

Purification, Biochemical and Kinetic Characterization of a Novel Alkaline sn-1,3-Regioselective Triacylglycerol Lipase from Penicilliumcrustosum Thom Strain P22 Isolated from Moroccan Olive Mill Wastewater
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Fungal Isolation and Identification
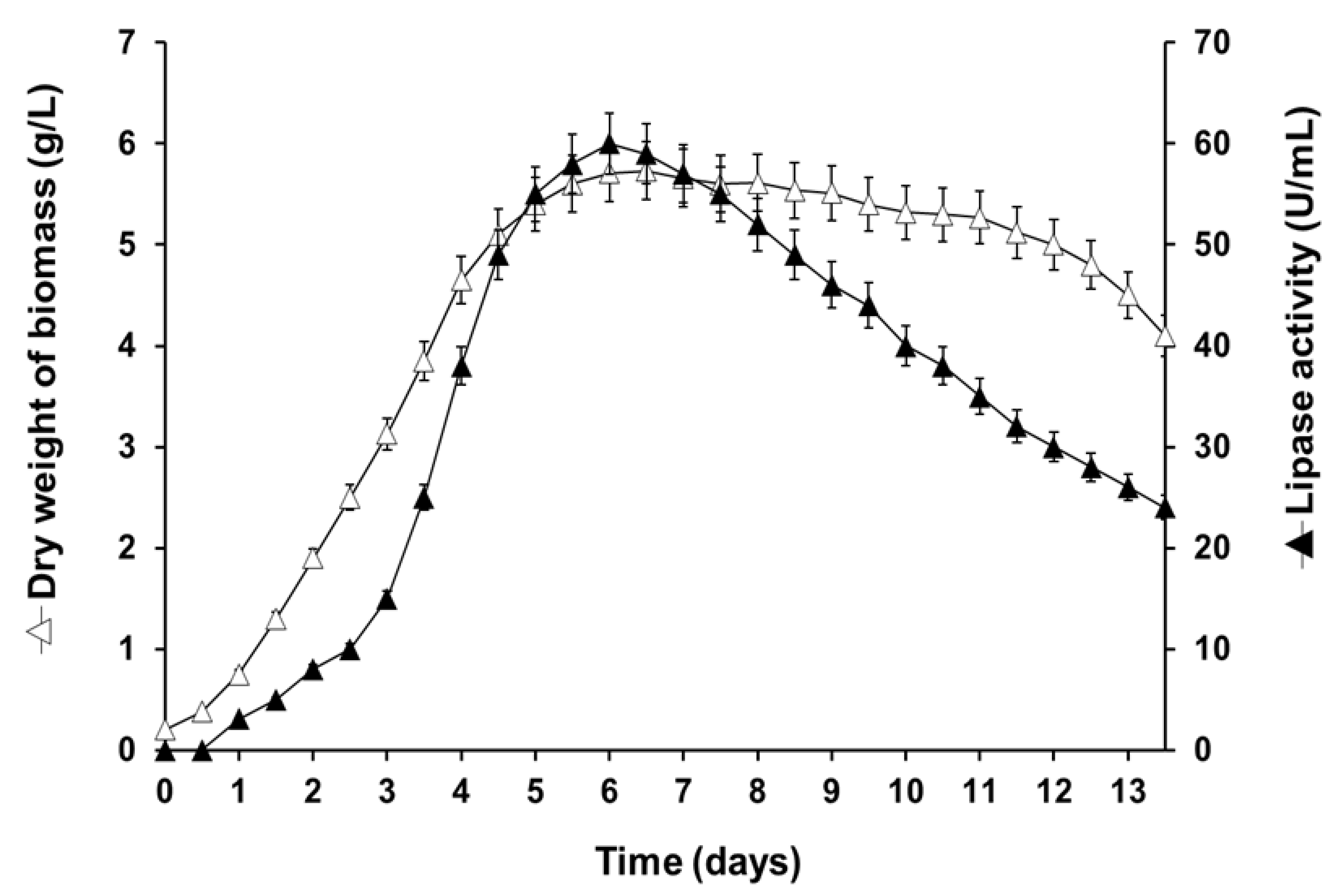
2.2. Lipase Production
2.3. Purification of PCrL
2.4. Biochemical Characterization of PCrL
2.4.1. Effects of Temperature on PCrL Activity and Stability
2.4.2. Effects of pH on PCrL Activity and Stability
2.4.3. Effect of Inhibitors, Metal Ions, and Bile Salts on the PCrL Activity
2.5. Substrate Specificity
2.6. Enzymatic Performance of the Purified PCrL
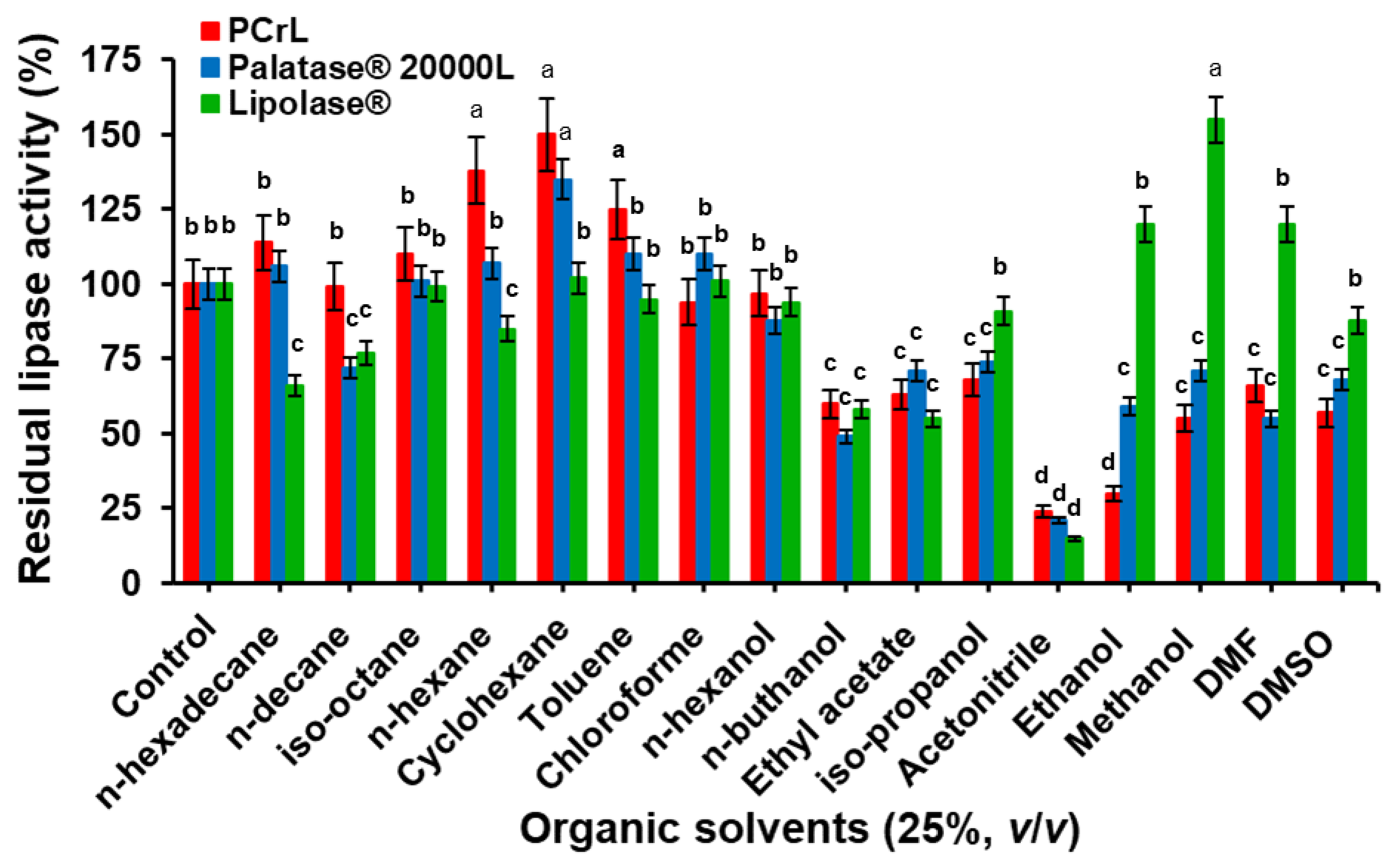
2.6.1. Effect of Organic Solvents on Enzyme Stability
2.6.2. Influence of Some Laundry Detergent Constituents on PcrL Stability
2.6.3. PCrL Compatibility with Laundry Detergents
2.6.4. Wash Performance Test on Oil Removal with PCrL
3. Discussion
4. Material and Methods
4.1. Material
4.2. Lipids
4.3. Isolation of a Lipase-Producing Strain, Media, and Culture Conditions
4.4. Taxonomic Identification of Fungal Strain P22
4.5. Lipase Activity Measurement
4.6. Lipase Production
4.7. Analytical Methods
4.8. Purification Procedure
4.9. Biochemical Characterization of the Purified PCrL
4.9.1. Effects of Temperature on Lipase Activity and Stability
4.9.2. Effects of pH on Lipase Activity and Stability
4.9.3. Substrate Specificity
4.9.4. Effects of Inhibitors, Reducing Agents, Chelating Reagents, Metal Ions, and Bile Salts on PCrL Activity
4.10. Performance Evaluation of the Purified PCrL Compared with Palatase® 20000 L and Lipolase®
4.10.1. Effect of Organic Solvents on Lipase Stability
4.10.2. Effect of Some Detergent Additives on PCrL Stability
4.10.3. Washing Performance Analysis of PCrL
4.11. Statistical Analysis
4.12. Culture Collection Depository’s Numbers and Nucleotide Sequence Accession Numbers
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Olive Oil Council (IOOC) 2021/2022, Olive Oil Production, Consumption, Export. IOOC, Madrid. Available online: https://www.internationaloliveoil.org/wp-content/uploads/2022/08/IOC-Imports-2021-22-1.html (accessed on 15 August 2022).
- Zahi, M.R.; Zam, W.; El Hattab, M. State of knowledge on chemical, biological and nutritional properties of olive mill wastewater. Food Chem. 2022, 381, 132238. [Google Scholar] [CrossRef]
- Misbah, A.; Aouine, M.; Raouan, S.E.; Lekbach, Y.; Ettadili, H.; Koraichi, S.I.; Jouti, M.A.T. Microorganisms isolated from moroccan olive-mill wastes: Screening of their enzymatic activities for biotechnological use. Eur. Sci. J. 2019, 15, 464. [Google Scholar] [CrossRef] [Green Version]
- Szymczak, T.; Cybulska, J.; Podleśny, M.; Frąc, M. Various perspectives on microbial lipase production using agri-food waste and renewable products. Agriculture 2021, 11, 540. [Google Scholar] [CrossRef]
- Abrunhosa, L.; Oliveira, F.; Dantas, D.; Goncalves, C.; Belo, I. Lipase production by Aspergillus ibericus using olive mill wastewater. Bioprocess Biosyst. Eng. 2013, 36, 285–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verger, R. ‘Interfacial activation’of lipases: Facts and artifacts. Trends Biotechnol. 1997, 15, 32–38. [Google Scholar] [CrossRef]
- Jiao, L.; Zhou, Q.; Su, Z.; Yan, Y. Efficient heterologous production of Rhizopus oryzae lipase via optimization of multiple expression-related helper proteins. Int. J. Mol. Sci. 2018, 19, 3372. [Google Scholar] [CrossRef] [Green Version]
- Singhania, V.; Cortes-Clerget, M.; Dussart-Gautheret, J.; Akkachairin, B.; Yu, J.; Akporji, N.; Gallou, F.; Lipshutz, B.H. Lipase-catalyzed esterification in water enabled by nanomicelles. Applications to 1-pot multi-step sequences. Chem. Sci. 2022, 13, 1440–1445. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Ma, M.; Yan, X.; Zhang, G.; Xia, J.; Zeng, Z.; Yu, P.; Deng, Q.; Gong, D. Green synthesis of polydopamine functionalized magnetic mesoporous biochar for lipase immobilization and its application in interesterification for novel structured lipids production. Food Chem. 2022, 379, 132148. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.; Bhatti, H.N.; Nawaz, S.; Bilal, M. Penicillium fellutanum lipase as a green and ecofriendly biocatalyst for depolymerization of poly (varepsilon-caprolactone): Biochemical, kinetic, and thermodynamic investigations. Biotechnol. Appl. Biochem. 2022, 69, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Boran, R.; Ugur, A.; Sarac, N.; Ceylan, O. Characterisation of Streptomyces violascens OC125-8 lipase for oily wastewater treatment. Biotech 2019, 9, 5. [Google Scholar] [CrossRef]
- Tian, M.; Yang, L.; Lv, P.; Wang, Z.; Fu, J.; Miao, C.; Li, Z.; Li, L.; Liu, T.; Du, W.; et al. Improvement of methanol tolerance and catalytic activity of Rhizomucor miehei lipase for one-step synthesis of biodiesel by semi-rational design. Bioresour. Technol. 2022, 348, 126769. [Google Scholar] [CrossRef] [PubMed]
- Ortega, N.; Sáez, L.; Palacios, D.; Busto, M.D. Kinetic Modeling, Thermodynamic approach and molecular dynamics simulation of thermal inactivation of lipases from Burkholderia cepacia and Rhizomucor miehei. Int. J. Mol. Sci. 2022, 23, 6828. [Google Scholar] [CrossRef] [PubMed]
- Chandra, P.; Singh, R.; Arora, P.K. Microbial lipases and their industrial applications: A comprehensive review. Microb. Cell Fact. 2020, 19, 1–42. [Google Scholar] [CrossRef] [PubMed]
- Dheeman, D.S.; Antony-Babu, S.; Frías, J.M.; Henehan, G.T. Purification and characterization of an extracellular lipase from a novel strain Penicillium sp. DS-39 (DSM 23773). J. Mol. Catal. B: Enzym. 2011, 72, 256–262. [Google Scholar] [CrossRef]
- De Menezes, L.H.S.; Ramos, M.; Araujo, S.C.; Santo, E.; Oliveira, P.C.; Tavares, I.M.C.; Santos, P.H.; Franco, M.; de Oliveira, J.R. Application of a constrained mixture design for lipase production by Penicillium roqueforti ATCC 10110 under solid-state fermentation and using agro-industrial wastes as substrate. Prep. Biochem. Biotechnol. 2021, 52, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Griebeler, N.; Polloni, A.E.; Remonatto, D.; Arbter, F.; Vardanega, R.; Cechet, J.L.; Di Luccio, M.; de Oliveira, D.; Treichel, H.; Cansian, R.L. Isolation and screening of lipase-producing fungi with hydrolytic activity. Food Bioprocess Technol. 2011, 4, 578–586. [Google Scholar] [CrossRef]
- Lima, V.; Krieger, N.; Mitchell, D.; Fontana, J. Activity and stability of a crude lipase from Penicillium aurantiogriseum in aqueous media and organic solvents. Biochem. Eng. J. 2004, 18, 65–71. [Google Scholar] [CrossRef]
- Sowa, M.A.; Kreuter, N.; Sella, N.; Albuquerque, W.; Manhard, J.; Siegl, A.; Ghezellou, P.; Li, B.; Spengler, B.; Weichhard, E.; et al. Replacement of pregastric lipases in cheese production: Identification and heterologous expression of a lipase from Pleurotus citrinopileatus. J. Agric. Food Chem. 2022, 70, 2998–3008. [Google Scholar] [CrossRef] [PubMed]
- Lima, V.M.; Krieger, N.; Sarquis, M.I.M.; Mitchell, D.A.; Ramos, L.P.; Fontana, J.D. Effect of nitrogen and carbon sources on lipase production by Penicillium aurantiogriseum. Food Technol. Biotechnol. 2003, 41, 105–110. [Google Scholar]
- Devi, R.; Madhavan Nampoothiri, K.; Sukumaran, R.K.; Sindhu, R.; Arumugam, M. Lipase of Pseudomonas guariconesis as an additive in laundry detergents and transesterification biocatalysts. J. Basic Microbiol. 2020, 60, 112–125. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zong, M.H. Lipases from the genus Penicillium: Production, purification, characterization and applications. J. Mol. Catal. B. Enzym. 2010, 66, 43–54. [Google Scholar] [CrossRef]
- Asgher, M.; Ramzan, M.; Bilal, M. Purification and characterization of manganese peroxidases from native and mutant Trametes versicolor IBL-04. Chin. J. Catal. 2016, 37, 561–570. [Google Scholar] [CrossRef]
- Rigo, E.; Ninow, J.L.; Tsai, S.M.; Durrer, A.; Foltran, L.L.; Remonatto, D.; Sychoski, M.; Vardanega, R.; de Oliveira, D.; Treichel, H. Preliminary characterization of novel extra-cellular lipase from Penicillium crustosum under solid-state fermentation and its potential application for triglycerides hydrolysis. Food Bioprocess Technol. 2012, 5, 1592–1600. [Google Scholar] [CrossRef]
- El-Ghonemy, D.H.; El-Gamal, M.S.; Tantawy, A.E.; Ali, T.H. Extracellular alkaline lipase from a novel fungus Curvularia sp. DHE 5: Optimisation of physicochemical parameters, partial purification and characterisation. Food Technol. Biotechnol. 2017, 55, 206–217. [Google Scholar] [CrossRef]
- Alhelli, A.M.; Abdul Manap, M.Y.; Mohammed, A.S.; Mirhosseini, H.; Suliman, E.; Shad, Z.; Mohammed, N.K.; Meor Hussin, A.S. Use of response surface methodology for partitioning, one-step purification of alkaline extracellular lipase from Penicillium candidum (PCA 1/TT031). J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2016, 1039, 66–73. [Google Scholar] [CrossRef]
- Tiss, A.; Miled, N.; Verger, R.; Gargouri, Y.; Abousalham, A. Digestive lipases inhibition: An in vitro study. In Lipases and Phospholipases in Drug Development: From Biochemistry to Molecular Pharmacology; Müller, G., Petry, S., Eds.; Wiley-VCH: Weinheim, Germany, 2004; pp. 155–193. [Google Scholar]
- El Alaoui, M.; Soulère, L.; Noiriel, A.; Queneau, Y.; Abousalham, A. α-Eleostearic acid-containing triglycerides for a continuous assay to determine lipase sn-1 and sn-3 regio-preference. Chem. Phys. Lipids 2017, 206, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Eddehech, A.; Rahier, R.; Smichi, N.; Arhab, Y.; Noiriel, A.; Abousalham, A.; Sayari, A.; Zarai, Z. Heterologous expression, kinetic characterization and molecular modeling of a new sn-1, 3-regioselective triacylglycerol lipase from Serratia sp. W3. Process Biochem. 2021, 103, 87–97. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seifert, K.A.; Samson, R.A.; Dewaard, J.R.; Houbraken, J.; Lévesque, C.A.; Moncalvo, J.M.; Louis-Seize, G.; Hebert, P.D. Prospects for fungus identification using CO1 DNA barcodes, with Penicillium as a test case. Proc. Natl. Acad. Sci. USA 2007, 104, 3901–3906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouhibi, S.; Santos, C.; Ghali, R.; Soares, C.; Hedhili, A.; Paterson, R.; Lima, N. Penicillium tunisiense sp. nov., a novel species of Penicillium section Ramosa discovered from Tunisian orchard apples. Int. J. Syst. Evol. Microbiol. 2018, 68, 3217–3225. [Google Scholar] [CrossRef] [PubMed]
- Visagie, C.; Houbraken, J.; Frisvad, J.C.; Hong, S.B.; Klaassen, C.; Perrone, G.; Seifert, K.; Varga, J.; Yaguchi, T.; Samson, R. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2005, 53, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, N.; Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of the genus Talaromyces. Stud. Mycol. 2014, 78, 175–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rong, C.; Ma, Y.; Wang, S.; Liu, Y.; Wang, L.; Ma, K.; Dou, S.; Yang, Y.; Xu, F. Penicillium chroogomphum, a new species in Penicillium section Ramosa isolated from fruiting bodies of Chroogomphus rutilus in China. Mycoscience 2016, 57, 79–84. [Google Scholar] [CrossRef]
- Houbraken, J.; Kocsube, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.A.; et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef]
- Rehman, S.; Bhatti, H.N.; Bilal, M.; Asgher, M.; Wang, P. Catalytic, kinetic and thermodynamic characteristics of an extracellular lipase from Penicillium notatum. Catal. Lett. 2017, 147, 281–291. [Google Scholar] [CrossRef]
- Tan, T.; Zhang, M.; Xu, J.; Zhang, J. Optimization of culture conditions and properties of lipase from Penicillium camembertii Thom PG-3. Process Biochem. 2004, 39, 1495–1502. [Google Scholar] [CrossRef]
- Lianghua, T.; Liming, X.; Min, S.; Huaying, G. Purification and application of a lipase from Penicillium expansum PED-03. Appl. Biochem. Biotechnol. 2007, 142, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Bian, C.; Yuan, C.; Lin, L.; Lin, J.; Shi, X.; Ye, X.; Huang, Z.; Huang, M. Purification and preliminary crystallographic analysis of a Penicillium expansum lipase. Biochim. Biophys. Acta 2005, 1752, 99–102. [Google Scholar] [CrossRef]
- Ruiz, B.; Farres, A.; Langley, E.; Masso, F.; Sanchez, S. Purification and characterization of an extracellular lipase from Penicillium candidum. Lipids 2001, 36, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Chahinian, H.; Vanot, G.; Ibrik, A.; Rugani, N.; Sarda, L.; Comeau, L.C. Production of extracellular lipases by Penicillium cyclopium purification and characterization of a partial acylglycerol lipase. Biosci. Biotechnol. Biochem. 2000, 64, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.B.; Li, J.F.; Li, X.T.; Gu, Y.; Wu, M.C.; Wu, J.; Wang, J.Q. A unique mono-and diacylglycerol lipase from Penicillium cyclopium: Heterologous expression, biochemical characterization and molecular basis for its substrate selectivity. PLoS ONE 2014, 9, e102040. [Google Scholar] [CrossRef] [PubMed]
- Stöcklein, W.; Sztajer, H.; Menge, U.; Schmid, R. Purification and properties of a lipase from Penicillium expansum. Biochim. Biophys. Acta. Lipids Lipid Metab. 1993, 1168, 181–189. [Google Scholar] [CrossRef]
- Paluzar, H.; Tuncay, D.; Aydogdu, H. Production and characterization of lipase from Penicillium aurantiogriseum under solid-state fermentation using sunflower pulp. Biocatal. Biotransformation 2021, 39, 333–342. [Google Scholar] [CrossRef]
- Krieger, N.; Taipa, M.; Melo, E.; Lima-Filho, J.; Aires-Barros, M.; Cabral, J. Purification of a Penicillium citrinum lipase by chromatographic processes. Bioprocess Eng. 1999, 20, 59–65. [Google Scholar] [CrossRef]
- Gupta, R.; Gigras, P.; Mohapatra, H.; Goswami, V.K.; Chauhan, B. Microbial α-amylases: A biotechnological perspective. Process Biochem. 2003, 38, 1599–1616. [Google Scholar] [CrossRef]
- Niyonzima, F.; More, S. Microbial detergent compatible lipases. J. Sci. Ind. Res. 2015, 74, 105–113. [Google Scholar]
- Haddar, A.; Agrebi, R.; Bougatef, A.; Hmidet, N.; Sellami-Kamoun, A.; Nasri, M. Two detergent stable alkaline serine-proteases from Bacillus mojavensis A21: Purification, characterization and potential application as a laundry detergent additive. Bioresour. Technol. 2009, 100, 3366–3373. [Google Scholar] [CrossRef]
- Bancerz, R.; Ginalska, G.; Fiedurek, J.; Gromada, A. Cultivation conditions and properties of extracellular crude lipase from the psychrotrophic fungus Penicillium chrysogenum 9′. J. Ind. Microbiol. Biotechnol. 2005, 32, 253–260. [Google Scholar] [CrossRef]
- Rahman, R.N.; Baharum, S.N.; Basri, M.; Salleh, A.B. High-yield purification of an organic solvent-tolerant lipase from Pseudomonas sp. strain S5. Anal. Biochem. 2005, 341, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Chinaglia, S.; Chiarelli, L.R.; Maggi, M.; Rodolfi, M.; Valentini, G.; Picco, A.M. Biochemistry of lipolytic enzymes secreted by Penicillium solitum and Cladosporium cladosporioides. Biosci. Biotechnol. Biochem. 2014, 78, 245–254. [Google Scholar] [CrossRef]
- Pranav, K.; Nidhi, H.; Preeti, N.; Savita, K. Screening of lipase producing fungi and its application. J. Res. Anal. Rev. 2019, 2, 649i–655i. [Google Scholar]
- Akmoussi-Toumi, S.; Khemili-Talbi, S.; Ferioune, I.; Kebbouche-Gana, S. Purification and characterization of an organic solvent-tolerant and detergent-stable lipase from Haloferax mediterranei CNCMM 50101. Int. J. Biol. Macromol. 2018, 116, 817–830. [Google Scholar] [CrossRef] [PubMed]
- Gurkok, S.; Ozdal, M. Purification and characterization of a novel extracellular, alkaline, thermoactive, and detergent-compatible lipase from Aeromonas caviae LipT51 for application in detergent industry. Protein Expr. Purif. 2021, 180, 105819. [Google Scholar] [CrossRef] [PubMed]
- Invernizzi, G.; Papaleo, E.; Grandori, R.; De Gioia, L.; Lotti, M. Relevance of metal ions for lipase stability: Structural rearrangements induced in the Burkholderia glumae lipase by calcium depletion. J. Struct. Biol. 2009, 168, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Aloulou, A.; Puccinelli, D.; De Caro, A.; Leblond, Y.; Carrière, F. A comparative study on two fungal lipases from Thermomyces lanuginosus and Yarrowia lipolytica shows the combined effects of detergents and pH on lipase adsorption and activity. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2007, 1771, 1446–1456. [Google Scholar] [CrossRef]
- Gutarra, M.L.; Godoy, M.G.; Maugeri, F.; Rodrigues, M.I.; Freire, D.M.; Castilho, L.R. Production of an acidic and thermostable lipase of the mesophilic fungus Penicillium simplicissimum by solid-state fermentation. Bioresour. Technol. 2009, 100, 5249–5254. [Google Scholar] [CrossRef]
- Gupta, R.; Rathi, P.; Bradoo, S. Lipase mediated upgradation of dietary fats and oils. Crit. Rev. Food Sci. Nutr. 2003, 43, 635–644. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, L.; Liu, L.; Liu, T.; Li, C.; Sun, L. 1, 3-Dioleoyl-2-palmitoylglycerol-rich triacylglycerol characterization by three processing methods. Int. J. Food Prop. 2019, 22, 1156–1171. [Google Scholar] [CrossRef] [Green Version]
- Esteban, L.; Jiménez, M.J.; Hita, E.; González, P.A.; Martín, L.; Robles, A. Production of structured triacylglycerols rich in palmitic acid at sn-2 position and oleic acid at sn-1, 3 positions as human milk fat substitutes by enzymatic acidolysis. Biochem. Eng. J. 2011, 54, 62–69. [Google Scholar] [CrossRef]
- Doukyu, N.; Ogino, H. Organic solvent-tolerant enzymes. Biochem. Eng. J. 2010, 48, 270–282. [Google Scholar] [CrossRef]
- Bancerz, R.; Ginalska, G. A novel thermostable lipase from Basidiomycete Bjerkandera adusta R59: Characterisation and esterification studies. J. Ind. Microbiol. Biotechnol. 2007, 34, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Ogino, H.; Ishikawa, H. Enzymes which are stable in the presence of organic solvents. J. Biosci. Bioeng. 2001, 91, 109–116. [Google Scholar] [CrossRef]
- Turati, D.F.M.; Almeida, A.F.; Terrone, C.C.; Nascimento, J.M.; Terrasan, C.R.; Fernandez-Lorente, G.; Pessela, B.C.; Guisan, J.M.; Carmona, E.C. Thermotolerant lipase from Penicillium sp. section Gracilenta CBMAI 1583: Effect of carbon sources on enzyme production, biochemical properties of crude and purified enzyme and substrate specificity. Biocatal. Agric. Biotechnol. 2019, 17, 15–24. [Google Scholar]
- Careri, G. Cooperative charge fluctuations by migrating protons in globular proteins. Prog. Biophys. Mol. Biol. 1998, 70, 223–249. [Google Scholar] [CrossRef]
- Serdakowski, A.L.; Dordick, J.S. Enzyme activation for organic solvents made easy. Trends Biotechnol. 2008, 26, 48–54. [Google Scholar] [CrossRef]
- Mechri, S.; Bouacem, K.; Chalbi, T.B.; Khaled, M.; Allala, F.; Bouanane-Darenfed, A.; Hacene, H.; Jaouadi, B. A Taguchi design approach for the enhancement of a detergent-biocompatible alkaline thermostable protease production by Streptomyces mutabilis strain TN-X30. J. Surfactants Deterg. 2022, 25, 478–504. [Google Scholar] [CrossRef]
- Abdullah, R.; Qaiser, H.; Iftikhar, T.; Kaleem, A.; Iqtedar, M.; Saleem, F.; Haq, I. Application of response surface methodology for statistical optimization of lipase production by Penicillium sp. employing solid state fermentation. Rev. Mex. Ing. Química 2018, 17, 863–875. [Google Scholar]
- Tiss, A.; Carrière, F.; Verger, R. Effects of gum arabic on lipase interfacial binding and activity. Anal. Biochem. 2001, 294, 36–43. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Laane, C.; Boeren, S.; Vos, K.; Veeger, C. Rules for optimization of biocatalysis in organic solvents. Biotechnol. Bioeng. 1987, 30, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Saraswat, R.; Verma, V.; Sistla, S.; Bhushan, I. Evaluation of alkali and thermotolerant lipase from an indigenous isolated Bacillus strain for detergent formulation. Electron. J. Biotechnol. 2017, 30, 33–38. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Steps a | Total Activity (U) | Total Protein (mg) | Specific Activity (U/mg) | Purification (-Fold) | Yield (%) |
|---|---|---|---|---|---|
| Crude extract | 7680 ± 21 | 48.0 ± 1 | 160 ± 2.5 | 1 | 100 |
| (NH4)2SO4 precipitation (70%) | 3900 ± 10 | 15.3 ± 0.5 | 254 ± 4.0 | 1.6 | 51 |
| HiTrap Q-Sepharose FF column | 2600 ± 6 | 0.2 ± 0 | 10,000 ± 5 | 62.5 | 34 |
| Origin | NH2-Terminal Amino Acid a | Identity (%) |
|---|---|---|
| Penicillium crustosum strain P22 (this study) | ATADAAAFLDLHMAAKLSSA b | - |
| Penicillium cyclopium strain PG37 (AF274320) | ATADAAAFPDLHRAAKLSSA | 90 |
| Penicillium camemberti strain FM013 (CRL22273) | ATAATAAFPDLHRAAKLSSA | 80 |
| Penicillium solitum strain IBT 29525 (XP_040813639) | ATAATAAFPDLNXAAKLSSA | 75 |
| Penicillium expansum strain DSM 1994 (AAB27002) | AVAASAAFPDLXRAAKLSSA | 70 |
| Inhibitors/Chemical Reagents/Metal Ions a | Concentration a | Residual Lipase Activity (%) b |
|---|---|---|
| None | - | 100 b ± 3 |
| PMSF | 5 mM | 0 |
| DFP | 2 mM | 0 |
| Benzamidine | 2 mM | 110 b ± 3 |
| DTNB | 10 mM | 73 bc ± 2 |
| NEM | 2 mM | 95 b ± 2 |
| Iodoacetamide | 5 mM | 106 b ± 3 |
| PAO | 10 mM | 94 b ± 2 |
| β-ME | 10 mM | 96 b ± 2 |
| dl-DTT | 10 mM | 98 b ± 2 |
| EDTA | 10 mM | 92 b ± 2 |
| EGTA | 1 mM | 96 b ± 2 |
| Ca2+ (CaCl2) | 2 mM | 147 a ± 4 |
| Mn2+ (MnCl2) | 2 mM | 146 a ± 4 |
| Mg2+ (MgCl2) | 2 mM | 92 b ± 2 |
| Fe2+ (FeSO4) | 2 mM | 70 bc ± 2 |
| Co2+ (CoCl2) | 2 mM | 41 cd ± 1 |
| Ba2+ (BaCl2) | 2 mM | 96 b ± 2 |
| Zn2+ (ZnSO4) | 2 mM | 9 d ± 0 |
| Cu2+ (CuCl2) | 2 mM | 23 d ± 1 |
| Ni2+ (NiCl2) | 2 mM | 16 d ± 0 |
| Hg2+ (HgCl2) | 2 mM | 0 |
| Cd2+ (CdCl2) | 2 mM | 0 |
| Detergent Additives | Final Concentration | Residual Lipase Activity (%) a,b | ||
|---|---|---|---|---|
| PCrL | Palatase® 20000 L | Lipolase® | ||
| Control | 0 | 100 cd ± 3 | 100 cd ± 3 | 100 cd ± 3 |
| Zeolite | 1% (w/v) | 95 cd ± 2 | 91 cd ± 2 | 86 cd ± 2 |
| STPP | 0.2% (w/v) | 150 ab ± 4 | 146 ab ± 4 | 15 f ± 4 |
| 0.5% (w/v) | 98 cd ± 2 | 87 cd ± 2 | 97 cd ± 2 | |
| SDS | 1% (w/v) | 185 a ± 4 | 90 cd ± 2 | 96 cd ± 2 |
| 2% (w/v) | 150 ab ± 4 | 66 de ± 2 | 75 d ± 2 | |
| Na2CO3 | 50 mM | 171 a ± 4 | 165 a ± 4 | 154 ab ± 4 |
| 100 mM | 147 ab ± 4 | 141 ab ± 4 | 126 bc ± 3 | |
| Tween 20 | 1% (v/v) | 96 cd ± 2 | 87 cd ± 2 | 73 d ± 2 |
| 5% (v/v) | 49 de ± 1 | 76 d ± 2 | 54 de ± 1 | |
| Tween 40 | 1% (v/v) | 108 bc ± 3 | 98 cd ± 2 | 87 cd ± 2 |
| 5% (v/v) | 45 e ± 1 | 43 e ± 1 | 36 ef ± 1 | |
| Tween 60 | 1% (v/v) | 84 cd ± 2 | 95 cd ± 2 | 98 cd ± 2 |
| 5% (v/v) | 31 ef ± 1 | 67 de ± 2 | 60 de ± 2 | |
| Tween 80 | 1% (v/v) | 83 cd ± 2 | 93 cd ± 2 | 90 cd ± 2 |
| 5% (v/v) | 29 ef ± 1 | 52 de ± 1 | 50 de ± 1 | |
| Na2CMC | 1% (w/v) | 143 ab ± 4 | 96 cd ± 2 | 92 cd ± 2 |
| 10% (w/v) | 105 c ± 3 | 64 de ± 2 | 52 de ± 1 | |
| Triton X-100 | 1% (v/v) | 140 ab ± 4 | 80 cd ± 2 | 85 cd ± 2 |
| 5% (v/v) | 103 cd ± 3 | 66 de ± 2 | 68 de ± 2 | |
| TAED | 0.5% (w/v) | 142 ab ± 4 | 126 bc ± 3 | 116 bc ± 3 |
| 5% (w/v) | 103 cd ± 3 | 92 cd ± 2 | 75 d ± 2 | |
| Sodium perborate | 1% (v/v) | 139 ab ± 3 | 80 b ± 2 | 113 bc ± 3 |
| 5% (v/v) | 99 cd ± 3 | 51 de ± 1 | 88 cd ± 2 | |
| H2O2 | 5% (v/v) | 146 abA ± 4 | 61 deA ± 2 | 80 cdA ± 2 |
| 10% (v/v) | 104 cdA ± 3 | 45 eA ± 1 | 66 deA ± 2 | |
| Commercialized Detergent Additives | Concentration (%, v/v) | Residual Lipase Activity (%) a,b | ||
|---|---|---|---|---|
| PCrL | Palatase® 20000 L | Lipolase® | ||
| Control | 0 | 100 cd ± 3 | 100 cd ± 3 | 100 cd ± 3 |
| SAFOL 23 E7 | 0.5 | 152 ab ± 4 | 145 ab ± 4 | 142 ab ± 4 |
| 1 | 95 cd ± 2.4 | 93 cd ± 2 | 90 cd ± 2 | |
| Dehydol® LT 7 | 0.5 | 154 ab ± 4 | 142 ab ± 4 | 164 a ± 4 |
| 1 | 99 cd ± 2 | 95 cd ± 2 | 108 bc ± 3 | |
| SURFAC® LM 30 | 0.5 | 183 a ± 5 | 142 ab ± 4 | 120 bc ± 3 |
| 1 | 150 ab ± 4 | 102 cd ± 3 | 94 cd ± 2 | |
| NEODOL® 25-7 | 0.5 | 75 d ± 2 | 96 cd ± 2 | 87 cd ± 2 |
| 1 | 25 ef ± 1 | 57 de ± 1 | 45 e ± 1 | |
| Galaxy LAS | 2 | 82 cd ± 2 | 99 cd ± 2 | 95 cd ± 2 |
| 5 | 30 ef ± 1 | 60 de ± 2 | 59 de ± 1 | |
| Galaxy LES 70 | 2 | 110 bc ± 3 | 103 cd ± 3 | 94 cd ± 2 |
| 5 | 55 de ± 1 | 53 de ± 1 | 50 de ± 1 | |
| Galaxy 110 | 2 | 149 ab ± 4 | 150 ab ± 4 | 125 cb ± 3 |
| 5 | 93 cd ± 2 | 98 cd ± 2 | 83 cd ± 2 | |
| Galaxy CAPB Plus | 1 | 153 ab ± 4 | 162 a ± 4 | 14 f ± 4 |
| 5 | 105 c ± 3 | 106 b ± 3 | 105 c ± 4 | |
| TERGITOLTM NP-9 SURFACTANT | 2 | 86 cd ± 2 | 76 d ± 2 | 89 cd ± 2 |
| 5 | 30 ef ± 1 | 46 e ± 1 | 50 de ± 1 | |
| FINDET® AR/52 | 0.5 | 78 d ± 2 | 82 cd ± 2 | 75 d ± 2 |
| 1 | 26 ef ± 1 | 35 ef ± 1 | 28 ef ± 1 | |
| Anti-foam | 0.5 | 85 cd ± 2 | 98 cd ± 2 | 88 cd ± 2 |
| 1 | 46 e ± 1 | 56 de ± 1 | 50 ed ± 1 | |
| Formol | 0.2 | 160 ab ± 4 | 153 ab ± 4 | 135 a ± 3 |
| 0.5 | 108 bc ± 3 | 96 cd ± 2 | 84 cd ± 2 | |
| Tinopal® CBS-X | 0.5 | 152 ab ±4 | 164 a ± 4 | 122 bc ± 3 |
| 1 | 95 cd ± 2 | 111 bc ± 3 | 86 cd ± 2 | |
| Sulfacid K | 10 | 77 d ± 2 | 84 cd ± 2 | 83 cd ± 2 |
| 15 | 15 f ± 1 | 32 ef ± 1 | 25 ef ± 1 | |
| Marlipal® 31/90 | 0.5 | 79 cd ± 2 | 66 de ± 2 | 83 cd ± 2 |
| 1 | 30 ef ± 1 | 42 ef ± 1 | 35 ef ± 1 | |
| EDTA | 0.2 | 130 b ± 3 | 111 bc ± 3 | 125 bc ± 3 |
| 0.5 | 76 d ± 2 | 71 de ± 2 | 62 de ± 2 | |
| Perfume I Class | 0.5 | 165 ab ± 4 | 152 ab ± 4 | 146 ab ± 4 |
| 1 | 109 bc ± 3 | 101 b ± 3 | 99 cd ± 2 | |
| Perfume II Class | 0.5 | 170 a ± 4 | 162 a ± 4 | 155 ab ± 4 |
| 1 | 110 bc ± 3 | 105 c ± 3 | 96 cd ± 2 | |
| Propyl betaine | 1 | 95 cd ± 2 | 85 cd ± 2 | 81 cd ± 2 |
| NaOH 50% | 1 | 113 bc ± 3 | 122 bc ± 3 | 119 bc ± 3 |
| Laundry Detergent (7 mg/mL) | Oil Removal (%) | |||
|---|---|---|---|---|
| Detergent | Detergent + PCrL | Detergent + Palatase® 20000 L | Detergent + Lipolase® | |
| Ariel | 54 bc ± 2 | 100 a ± 3 | 93 a ± 2 | 98 a ± 3 |
| OMO | 40 c ± 1 | 81 a ± 2 | 88 a ± 2 | 100 a ± 3 |
| Nadhif | 34 c ± 1 | 51 bc ± 2 | 45 bc ± 1 | 42 c ± 1 |
| EcoVax | 35 c ± 1 | 72 b ± 1 | 53 bc ± 2 | 61 b ± 1 |
| Dipex | 27 cd ± 1 | 54 bc ± 1 | 59 b ± 2 | 68 b ± 1 |
| Maison Det | 44 c ± 1 | 91 a ± 2 | 76 ab ± 1 | 82 a ± 2 |
| Pro-Clean | 55 bc ± 1 | 100 a ± 3 | 100 a ± 3 | 87 a ± 2 |
| New Det | 29 c ± 1 | 61 b ± 1 | 41 c ± 1 | 39 c ± 1 |
| New Dex | 39 c ± 1 | 98 a ± 3 | 91 a ± 2 | 86 a ± 2 |
| Class | 49 bc ± 2 | 96 a ± 2 | 89 a ± 2 | 86 a ± 2 |
| Fino | 21 d ± 2 | 46 bc ± 1 | 50 cb ± 2 | 39 c ± 1 |
| X-Clean | 46 bc ± 1 | 90 a ± 2 | 100 a ± 3 | 98 a ± 3 |
| Dixan | 21 d ± 1 | 54 bc ± 1 | 45 cb ± 1 | 59 b ± 2 |
| Skip | 25 cd ± 1 | 65 b ± 1 | 57 b ± 1 | 62 b ± 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasnaoui, I.; Dab, A.; Mechri, S.; Abouloifa, H.; Saalaoui, E.; Jaouadi, B.; Noiriel, A.; Asehraou, A.; Abousalham, A. Purification, Biochemical and Kinetic Characterization of a Novel Alkaline sn-1,3-Regioselective Triacylglycerol Lipase from Penicilliumcrustosum Thom Strain P22 Isolated from Moroccan Olive Mill Wastewater. Int. J. Mol. Sci. 2022, 23, 11920. https://doi.org/10.3390/ijms231911920
Hasnaoui I, Dab A, Mechri S, Abouloifa H, Saalaoui E, Jaouadi B, Noiriel A, Asehraou A, Abousalham A. Purification, Biochemical and Kinetic Characterization of a Novel Alkaline sn-1,3-Regioselective Triacylglycerol Lipase from Penicilliumcrustosum Thom Strain P22 Isolated from Moroccan Olive Mill Wastewater. International Journal of Molecular Sciences. 2022; 23(19):11920. https://doi.org/10.3390/ijms231911920
Chicago/Turabian StyleHasnaoui, Ismail, Ahlem Dab, Sondes Mechri, Houssam Abouloifa, Ennouamane Saalaoui, Bassem Jaouadi, Alexandre Noiriel, Abdeslam Asehraou, and Abdelkarim Abousalham. 2022. "Purification, Biochemical and Kinetic Characterization of a Novel Alkaline sn-1,3-Regioselective Triacylglycerol Lipase from Penicilliumcrustosum Thom Strain P22 Isolated from Moroccan Olive Mill Wastewater" International Journal of Molecular Sciences 23, no. 19: 11920. https://doi.org/10.3390/ijms231911920