
Cyclanilide Induces Lateral Bud Outgrowth by Modulating Cytokinin Biosynthesis and Signalling Pathways in Apple Identified via Transcriptome Analysis

Abstract
:1. Introduction
2. Results
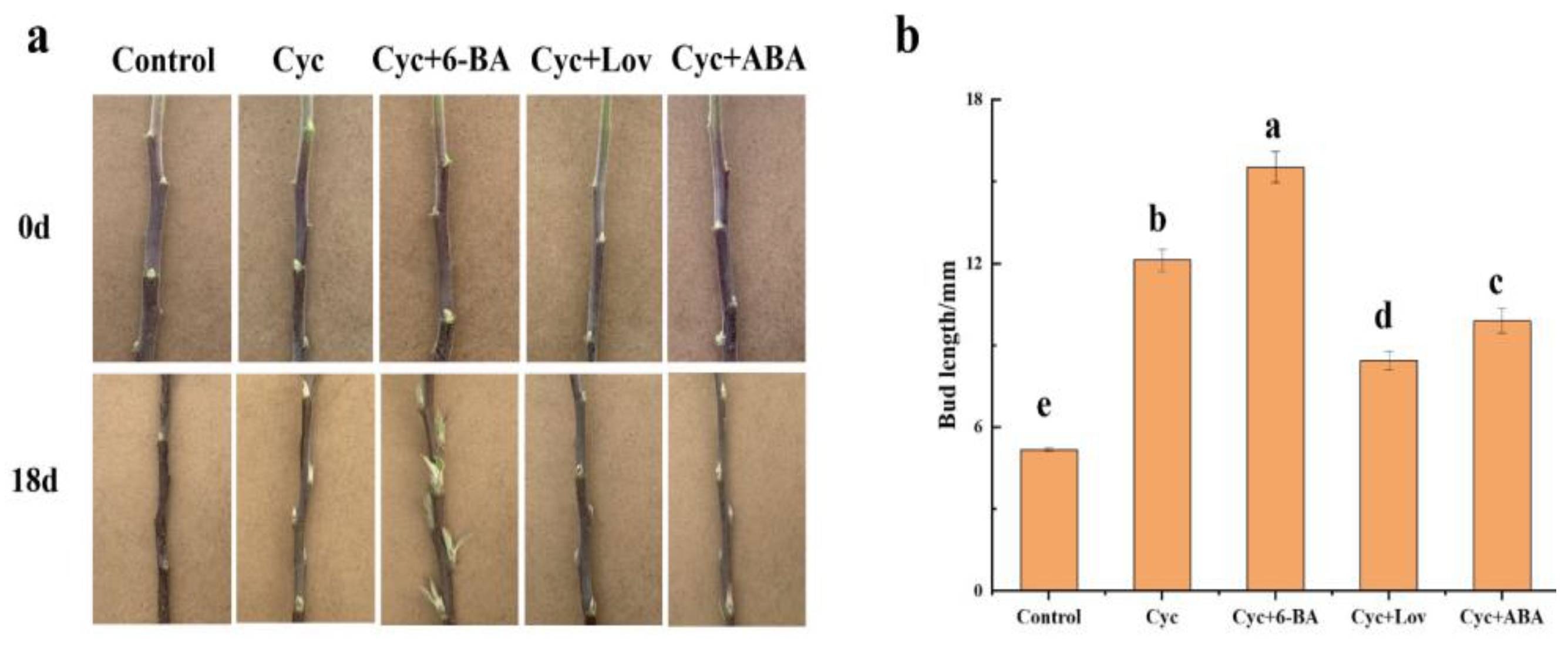
2.1. Influence of CYC on Bud Outgrowth Patterns
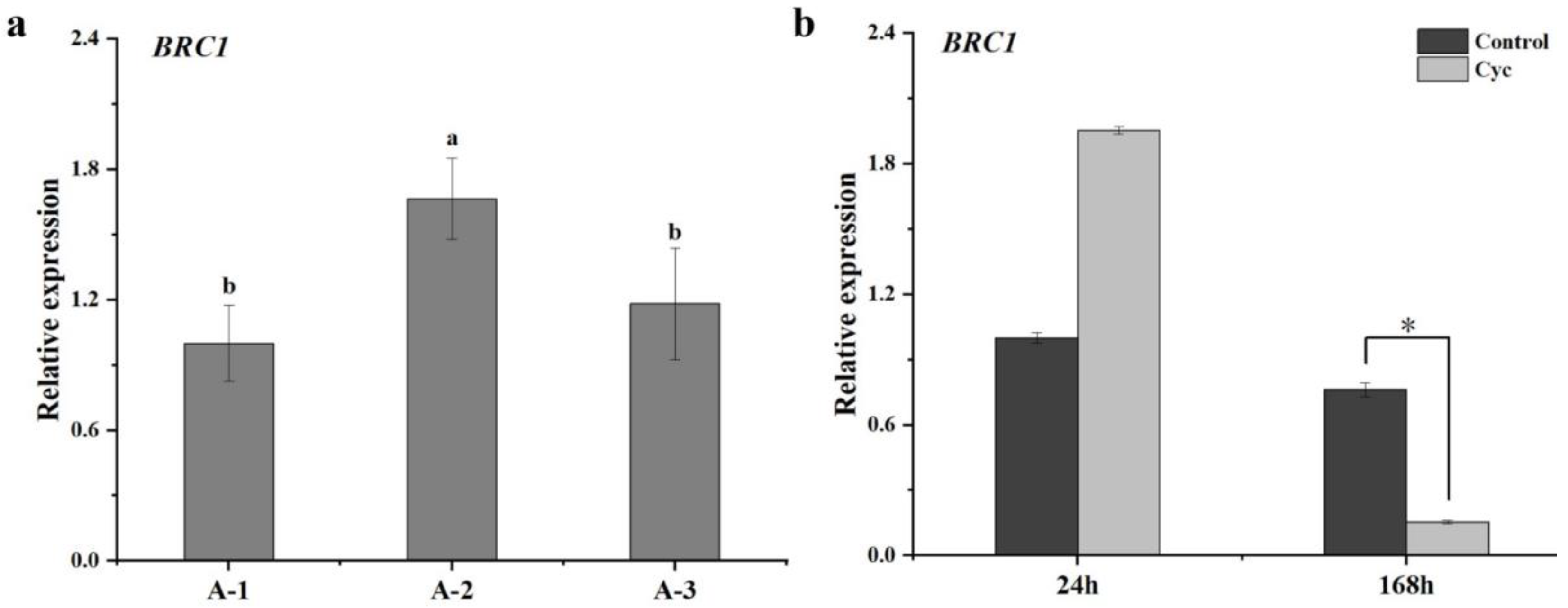
2.2. Influence of CYC on BRC1 Expression
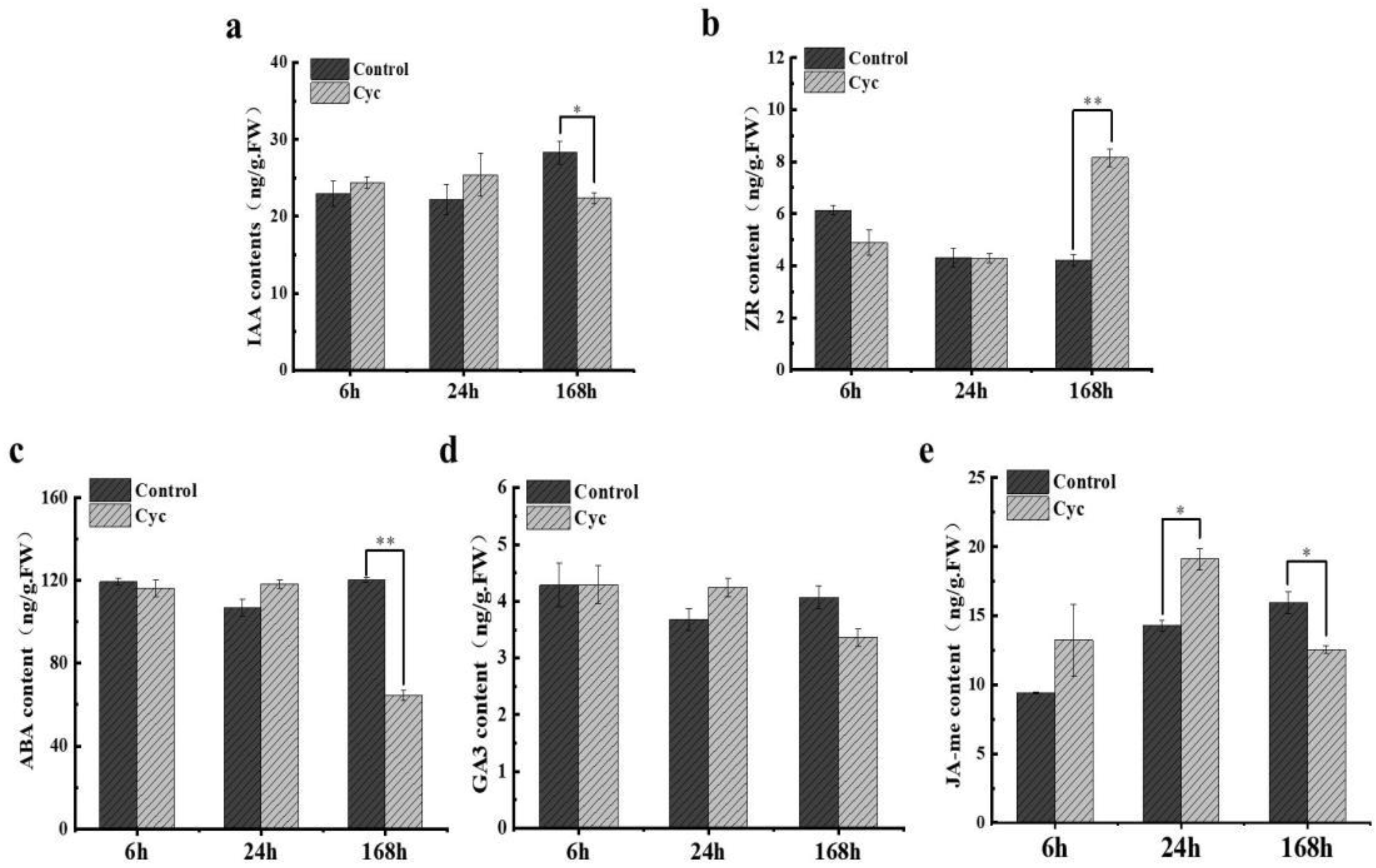
2.3. Impact of CYC on Phytohormones Concentration
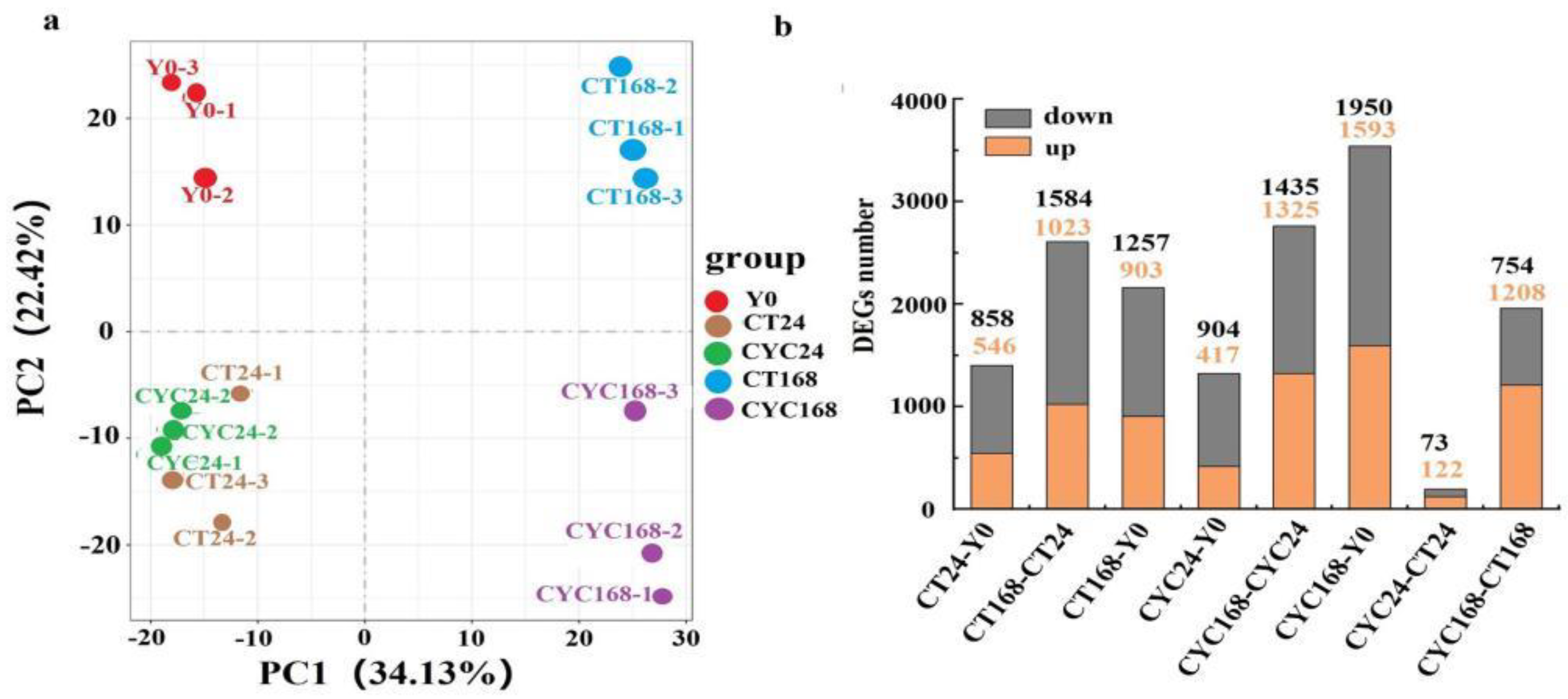
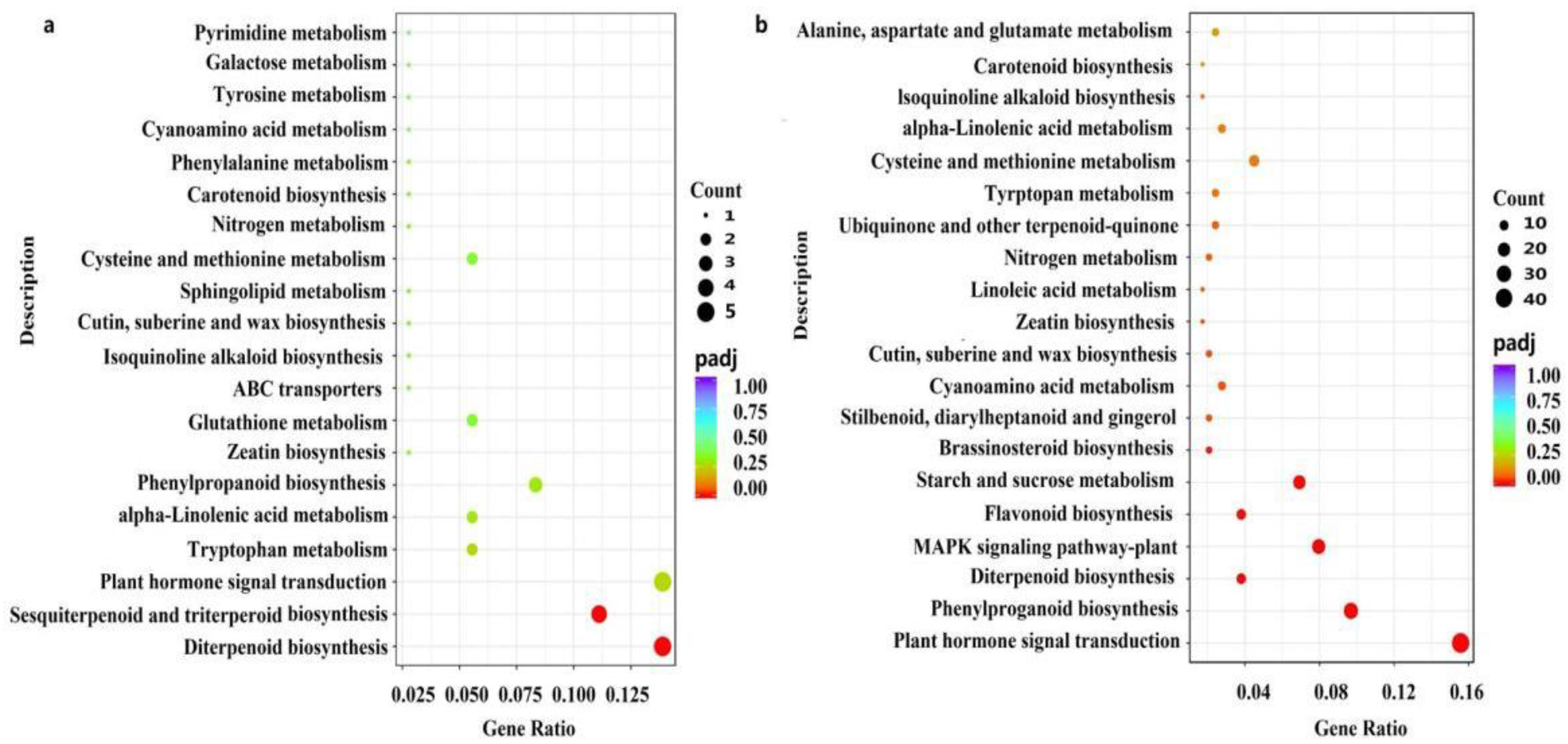
2.4. Genome-Wide Transcriptional Profiles of Axillary Bud after CYC Treatment
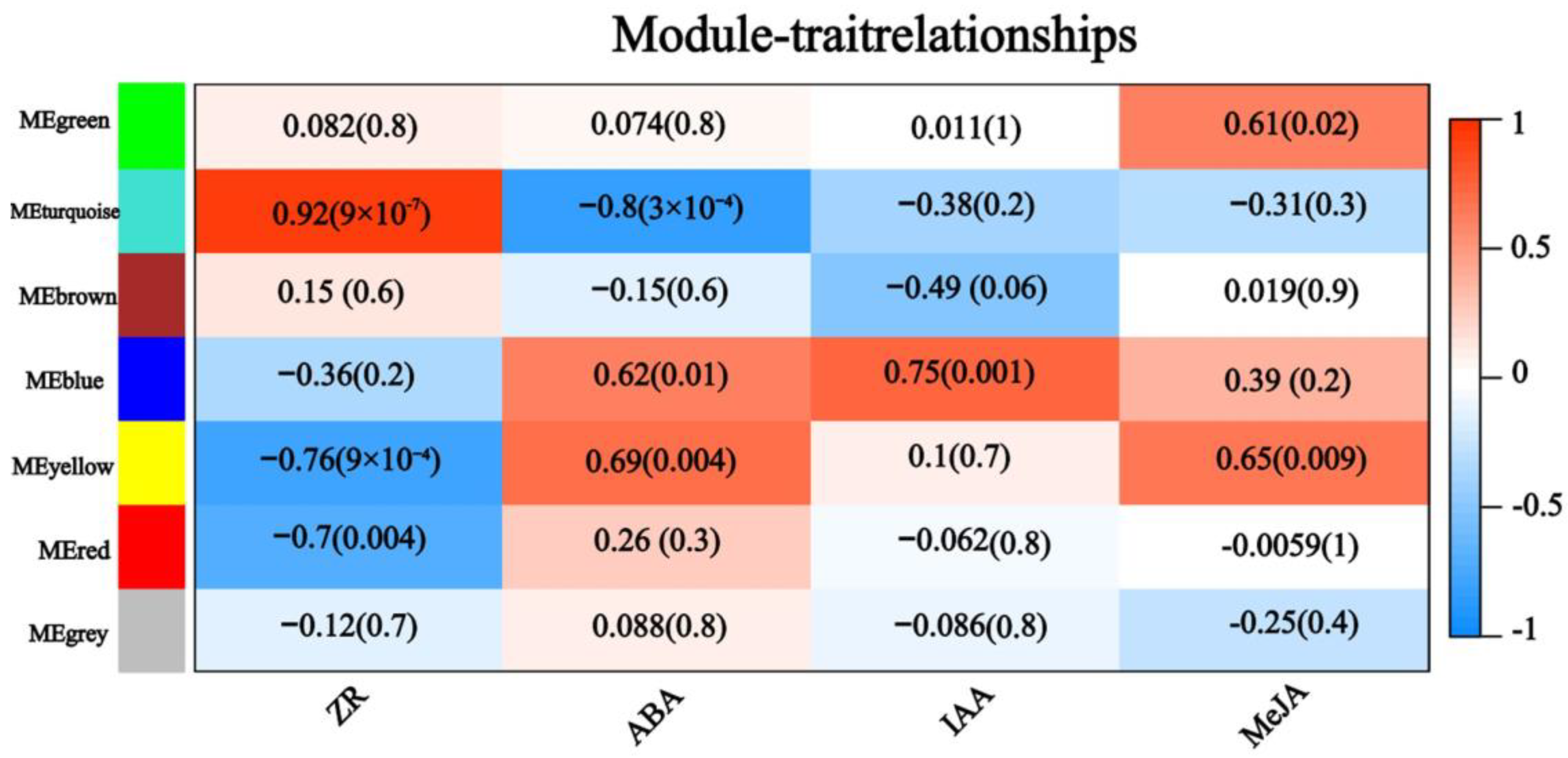
2.5. Weighted Gene Co-Expression Network Analysis of DEGs Produced by CYC Treatment
2.6. CTK and ABA Involved in CYC-Regulated Bud Outgrowth
3. Discussion
3.1. Effect of CYC on Bud Outgrowth in Apple
3.2. Effect of BRC1 on CYC-Controlled Branching
3.3. Effects of CTK and ABA on CYC-Induced Branching
3.4. Other Factors Involved in the Effect of CYC on Branching
4. Materials and Methods
4.1. Plant Materials
4.2. Experimental Arrangements
4.3. Hormone Measurements
4.4. RNA-seq Analysis
4.5. Quantification of Gene Expression
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burton, J.D.; Pedersen, M.K.; Coble, H.D. Effect of Cyclanilide on Auxin Activity. J. Plant Growth Regul. 2008, 27, 342–352. [Google Scholar] [CrossRef]
- Elfving, D.C.; Visser, D.B. Cyclanilide Induces Lateral Branching in Apple Trees. HortScience 2005, 40, 119–122. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Han, M.; Zhao, C. Research progress in promoting branching techniques in apple nursery and young tree. J. Fruit Sci. 2011, 28, 108–113. [Google Scholar]
- Robinson, T.; Sazo, M.M. Effect of promalin, benzyladenine and cyclanilide on lateral branching of apple trees in the nursery. Acta Hortic. 2014, 1042, 293–302. [Google Scholar] [CrossRef]
- Rameau, C.; Bertheloot, J.; LeDuc, N.; Andrieu, B.; Foucher, F.; Sakr, S. Multiple pathways regulate shoot branching. Front. Plant Sci. 2015, 5, 741. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Smith, S.M.; Li, J. Genetic Regulation of Shoot Architecture. Annu. Rev. Plant Biol. 2018, 69, 437–468. [Google Scholar] [CrossRef]
- Barbier, F.F.; Dun, E.A.; Kerr, S.C.; Chabikwa, T.G.; Beveridge, C.A. An Update on the Signals Controlling Shoot Branching. Trends Plant Sci. 2019, 24, 220–236. [Google Scholar] [CrossRef] [PubMed]
- Leyser, O. Regulation of shoot branching by auxin. Trends Plant Sci. 2003, 8, 541–545. [Google Scholar] [CrossRef]
- Domagalska, M.A.; Leyser, O. Signal integration in the control of shoot branching. Nat. Rev. Mol. Cell Biol. 2011, 12, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Shimizu-Sato, S.; Tanaka, M.; Mori, H. Auxin–cytokinin interactions in the control of shoot branching. Plant Mol. Biol. 2008, 69, 429–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, D.; Leyser, O. Auxin, cytokinin and the control of shoot branching. Ann. Bot. 2011, 107, 1203–1212. [Google Scholar] [CrossRef] [Green Version]
- Crawford, S.; Shinohara, N.; Sieberer, T.; Williamson, L.; George, G.; Hepworth, J.; Müller, D.; Domagalska, M.A.; Leyser, O. Strigolactones enhance competition between shoot branches by dampening auxin transport. Development 2010, 137, 2905–2913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldie, T.; Leyser, O. Cytokinin Targets Auxin Transport to Promote Shoot Branching. Plant Physiol. 2018, 177, 803–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, J.; Gao, C.; Chen, M.-S.; Pan, B.-Z.; Ye, K.; Xu, Z.-F. Gibberellin Promotes Shoot Branching in the Perennial Woody PlantJatropha curcas. Plant Cell Physiol. 2015, 56, 1655–1666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Grandío, E.; Pajoro, A.; Franco-Zorrilla, J.M.; Tarancón, C.; Immink, R.G.; Cubas, P. Abscisic acid signalling is con-trolled by a BRANCHED1/HD-ZIP I cascade in Arabidopsis axillary buds. Proc. Natl. Acad. Sci. USA 2016, 114, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Finlayson, S.A. Sorghum tiller bud growth is repressed by contact with the overlying leaf. Plant Cell Environ. 2019, 42, 2120–2132. [Google Scholar] [CrossRef]
- Xia, X.; Dong, H.; Yin, Y.; Song, X.; Gu, X.; Sang, K.; Zhou, J.; Shi, K.; Zhou, Y.; Foyer, C.H.; et al. Brassinosteroid signalling integrates multiple pathways to release apical dominance in tomato. Proc. Natl. Acad. Sci. USA 2021, 118, e2004384118. [Google Scholar] [CrossRef]
- Barbier, F.; Péron, T.; Lecerf, M.; Perez-Garcia, M.-D.; Barrière, Q.; Rolčík, J.; Boutet-Mercey, S.; Citerne, S.; Lemoine, R.; Porcheron, B.; et al. Sucrose is an early modulator of the key hormonal mechanisms controlling bud outgrowth in Rosa hybrida. J. Exp. Bot. 2015, 66, 2569–2582. [Google Scholar] [CrossRef] [Green Version]
- Bertheloot, J.; Barbier, F.; Boudon, F.; Perez-Garcia, M.D.; Péron, T.; Citerne, S.; Dun, E.; Beveridge, C.; Godin, C.; Sakr, S. Sugar availability suppresses the auxin-induced strigolactone pathway to promote bud outgrowth. New Phytol. 2019, 225, 866–879. [Google Scholar] [CrossRef] [Green Version]
- Aguilar-Martínez, J.A.; Poza-Carrión, C.; Cubas, P. Arabidopsis BRANCHED1Acts as an Integrator of Branching Signals within Axillary Buds. Plant Cell 2007, 19, 458–472. [Google Scholar] [CrossRef]
- Schneider, A.; Godin, C.; Boudon, F.; Demotes-Mainard, S.; Sakr, S.; Bertheloot, J. Light Regulation of Axillary Bud Outgrowth along Plant Axes: An Overview of the Roles of Sugars and Hormones. Front. Plant Sci. 2019, 10, 1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saracoglu, O.; Cebe, U. Cyclanilide treatments increase lateral branching of apple and pear nursery trees. Appl. Ecol. Environ. Res. 2018, 16, 4575–4583. [Google Scholar] [CrossRef]
- Rufato, L.; Marchioretto, L.; Orlandi, J.C.; Michelon, M.F.; Rossi, A.D.; Sander, G.F.; Macedo, T.A.D. Lateral branch induc-tion at nursery with growth regulators in ‘MAXI GALA’ apple trees grafted on four rootstocks. Sci. Hortic. 2019, 253, 349–357. [Google Scholar] [CrossRef]
- Elfving, D.C.; Visser, D.B. Cyclanilide Induces Lateral Branching in Sweet Cherry Trees. HortScience 2006, 41, 149–153. [Google Scholar] [CrossRef] [Green Version]
- Costes, E.; Crespel, L.; Denoyes, B.; Morel, P.; Demene, M.-N.; Lauri, P.; Wenden, B.; Denoyes, B.; Wenden, B. Bud structure, position and fate generate various branching patterns along shoots of closely related Rosaceae species: A review. Front. Plant Sci. 2014, 5, 666. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.; Yuan, C.; Yang, Q.; Yang, Y.; Cheng, T.; Wang, J.; Pan, H.; Zhang, Q. Morpho-physiological integrators, transcrip-tome and coexpression network analyses signify the novel molecular signatures associated with axillary bud in chrysanthemum. BMC Plant Biol. 2020, 20, 145. [Google Scholar] [CrossRef]
- Shinohara, N.; Taylor, C.; Leyser, O. Strigolactone Can Promote or Inhibit Shoot Branching by Triggering Rapid Depletion of the Auxin Efflux Protein PIN1 from the Plasma Membrane. PLOS Biol. 2013, 11, e1001474. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.J.; Zhang, Y.Q.; Ge, D.F.; Wang, Z.Y.; Song, W.Y.; Gu, R.; Che, G.; Cheng, Z.H.; Liu, R.Y.; Zhang, X.L. CsBRC1 inhib-its axillary bud outgrowth by directly repressing the auxin efflux carrier CsPIN3 in cucumber. Proc. Natl. Acad. Sci. USA 2019, 116, 17105–17114. [Google Scholar] [CrossRef] [Green Version]
- Robert, H.; Grunewald, W.; Sauer, M.; Cannoot, B.; Soriano, M.; Swarup, R.; Weijers, D.; Bennett, M.; Boutilier, K.; Friml, J. Plant embryogenesis requires AUX/LAX-mediated auxin influx. Development 2015, 142, 702–711. [Google Scholar] [CrossRef] [Green Version]
- Braun, N.; Germain, A.; Pillot, J.P.; Boutetmercey, S.; Dalmais, M.; Antoniadi, I.; Li, X.; Maia-Grondard, A.L.; Signor, C.; Bouteiller, N.; et al. The pea TCP transcription factor PsBRC1 acts down-stream of Strigolactones to control shoot branching. Plant Physiol. 2012, 158, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Dun, E.; Germain, A.D.S.; Rameau, C.; Beveridge, C.A. Antagonistic Action of Strigolactone and Cytokinin in Bud Outgrowth Control. Plant Physiol. 2011, 158, 487–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seale, M.; Bennett, T.; Leyser, O. BRC1 expression regulates bud activation potential, but is not necessary or sufficient for bud growth inhibition in Arabidopsis. Development 2017, 144, 1661–1673. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Ji, Y.; Hu, X.; Sun, S.; Wang, X. BES1 Functions as the Co-regulator of D53-like SMXLs to Inhibit BRC1 Expression in Strigolactone-Regulated Shoot Branching in Arabidopsis. Plant Commun. 2019, 1, 100014. [Google Scholar] [CrossRef] [PubMed]
- Van Rongen, M.; Bennett, T.; Ticchiarelli, F.; Leyse, O. Connective auxin transport contributes to strigolactone-mediated shoot branching control independent of the transcription factor BRC1. PLoS Genet. 2019, 15, e1008023. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Janssen, B.J.; Snowden, K.C. The molecular and genetic regulation of shoot branching. Plant Physiol. 2021, 187, 1033–1044. [Google Scholar] [CrossRef]
- Li, G.; Tan, M.; Cheng, F.; Liu, X.; Qi, S.; Chen, H.; Zhang, N.; Zhao, C.; Han, M.; Ma, J. Molecular role of cytokinin in bud activation and outgrowth in apple branching based on transcriptomic analysis. Plant Mol. Biol. 2018, 98, 261–274. [Google Scholar] [CrossRef]
- Tan, M.; Li, G.; Chen, X.; Xing, L.; Ma, J.; Zhang, D.; Ge, H.; Han, M.; Sha, G.; An, N. Role of Cytokinin, Strigolactone, and Auxin Export on Outgrowth of Axillary Buds in Apple. Front. Plant Sci. 2019, 10, 616. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Li, G.; Liu, X.; Cheng, F.; Ma, J.; Zhao, C.; Zhang, D.; Han, M. Exogenous application of GA3 inactively regulates axillary bud outgrowth by influencing of branching-inhibitors and bud-regulating hormones in apple (Malus domestica Borkh.). Mol. Genet. Genom. 2018, 293, 1547–1563. [Google Scholar] [CrossRef]
- Roman, H.; Girault, T.; Barbier, F.; Péron, T.; Brouard, N.; Pěnčík, A.; Novák, O.; Vian, A.; Sakr, S.; Lothier, J.; et al. Cytokinins Are Initial Targets of Light in the Control of Bud Outgrowth. Plant Physiol. 2016, 172, 489–509. [Google Scholar] [CrossRef] [Green Version]
- Kotov, A.A.; Kotova, L.M.; Romanov, G.A. Signaling network regulating plant branching: Recent advances and new chal-lenges. Plant Sci. 2021, 307, 110–880. [Google Scholar] [CrossRef]
- To, J.P.; Haberer, G.; Ferreira, F.J.; Deruère, J.; Mason, M.G.; Schaller, G.E.; Alonso, J.M.; Ecker, J.R.; Kieber, J.J. Type-A Arabidopsis Response Regulators Are Partially Redundant Negative Regulators of Cytokinin Signaling. Plant Cell 2004, 16, 658–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.; Liu, L.; Li, M.; Fang, S.; Shen, X.; Chu, J.; Zhang, Z. UNBRANCHED3 regulates branching by modulating cytokinin biosynthesis and signalling in maize and rice. New Phytol. 2017, 214, 721–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashikari, M.; Sakakibara, H.; Lin, S.; Yamamoto, T.; Takashi, T.; Nishimura, A.; Angeles, E.R.; Qian, Q.; Kitano, H.; Matsuoka, M. Cytokinin Oxidase Regulates Rice Grain Production. Science 2005, 309, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Yu, H.; Yuan, K.; Liao, Z.; Meng, X.; Jing, Y.; Liu, G.; Chu, J.; Li, J. Strigolactone promotes cytokinin degradation through transcriptional activation of CYTOKININ OXIDASE/DEHYDROGENASE 9in rice. Proc. Natl. Acad. Sci. USA 2019, 116, 14319–14324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holalu, S.V.; Finlayson, S.A. The ratio of red light to far red light alters Arabidopsis axillary bud growth and abscisic acid signalling before stem auxin changes. J. Exp. Bot. 2017, 68, 943–952. [Google Scholar] [CrossRef] [Green Version]
- Zhan, J.; Chu, Y.; Wang, Y.; Diao, Y.; Zhao, Y.; Liu, L.; Wei, X.; Meng, Y.; Li, F.; Ge, X. The miR164-GhCUC2-GhBRC1 mod-ule regulates plant architecture through abscisic acid in cotton. Plant Biotechnol. J. 2021, 19, 1839–1851. [Google Scholar] [CrossRef]
- Holalu, S.V.; Reddy, S.K.; Blackman, B.K.; Finlayson, S.A. Phytochrome interacting factors 4 and 5 regulate axillary branch-ing via bud abscisic acid and stem auxin signalling. Plant Cell Environ. 2020, 43, 2224–2238. [Google Scholar] [CrossRef]
- Merlot, S.; Gosti, F.; Guerrier, D.; Vavasseur, A.; Giraudat, J. The ABI1 and ABI2 protein phosphatases 2C act in a negative feedback regulatory loop of the abscisic acid signalling pathway. Plant J. 2001, 25, 295–303. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, S.; Zhu, W.; Jia, K.; Yang, H.; Wang, X. Strigolactone/MAX2-induced degradation of brassinosteroid tran-scriptional effector BES1 regulates shoot branching. Dev. Cell. 2013, 27, 681–688. [Google Scholar] [CrossRef] [Green Version]
- Mason, M.; Ross, J.J.; Babst, B.A.; Wienclaw, B.N.; Beveridge, C.A. Sugar demand, not auxin, is the initial regulator of apical dominance. Proc. Natl. Acad. Sci. USA 2014, 111, 6092–6097. [Google Scholar] [CrossRef] [Green Version]
- Barbier, F.F.; Cao, D.; Fichtner, F.; Weiste, C.; Perez-Garcia, M.D.; Caradeuc, M.; Gourrierec, J.L.; Le Gourrierec, J.; Sakr, S.; Beveridge, C.A. HEXOKINASE1 signalling promotes shoot branching and interacts with cytokinin and strigolactone path-ways. New Phytol. 2021, 231, 1088–1104. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.B.; Barbier, F.F.; Zhao, J.; Zafar, S.A.; Uzair, M.; Sun, Y.; Fang, J.; Perez-Garcia, M.; Bertheloot, J.; Sakr, S.; et al. Sucrose promotes D53 accumulation and tillering in rice. New Phytol. 2021. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, F.Y.; Ma, J.J.; Chen, H.; Xie, L.L.; Zhao, C.P. Effects of cyclanilide on growth and branching characteristics of 1-year old apple plants. J. Fruit Sci. 2021, 38, 1468–1478. [Google Scholar]
- Gambino, G.; Perrone, I.; Gribaudo, I. A Rapid and effective method for RNA extraction from different tissues of grapevine and other woody plants. Phytochem. Anal. 2008, 19, 520–525. [Google Scholar] [CrossRef]
- Li, G.; Ma, J.; Tan, M.; Mao, J.; An, N.; Sha, G.; Zhang, D.; Zhao, C.; Han, M. Transcriptome analysis reveals the effects of sugar metabolism and auxin and cytokinin signalling pathways on root growth and development of grafted apple. BMC Genom. 2016, 17, 150. [Google Scholar] [CrossRef] [Green Version]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guitton, B.; Kelner, J.J.; Celton, J.M.; Sabau, X.; Renou, J.P.; Costes, E. Analysis of transcripts differentially expressed between fruited and deflowered ‘gala’ adult trees: A contribution to biennial bearing understanding in apple. BMC Plant Biol. 2016, 16, 55. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | FPKM | Description | |
|---|---|---|---|
| CYC168 | CT168 | ||
| MD15G1365900 | 0.8 | 6.8 | Haloacid dehalogenase-like hydrolase (HAD) superfamily protein |
| MD15G1326700 | 1.7 | 6.8 | Haloacid dehalogenase-like hydrolase (HAD) superfamily protein |
| MD14G1183700 | 1.7 | 4.9 | Trehalose-phosphatase/synthase 9 |
| MD00G1171800 | 4.9 | 14.5 | Trehalose-phosphatase |
| MD13G1089600 | 3.3 | 9.7 | Trehalose-phosphatase/synthase 7 |
| MD08G1180400 | 2.8 | 7.7 | Haloacid dehalogenase-like hydrolase (HAD) superfamily protein |
| MD09G1234800 | 6.0 | 14.3 | Trehalose-phosphatase/synthase 7 |
| MD15G1223500 | 40.3 | 84.2 | Sucrose synthase 4 |
| MD06G1237200 | 0.9 | 1.8 | Sucrose synthase 6 |
| MD09G1192100 | 4.5 | 2.2 | O-Glycosyl hydrolases family 17 protein |
| MD14G1004200 | 4.3 | 2.0 | Glycosyl hydrolase family protein |
| MD11G1270400 | 11.5 | 5.2 | Trehalose-6-phosphate synthase |
| MD13G1186100 | 5.8 | 2.5 | ADP glucose pyrophosphorylase |
| MD11G1195800 | 53.9 | 23.4 | Glycosyl hydrolase family protein |
| MD15G1113000 | 6.8 | 2.9 | O-Glycosyl hydrolases family 17 protein |
| MD11G1178000 | 20.0 | 7.8 | Glycosyl hydrolase family protein |
| MD13G1030200 | 10.8 | 4.2 | O-Glycosyl hydrolases family 17 protein |
| MD11G1240900 | 27.9 | 9.5 | Beta glucosidase 46 |
| MD14G1128000 | 56.4 | 19.3 | Glycosyl hydrolase 9C2 |
| MD10G1316100 | 64.6 | 13.2 | Glucose-6-phosphate/phosphate translocator 2 |
| Gene ID | FPKM | Description | Pathway | |
|---|---|---|---|---|
| CYC168 | CT168 | |||
| MD13G1058000 | 0.8 | 2.7 | Cytochrome P450 superfamily protein | BR biosynthesis |
| MD17G1064800 | 1.9 | 3.9 | Brassinosteroid-6-oxidase 2 | BR biosynthesis |
| MD06G1146700 | 8.2 | 21.0 | Cytochrome P450 superfamily protein | BR biosynthesis |
| MD06G1146000 | 23.0 | 46.9 | Cytochrome P450 superfamily protein | BR biosynthesis |
| MD17G1167200 | 6.7 | 3.2 | Cytochrome P450 superfamily protein | BR biosynthesis |
| MD17G1120200 | 1.6 | 0.0 | Cytochrome P450 superfamily protein DWF4 | BR biosynthesis |
| MD14G1078600 | 0.5 | 6.1 | Cytokinin oxidase/dehydrogenase 1 | Zeatin biosynthesis |
| MD16G1041700 | 0.9 | 2.8 | Isopentenyl transferase 1 | Zeatin biosynthesis |
| MD03G1267900 | 2.6 | 6.3 | Isopentenyl transferase 3 | Zeatin biosynthesis |
| MD17G1076700 | 1.9 | 0.8 | Cytochrome P450, family 735, subfamily A, polypeptide 1 | Zeatin biosynthesis |
| MD15G1208300 | 4.0 | 0.5 | Cytokinin oxidase/dehydrogenase 3 | Zeatin biosynthesis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, J.; Xie, L.; Zhao, Q.; Sun, Y.; Zhang, D. Cyclanilide Induces Lateral Bud Outgrowth by Modulating Cytokinin Biosynthesis and Signalling Pathways in Apple Identified via Transcriptome Analysis. Int. J. Mol. Sci. 2022, 23, 581. https://doi.org/10.3390/ijms23020581
Ma J, Xie L, Zhao Q, Sun Y, Zhang D. Cyclanilide Induces Lateral Bud Outgrowth by Modulating Cytokinin Biosynthesis and Signalling Pathways in Apple Identified via Transcriptome Analysis. International Journal of Molecular Sciences. 2022; 23(2):581. https://doi.org/10.3390/ijms23020581
Chicago/Turabian StyleMa, Juanjuan, Lingling Xie, Qian Zhao, Yiting Sun, and Dong Zhang. 2022. "Cyclanilide Induces Lateral Bud Outgrowth by Modulating Cytokinin Biosynthesis and Signalling Pathways in Apple Identified via Transcriptome Analysis" International Journal of Molecular Sciences 23, no. 2: 581. https://doi.org/10.3390/ijms23020581