
Genetic Analysis and Fine Mapping of ZmGHT1 Conferring Glufosinate Herbicide Tolerance in Maize (Zea mays L.)
 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Isolation and Response of L336R to Glufosinate
2.2. Inheritance of ZmGHT1
2.3. Genetic Mapping of ZmGHT1
2.4. Candidate Gene Prediction, Functional Annotation, and Expression Analysis
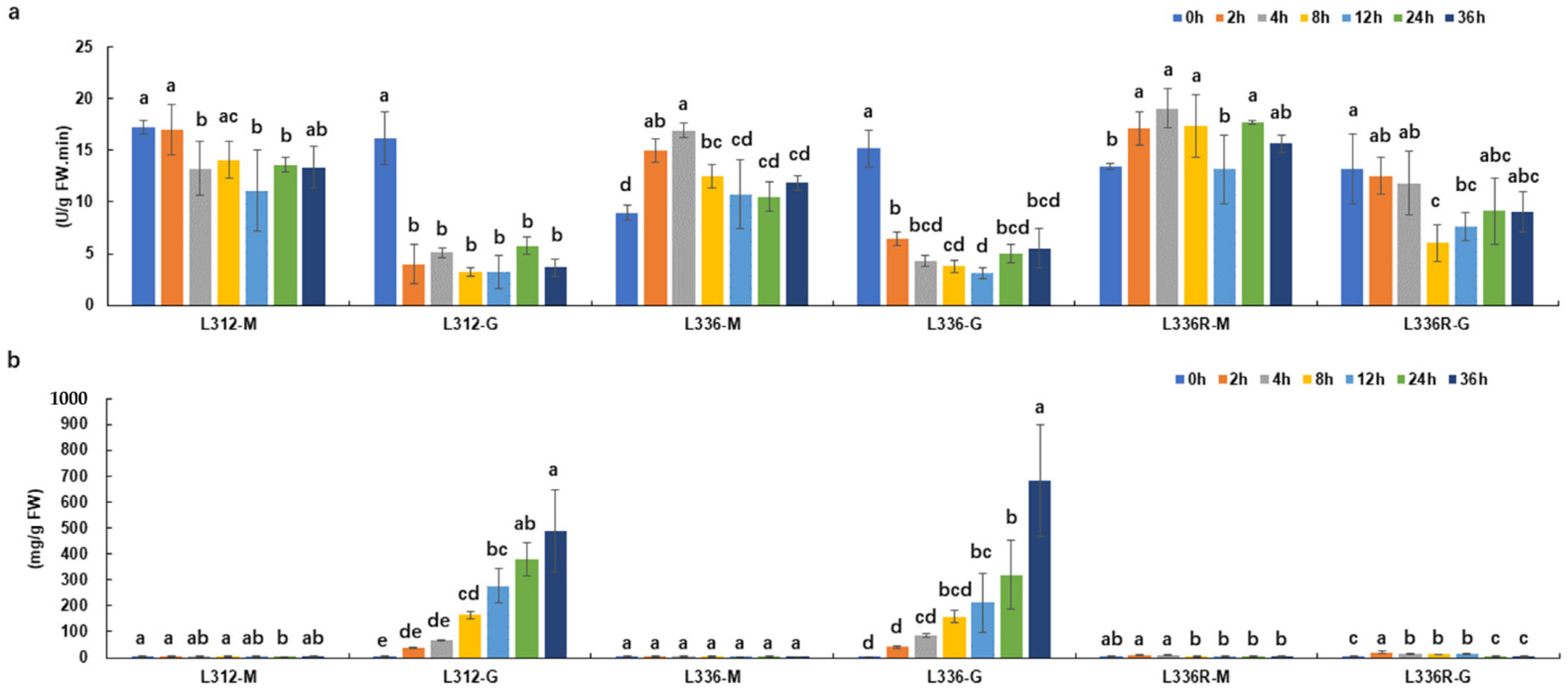
2.5. GS Activity Levels and Ammonia Content Determination during Glufosinate Treatment
2.6. Detection Efficiency Test of M9 and M10 for Identifying Functional Allele of ZmGHT1
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Phenotyping
4.2. Genetic Mapping Population Construction and Fine Mapping of ZmGHT1
4.3. Molecular Marker Development
4.4. Extraction of Genomic DNA and PCR Conditions
4.5. Sequence Analysis
4.6. RNA Extraction and Gene Expression Analysis
4.7. Ammonia Accumulation Measurement
4.8. GS Activity Determination
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oerke, E.C. Crop losses to pests. J. Agri. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Holm, L.G.; Plucknett, D.L.; Pancho, J.V.; Herberger, J.P. The World Worsts Weeds. Distribution and Biology; University Press of Hawaii: Honolulu, HI, USA, 1977. [Google Scholar]
- Li, Y.H.; Yang, R.J.; Yang, B.Z.; Wu, W.C.; Zhang, L.Y.; Qiang, S.; Islam, F.; Yang, C.; Chen, X.; Zhou, W. Weed Flora in China; China Agriculture Press: Beijing, China, 1998. [Google Scholar]
- Zhu, J.; Wang, J.; Ditommaso, A.; Zhang, C.; Zheng, G.; Liang, W.; Islam, F.; Yang, C.; Chen, X.; Zhou, W. Weed research status, challenges, and opportunities in china. Crop Prot. 2020, 134, 104449. [Google Scholar] [CrossRef]
- Soltani, N.; Dille, J.A.; Burke, I.C.; Everman, W.J.; Van Gessel, M.J.; Davis, V.M.; Sikkema, P.H. Potential corn yield losses from weeds in North America. Weed Technol. 2016, 30, 979–984. [Google Scholar] [CrossRef]
- Clay, S.A. Near term challenges for global agriculture-herbicide resistant weeds. Agron J. 2021, 113, 4463–4472. [Google Scholar] [CrossRef]
- Liu, X.; Merchant, A.; Xiang, S.; Zong, T.; Zhou, X.; Bai, L. Managing herbicide resistance in China. Weed Sci. 2021, 69, 4–17. [Google Scholar] [CrossRef]
- Heap, I. The International Herbicide-Resistant Weed Database. Available online: www.weedscience.org (accessed on 12 August 2022).
- Green, J.M. Current state of herbicides in herbicide-resistant crops. Pest Manag. Sci. 2014, 70, 1351–1357. [Google Scholar] [CrossRef]
- Gaines, T.A.; Busi, R.; Küpper, A. Can new herbicide discovery allow weed management to outpace resistance evolution? Pest Manag. Sci. 2021, 77, 3036–3041. [Google Scholar] [CrossRef]
- Gaines, T.A.; Duke, S.O.; Morran, S.; Rigon, C.A.G.; Tranel, P.J.; Küpper, A.; Dayan, F.E. Mechanisms of evolved herbicide resistance. J. Biol. Chem. 2020, 295, 10307–10330. [Google Scholar] [CrossRef]
- Hall, C.J.; Mackie, E.R.; Gendall, A.R.; Perugini, M.A.; Soares da Costa, T.P. Review: Amino acid biosynthesis as a target for herbicide development. Pest. Manag. Sci. 2020, 76, 3896–3904. [Google Scholar] [CrossRef]
- Dill, G.M. Glyphosate-resistant crops: History, status and future, Pest Manag. Sci. 2005, 6, 1219–1224. [Google Scholar]
- Sammons, D.R.; Gaines, T.A. Glyphosate resistance: State of knowledge. Pest Manag. Sci. 2014, 70, 1367–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidhu, R.S.; Hammond, B.G.; Fuchs, R.L.; Mutz, J.N.; Holden, L.R.; George, B.; Olson, T. Glyphosate-tolerant corn: The composition and feeding value of grain from glyphosate-tolerant corn is equivalent to that of conventional corn (Zea mays L.). J. Agric. Food Chem. 2000, 48, 2305–2312. [Google Scholar] [CrossRef] [PubMed]
- Funke, T.; Yang, Y.; Han, H.; Healy-Fried, M.; Olesen, S.; Becker, A.; Schönbrunn, E. Structural basis of glyphosate resistance resulting from the double mutation Thr97 → Ile and Pro101 → Ser in 5-enolpyruvylshikimate-3-phosphate synthase from Escherichia coli. J. Biol. Chem. 2009, 284, 9854–9860. [Google Scholar] [CrossRef] [PubMed]
- Kishore, G.M.; Shah, D.M. Amino acid biosynthesis inhibitors as herbicides. Annu. Rev. Biochem. 1988, 57, 627–663. [Google Scholar] [CrossRef] [PubMed]
- Takano, H.K.; Dayan, F.E. Glufosinate-ammonium: A review of the current state of knowledge. Pest Manag. Sci. 2020, 76, 3911–3925. [Google Scholar] [CrossRef]
- Hess, F.D. Light-dependent herbicides: An overview. Weed Sci. 2000, 48, 160–170. [Google Scholar] [CrossRef]
- Unno, H.; Uchida, T.; Sugawara, H.; Kurisu, G.; Sugiyama, T.; Yamaya, T.; Sakakibara, H.; Hase, T.; Kusunoki, M. Atomic structure of plant glutamine synthetase: A key enzyme for plant productivity. J. Biol. Chem. 2006, 281, 29287–29296. [Google Scholar] [CrossRef]
- Avila-Garcia, W.V.; Sanchez-Olguin, E.; Hulting, A.G.; Mallory-Smith, C. Target-site mutation associated with glufosinate resistance in Italian ryegrass (Lolium perenne L. ssp. multiflorum). Pest Manag. Sci. 2012, 68, 1248–1254. [Google Scholar] [CrossRef]
- Zhang, C.; Yu, Q.; Han, H.; Yu, C.; Nyporko, A.; Tian, X.; Beckie, H.; Powles, S. A naturally evolved mutation (Ser59Gly) in glutamine synthetase confers glufosinate resistance in plants. J. Exp. Bot. 2022, 73, 2251–2262. [Google Scholar] [CrossRef]
- Sharma, P.; Singh, S.P.; Iqbal, H.M.N.; Parra-Saldivar, R.; Varjani, S.; Tong, Y.W. Genetic modifications associated with sustainability aspects for sustainable developments. Bioengineered 2022, 13, 9508–9520. [Google Scholar] [CrossRef]
- Kuang, Y.; Li, S.; Ren, B.; Yan, F.; Spetz, C.; Li, X.; Zhou, X.; Zhou, H. Base-editing-mediated artificial evolution of OsALS1 in planta to develop novel herbicide-tolerant rice germplasms. Mol. Plant 2020, 13, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Murata, K.; Sakuma, N.; Takei, S.; Yamazaki, A.; Karim, M.R.; Kawata, M.; Hirose, S.; Kawagishi-Kobayashi, M.; Taniguchi, Y.; et al. A rice gene that confers broad-spectrum resistance to β-triketone herbicides. Science 2019, 365, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhang, X.; Wu, C.; He, Y.; Ma, Y.; Hou, H.; Guo, X.; Du, W.; Zhao, Y.; Xia, L. Engineering herbicide-resistant rice plants through CRISPR/Cas9-mediated homologous recombination of acetolactate synthase. Mol. Plant 2016, 9, 628–631. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.Y.; Chai, Y.P.; Lu, M.H.; Han, X.L.; Lin, Q.; Zhang, Y.; Zhang, Q.; Zhou, Y.; Wang, X.C.; Gao, C.; et al. Prime editing efficiently generates W542L and S621I double mutations in two ALS genes in maize. Genome Biol. 2020, 21, 257. [Google Scholar] [CrossRef]
- Qin, W.; Bao, J.; Wang, Y.; Ma, Y.; Long, Y.; Li, J.; Dong, Z.; Wan, X. Genome-wide association study of leaf angle traits and mining of elite alleles from the major loci in maize. Acta Agron. Sin. 2022, 48, 2691–2705. [Google Scholar] [CrossRef]
- Anderson, S.N.; Stitzer, M.C.; Brohammer, A.B.; Zhou, P.; Noshay, J.M.; O’Connor, C.H.; Hirsch, C.D.; Ross-Ibarra, J.; Hirsch, C.N.; Springer, N.M. Transposable elements contribute to dynamic genome content in maize. Plant J. 2019, 100, 1052–1065. [Google Scholar] [CrossRef]
- Qiu, Y.; O’Connor, C.H.; Della Coletta, R.; Renk, J.S.; Monnahan, P.J.; Noshay, J.M.; Liang, Z.; Gilbert, A.; Anderson, S.N.; McGaugh, S.E.; et al. Whole-genome variation of transposable element insertions in a maize diversity panel. G3 (Bethesda) 2021, 11, jkab238. [Google Scholar] [CrossRef]
- Souvorov, A.; Kapustin, Y.; Kiryutin, B.; Chetvernin, V.; Tatusova, T.; Lipman, D. Gnomon-NCBI Eukaryotic Gene Prediction Tool; National Center for Biotechnology Information: Bethesda, MD, USA, 2010. [Google Scholar]
- Swarbreck, S.M.; Defoin-Platel, M.; Hindle, M.; Saqi, M.; Habash, D.Z. New perspectives on glutamine synthetase in grasses. J. Exp. Bot. 2011, 62, 1511–1522. [Google Scholar] [CrossRef]
- Prinsi, B.; Espen, L. Mineral nitrogen sources differently affect root glutamine synthetase isoforms and amino acid balance among organs in maize. BMC Plant Biol. 2015, 15, 96. [Google Scholar] [CrossRef]
- He, Y.; Wang, M.; Dukowic-Schulze, S.; Zhou, A.; Tiang, C.L.; Shilo, S.; Sidhu, G.K.; Eichten, S.; Bradbury, P.; Springer, N.M.; et al. Genomic features shaping the landscape of meiotic double-strand-break hotspots in maize. Proc. Natl. Acad. Sci. USA 2017, 114, 12231–12236. [Google Scholar] [CrossRef]
- Luo, C.; Li, X.; Zhang, Q.; Yan, J. Single gametophyte sequencing reveals that crossover events differ between sexes in maize. Nat. Commun. 2019, 10, 785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Li, W.; Wang, L.; Yan, J.; Lu, G.; Yang, N.; Xu, J.; Wang, Y.; Gui, S.; Chen, G.; et al. Three types of genes underlying the Gametophyte factor1 locus cause unilateral cross incompatibility in maize. Nat. Commun. 2022, 13, 4498. [Google Scholar] [CrossRef] [PubMed]
- Boideau, F.; Richard, G.; Coriton, O.; Huteau, V.; Belser, C.; Deniot, G.; Eber, F.; Falentin, C.; Ferreira de Carvalho, J.; Gilet, M.; et al. Epigenomic and structural events preclude recombination in Brassica napus. New Phytol. 2022, 234, 545–559. [Google Scholar] [CrossRef] [PubMed]
- Concepcion, J.C.T.; Kaundun, S.S.; Morris, J.A.; Hutchings, S.J.; Strom, S.A.; Lygin, A.V.; Riechers, D.E. Resistance to a nonselective 4-hydroxyphenylpyruvate dioxygenase-inhibiting herbicide via novel reduction-dehydration-glutathione conjugation in Amaranthus tuberculatus. New Phytol. 2021, 232, 2089–2105. [Google Scholar] [CrossRef]
- Huang, X.X.; Zhao, S.M.; Zhang, Y.Y.; Li, Y.J.; Shen, H.N.; Li, X.; Hou, B.K. A novel UDP-glycosyltransferase 91C1 confers specific herbicide resistance through detoxification reaction in Arabidopsis. Plant Physiol. Biochem. 2021, 159, 226–233. [Google Scholar] [CrossRef]
- Lv, Z.; Zhao, M.; Wang, W.; Wang, Q.; Huang, M.; Li, C.; Lian, Q.; Xia, J.; Qi, J.; Xiang, C.; et al. Changing Gly311 to an acidic amino acid in the MATE family protein DTX6 enhances Arabidopsis resistance to the dihydropyridine herbicides. Mol. Plant 2021, 14, 2115–2125. [Google Scholar] [CrossRef]
- Pan, L.; Yu, Q.; Wang, J.; Han, H.; Mao, L.; Nyporko, A.; Maguza, A.; Fan, L.; Bai, L.; Powles, S. An ABCC-type transporter endowing glyphosate resistance in plants. Proc. Natl. Acad. Sci. USA 2021, 118, e2100136118. [Google Scholar] [CrossRef]
- Xu, R.; Liu, X.; Li, J.; Qin, R.; Wei, P. Identification of herbicide resistance OsACC1 mutations via in planta prime-editing-library screening in rice. Nat. Plants 2021, 7, 888–892. [Google Scholar] [CrossRef]
- Shen, J.; Yang, Q.; Hao, L.; Zhang, L.; Li, X.; Zheng, M. The Metabolism of a Novel Cytochrome P450 (CYP77B34) in Tribenuron-Methyl-Resistant Descurainia sophia L. to Herbicides with Different Mode of Actions. Int. J. Mol. Sci. 2022, 23, 5812. [Google Scholar] [CrossRef]
- Garcia, M.D.; Nouwens, A.; Lonhienne, T.G.; Guddat, L.W. Comprehensive understanding of acetohydroxyacid synthase inhibition by different herbicide families. Proc. Natl. Acad. Sci. USA 2017, 114, E1091–E1100. [Google Scholar] [CrossRef]
- Tian, H.; Yang, Y.; Yi, H.; Xu, L.; He, H.; Fan, Y.; Wang, L.; Ge, J.; Liu, Y.; Wang, F.; et al. New resources for genetic studies in maize (Zea mays L.): A genome-wide Maize6H-60K single nucleotide polymorphism array and its application. Plant J. 2021, 105, 1113–1122. [Google Scholar] [CrossRef] [PubMed]
- da Maia, L.C.; Palmieri, D.A.; de Souza, V.Q.; Kopp, M.M.; de Carvalho, F.I.; Costa de Oliveira, A. SSR locator: Tool for simple sequence repeat discovery integrated with primer design and PCR simulation. Int. J. Plant Genom. 2008, 2008, 412696. [Google Scholar] [CrossRef] [PubMed]
- Saghai-Maroof, M.A.; Soliman, K.M.; Jorgenson, R.A.; Allard, R.W. Ribosomal DNA spacer-length polymorphisms in barley: Mendelian inheritance, chromosomal location, and population dynamics. Proc. Natl. Acad. Sci. USA 1984, 81, 8014–8018. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.; Gish, W.; Miller, W.; Myers, E.; Lipman, D. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Campanella, J.J.; Bitincka, L.; Smalley, J. MatGAT: An application that generates similarity/identity matrices using protein or DNA sequences. BMC Bioinform. 2003, 4, 29. [Google Scholar] [CrossRef]
- Wintersinger, J.A.; Wasmuth, J.D. Kablammo: An interactive, web-based BLAST results visualizer. Bioinformatics 2015, 31, 1305–1306. [Google Scholar] [CrossRef]
- An, X.; Ma, B.; Duan, M.; Dong, Z.; Liu, R.; Yuan, D.; Hou, Q.; Wu, S.; Zhang, D.; Liu, D.; et al. Molecular regulation of ZmMs7 required for maize male fertility and development of a dominant male-sterility system in multiple species. Proc. Natl. Acad. Sci. USA 2020, 117, 23499–23509. [Google Scholar] [CrossRef]
- Weatherburn, M.W. Phenol hipochlorite reaction for determination of ammonia. Anal. Chem. 1967, 39, 971–974. [Google Scholar] [CrossRef]
- Ding, F.; Hu, Q.; Wang, M.; Zhang, S. Knockout of SlSBPASE suppresses carbon sssimilation and slters snitrogen metabolism in tomato plants. Int. J. Mol. Sci. 2018, 19, 4046. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| F2:3 Family | Observed Frequency | Expected Frequency | χ2 |
|---|---|---|---|
| Homozygous resistance | 47 | 48 | 0.198 |
| Heterozygous resistance | 99 | 96 | |
| Susceptible | 46 | 48 |
| Marker | Forward Primer Sequence | Reverse Primer Sequence |
|---|---|---|
| M1 | CTGGAAAAGGAGGACTACTG | AAAGCAAGAAGTTGATATAGCC |
| M2 | GGGACGAGTGCCAAATCAG | GGCCTTCGCAAAGAACCAA |
| M3 | CAATCTATCGCTTGACTCATC | GCTACTACTAACTTGCCCAAAA |
| M4 | CATACTATGTCAAAATGCATCC | GAAAACGATGATAAATGGAACT |
| M5 | TTTCCCACAATACCAACT | GCTAGCTCTTCTGCAATC |
| M6 | AGCCTTGTCTTTGTGGTG | GAACAGTCTGCGACTTGC |
| M7 | CTTCCTTTAACGCCACAA | CGGCACAAGTGTTTCATCA |
| M8 | GGAGATTAAGAGCTGCAA | ATGTTCTGGAAAGTGGTT |
| M9 | TAGACAAGAAACCAAACAT | CAAAAGGTAAGGTGAGAC |
| M10 | CCAACACGTGGCAGGCAG | CGCCGAGTCACCAATCCAC |
| M11 | TATGAGTAAGTAGCGTGAAGCA | CCTATCTATTGGAAGCCTATGA |
| Gene ID | Start | End | Strand | Functional Annotation |
|---|---|---|---|---|
| Zm00001eb361900 | 160124107 | 160128959 | + | Epstein–Barr nuclear antigen |
| gene.8455045 | 160775402 | 160775752 | − | No significant similarity |
| gene.8455061 | 160782742 | 160783140 | − | No significant similarity |
| gene.8455077 | 160783139 | 160783441 | + | No significant similarity |
| gene.8455093 | 161011640 | 161011329 | + | No significant similarity |
| gene.8455109 | 161124578 | 161128321 | + | Uncharacterized protein |
| gene.8455125 | 161128385 | 161129846 | − | Embryogenesis transmembrane protein-like |
| Zm00001eb361920 | 161130057 | 161130464 | − | Embryogenesis transmembrane protein-like |
| gene.3251669862 | 161493564 | 161495449 | − | Retrovirus-related Pol polyprotein LINE-1 |
| gene.3251669894 | 161495561 | 161496568 | − | SH3 domain-containing protein 2 |
| gene.3251669926 | 162029341 | 162030174 | − | class III aminotransferase |
| Zm00001eb361930 | 162030458 | 162031088 | − | class III aminotransferase |
| gene.8455157 | 162031992 | 162033874 | − | Cationic amino acid transporter 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bao, J.; Gao, Y.; Li, Y.; Wu, S.; Li, J.; Dong, Z.; Wan, X. Genetic Analysis and Fine Mapping of ZmGHT1 Conferring Glufosinate Herbicide Tolerance in Maize (Zea mays L.). Int. J. Mol. Sci. 2022, 23, 11481. https://doi.org/10.3390/ijms231911481
Bao J, Gao Y, Li Y, Wu S, Li J, Dong Z, Wan X. Genetic Analysis and Fine Mapping of ZmGHT1 Conferring Glufosinate Herbicide Tolerance in Maize (Zea mays L.). International Journal of Molecular Sciences. 2022; 23(19):11481. https://doi.org/10.3390/ijms231911481
Chicago/Turabian StyleBao, Jianxi, Yuexin Gao, Yanan Li, Suowei Wu, Jinping Li, Zhenying Dong, and Xiangyuan Wan. 2022. "Genetic Analysis and Fine Mapping of ZmGHT1 Conferring Glufosinate Herbicide Tolerance in Maize (Zea mays L.)" International Journal of Molecular Sciences 23, no. 19: 11481. https://doi.org/10.3390/ijms231911481