law where any increase in laser intensity provides better SBR ratio, but does not change the brightness/photobleaching rate ratio. At shorter wavelengths, corresponding to a higher, S
0 → S
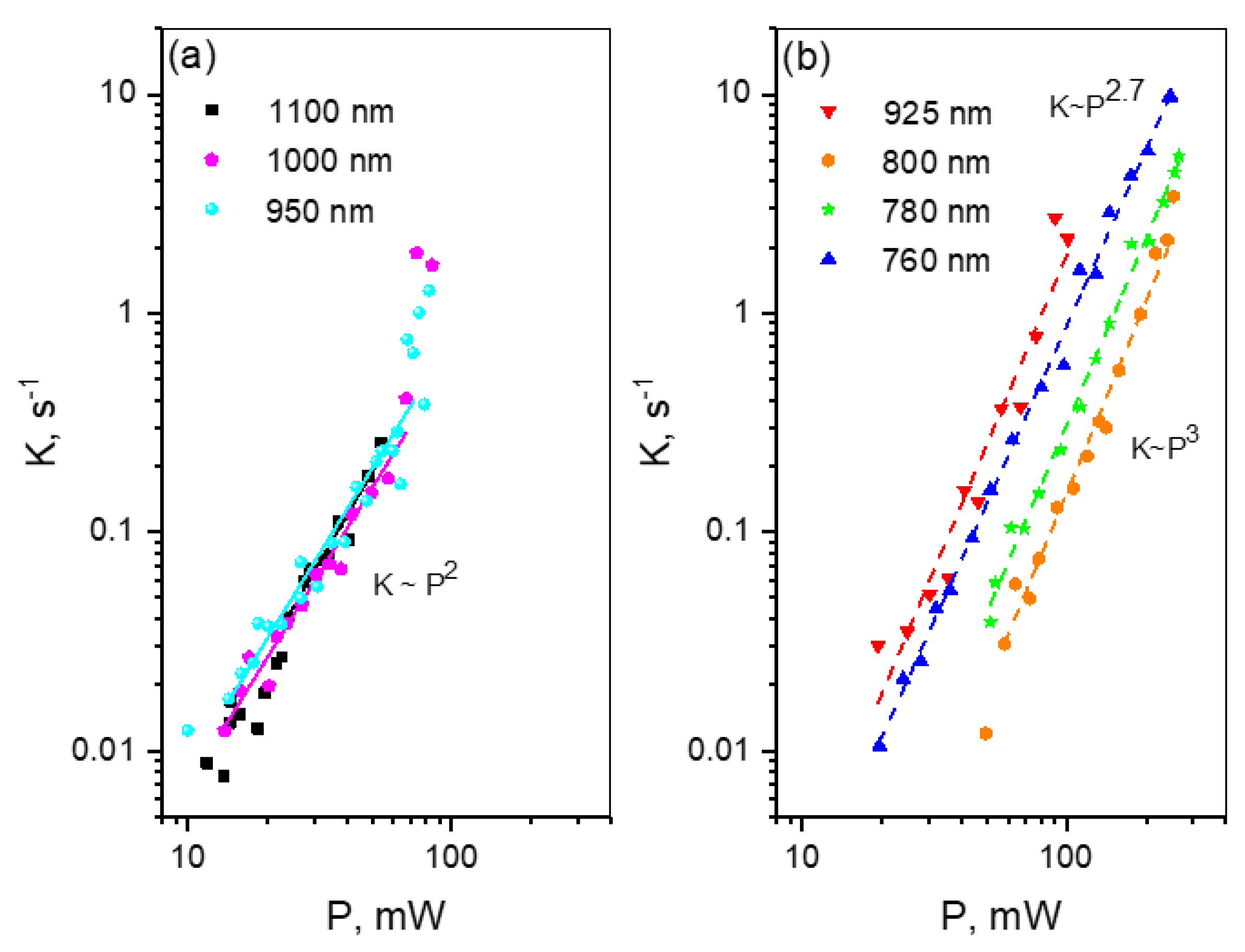
m transition (760–800 nm), there exists a laser intensity threshold, specific for each wavelength, where the power dependence switches from quadratic to much faster law, corresponding to four-photon process. Such switching was also observed for mPlum.
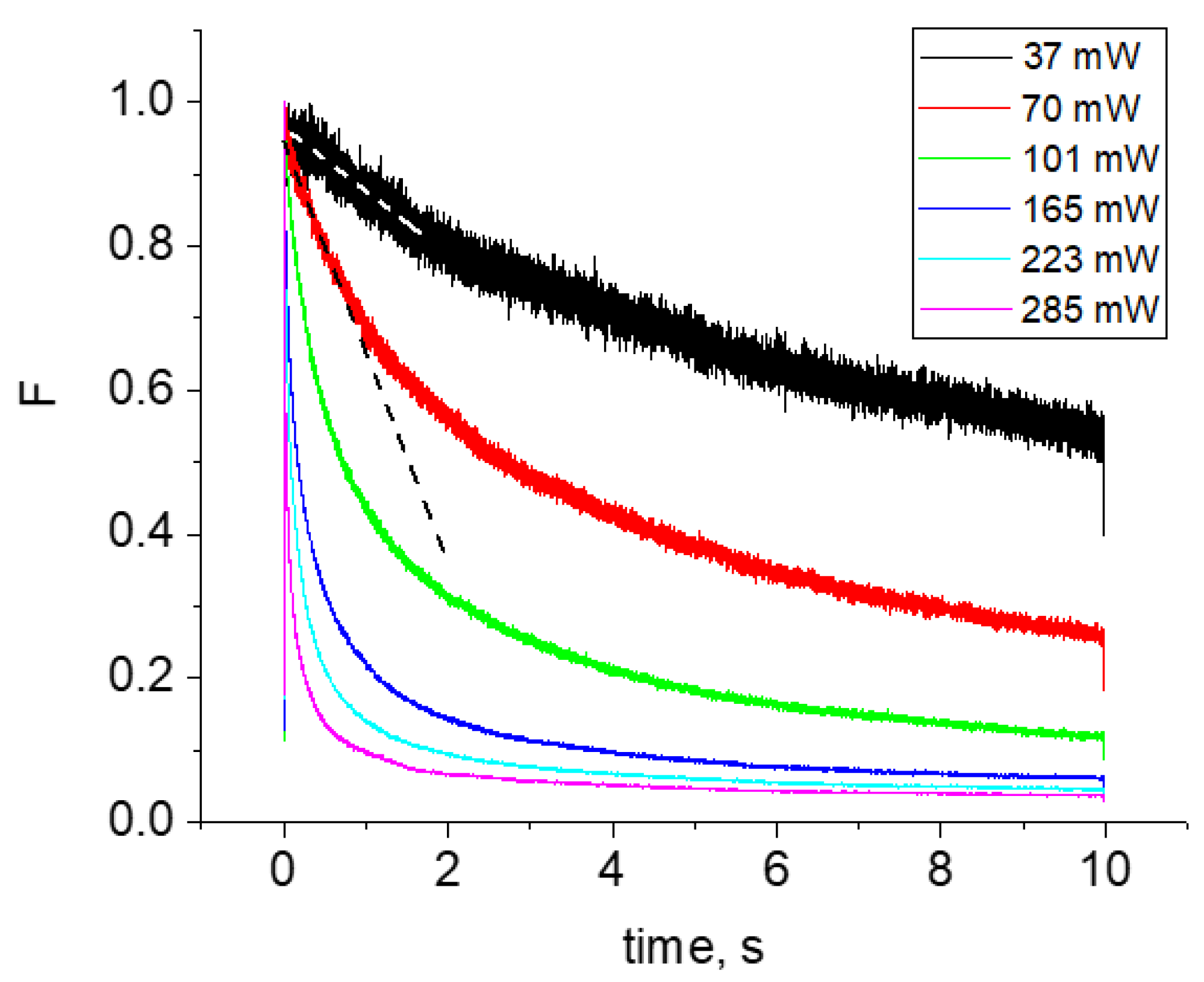
For the new, red genetically-encoded Ca2+-sensor, jREX-GECO1, with a long Stokes shift in the Ca2+-bound state, we observe slow photobleaching, following quadratic power law in a broad spectral region from 950–1100 nm. At shorter wavelengths, λ < 950 nm, the bleaching rate starts to grow faster, i.e., according to a third power law. Therefore, excitation at these wavelengths should be avoided.
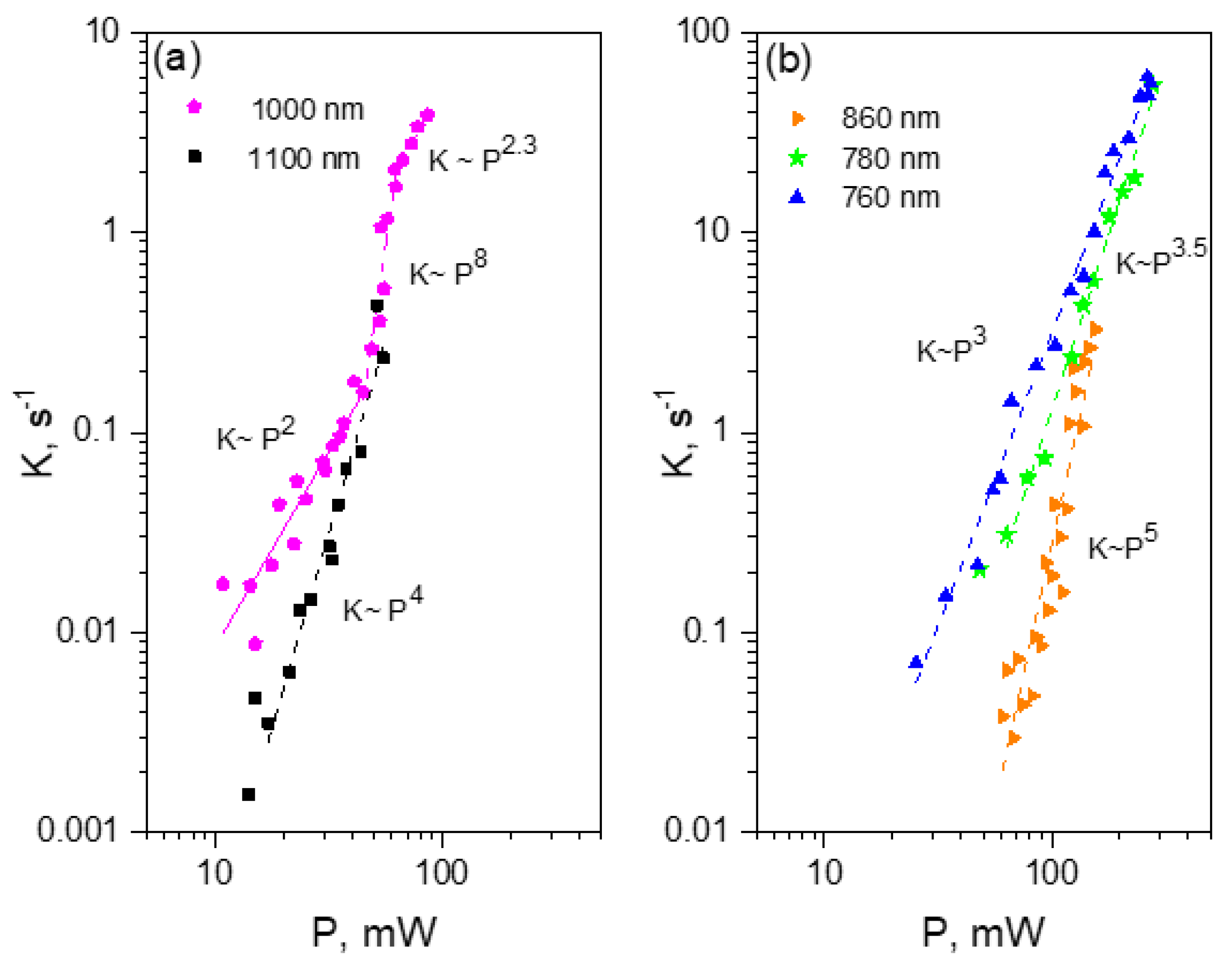
One of the brightest under two-photon excitation red FP, tdTomato, shows unexpected power dependences of the bleaching rate at some particular wavelengths. At the longest wavelength studied, λ = 1100 nm, the bleaching follows a K~P4 dependence, reflecting four-photon process. At 970–1000 nm, the dependence starts as quadratic, and then after passing some critical power, becomes very sharp, following the K~P8 law. At even higher powers, it quickly saturates to become close to quadratic again. At shorter wavelength region, 760–820 nm, the power dependence follows the K~P4 law with some signs of saturation, typical for mCherry and mPlum. We tentatively explain these observations by a presence of two forms of the chromophore in tdTomato, with the neutral form dominating in the region of 950–1000 nm.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}