
A Combined Cell-Worm Approach to Search for Compounds Counteracting the Toxicity of Tau Oligomers In Vivo
 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
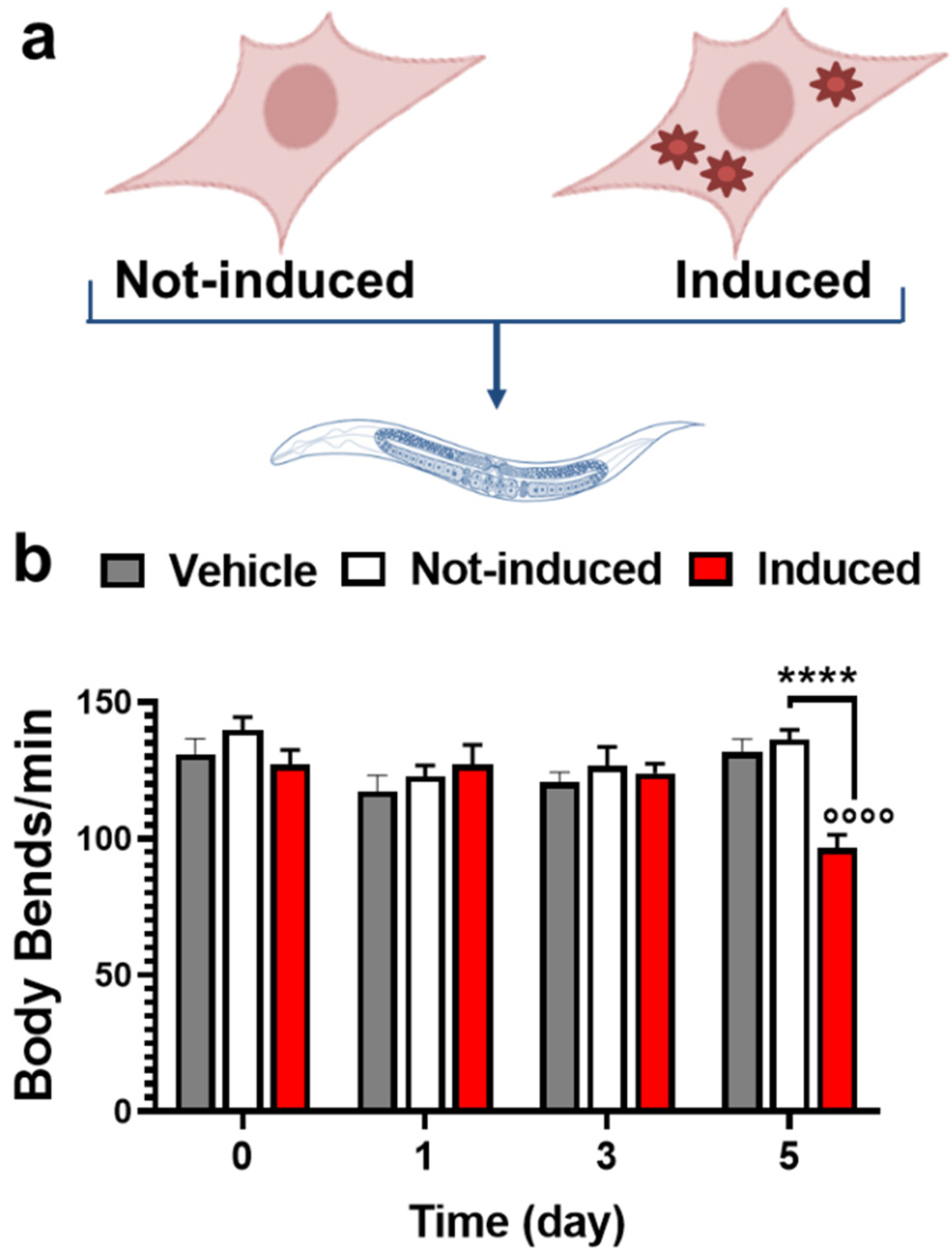
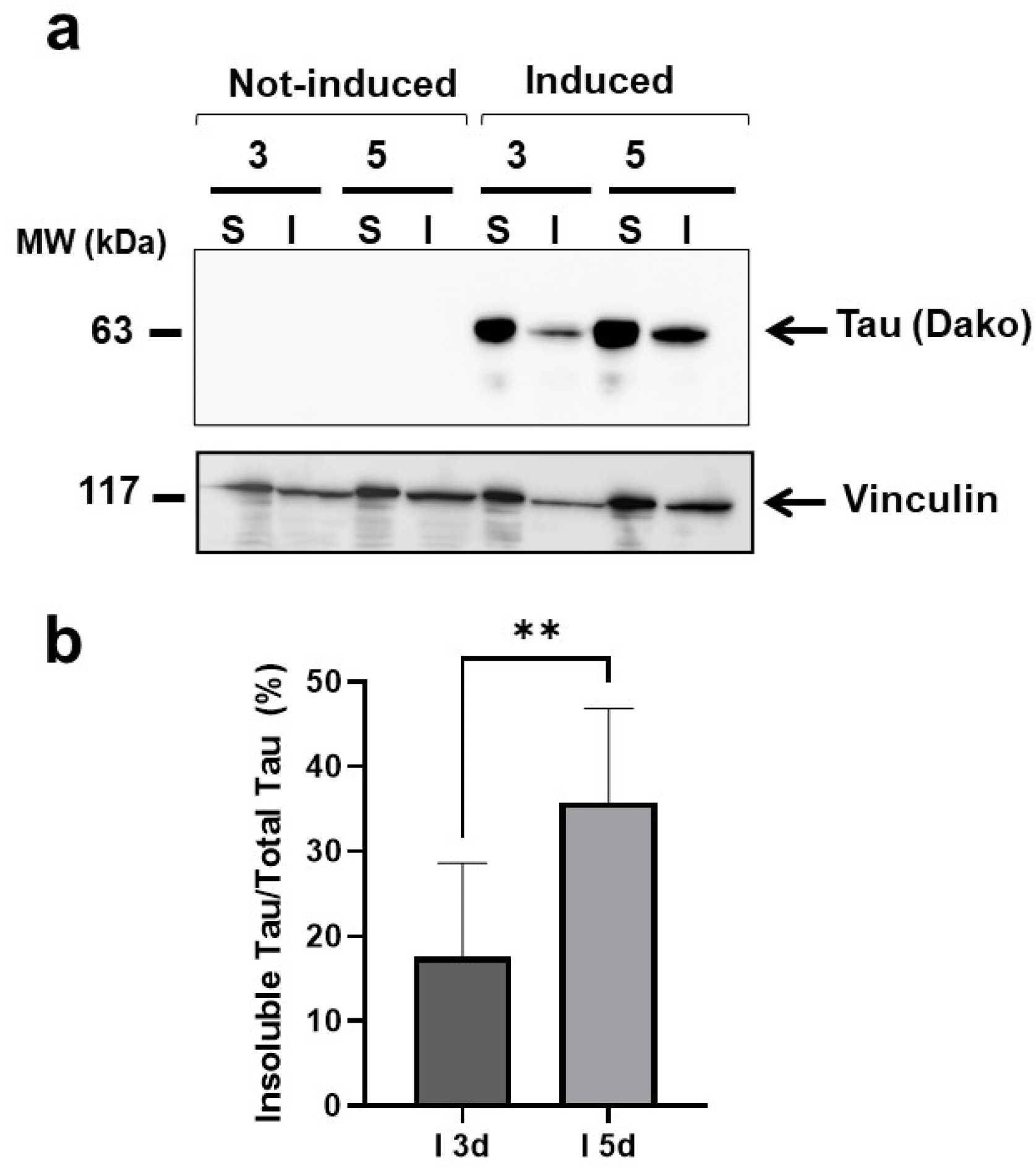
2.1. Multimeric Tau Assemblies from HEK T-Rex Are Proteotoxic for C. elegans
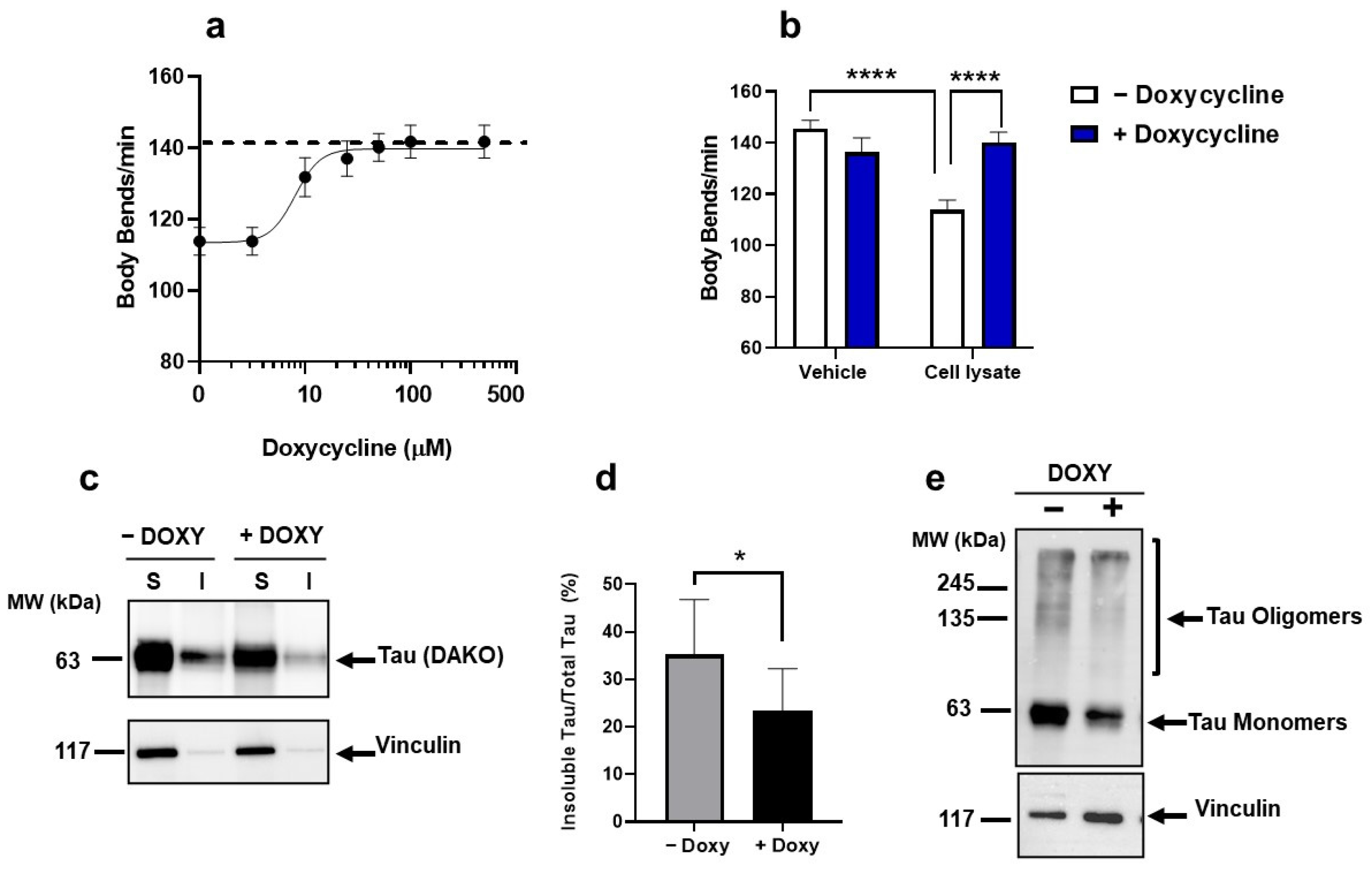
2.2. Doxy Protected from the Toxicity Induced by Multimeric Tau
3. Discussion
4. Materials and Methods
4.1. HEK T-Rex Cells
4.2. Immunochemical Analysis
4.3. Detergent Insolubility Assay
4.4. Tau Immunoprecipitation
4.5. C. elegans Studies
4.6. Effect of Doxy
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, G.; Leugers, C.J. Tau and Tauopathies. Prog. Mol. Biol. Transl. Sci. 2012, 107, 263–293. [Google Scholar] [CrossRef] [PubMed]
- Brunden, K.R.; Trojanowski, J.Q.; Lee, V.M.-Y. Evidence That Non-Fibrillar Tau Causes Pathology Linked To Neurodegeneration And Behavioral Impairments. J. Alzheimers Dis. 2008, 14, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Tau Suppression in a Neurodegenerative Mouse Model Improves Memory Function—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/16020737/ (accessed on 12 July 2022).
- Lasagna-Reeves, C.A.; Castillo-Carranza, D.L.; Sengupta, U.; Sarmiento, J.; Troncoso, J.; Jackson, G.R.; Kayed, R. Identification of Oligomers at Early Stages of Tau Aggregation in Alzheimer’s Disease. FASEB J. 2012, 26, 1946–1959. [Google Scholar] [CrossRef] [PubMed]
- Ghag, G.; Bhatt, N.; Cantu, D.V.; Guerrero-Munoz, M.J.; Ellsworth, A.; Sengupta, U.; Kayed, R. Soluble Tau Aggregates, Not Large Fibrils, Are the Toxic Species That Display Seeding and Cross-seeding Behavior. Protein Sci. 2018, 27, 1901–1909. [Google Scholar] [CrossRef]
- Usenovic, M.; Niroomand, S.; Drolet, R.E.; Yao, L.; Gaspar, R.C.; Hatcher, N.G.; Schachter, J.; Renger, J.J.; Parmentier-Batteur, S. Internalized Tau Oligomers Cause Neurodegeneration by Inducing Accumulation of Pathogenic Tau in Human Neurons Derived from Induced Pluripotent Stem Cells. J. Neurosci. 2015, 35, 14234–14250. [Google Scholar] [CrossRef]
- Fá, M.; Puzzo, D.; Piacentini, R.; Staniszewski, A.; Zhang, H.; Baltrons, M.A.; Li Puma, D.D.; Chatterjee, I.; Li, J.; Saeed, F.; et al. Extracellular Tau Oligomers Produce An Immediate Impairment of LTP and Memory. Sci. Rep. 2016, 6, 19393. [Google Scholar] [CrossRef]
- Berger, Z.; Roder, H.; Hanna, A.; Carlson, A.; Rangachari, V.; Yue, M.; Wszolek, Z.; Ashe, K.; Knight, J.; Dickson, D.; et al. Accumulation of Pathological Tau Species and Memory Loss in a Conditional Model of Tauopathy. J. Neurosci. 2007, 27, 3650–3662. [Google Scholar] [CrossRef]
- Arrasate, M.; Pérez, M.; Armas-Portela, R.; Ávila, J. Polymerization of Tau Peptides into Fibrillar Structures. The Effect of FTDP-17 Mutations. FEBS Lett. 1999, 446, 199–202. [Google Scholar] [CrossRef]
- Nacharaju, P.; Lewis, J.; Easson, C.; Yen, S.; Hackett, J.; Hutton, M.; Yen, S.H. Accelerated Filament Formation from Tau Protein with Specific FTDP-17 Missense Mutations. FEBS Lett. 1999, 447, 195–199. [Google Scholar] [CrossRef]
- Dayanandan, R.; Van Slegtenhorst, M.; Mack, T.G.A.; Ko, L.; Yen, S.-H.; Leroy, K.; Brion, J.-P.; Anderton, B.H.; Hutton, M.; Lovestone, S. Mutations in Tau Reduce Its Microtubule Binding Properties in Intact Cells and Affect Its Phosphorylation. FEBS Lett. 1999, 446, 228–232. [Google Scholar] [CrossRef]
- Miyasaka, T.; Morishima-Kawashima, M.; Ravid, R.; Kamphorst, W.; Nagashima, K.; Ihara, Y. Selective Deposition of Mutant Tau in the FTDP-17 Brain Affected by the P301L Mutation. J. Neuropathol. Exp. Neurol. 2001, 60, 872–884. [Google Scholar] [CrossRef] [PubMed]
- Lasagna-Reeves, C.A.; Castillo-Carranza, D.L.; Sengupta, U.; Clos, A.L.; Jackson, G.R.; Kayed, R. Tau Oligomers Impair Memory and Induce Synaptic and Mitochondrial Dysfunction in Wild-Type Mice. Mol. Neurodegener. 2011, 6, 39. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Davidowitz, E.; Lopez, P.; Emadi, S.; Moe, J.; Sierks, M. Trimeric Tau Is Toxic to Human Neuronal Cells at Low Nanomolar Concentrations. Int. J. Cell Biol. 2013, 2013, 260787. [Google Scholar] [CrossRef]
- Mirbaha, H.; Holmes, B.B.; Sanders, D.W.; Bieschke, J.; Diamond, M.I. Tau Trimers Are the Minimal Propagation Unit Spontaneously Internalized to Seed Intracellular Aggregation. J. Biol. Chem. 2015, 290, 14893–14903. [Google Scholar] [CrossRef] [PubMed]
- Yao, F.; Svensjö, T.; Winkler, T.; Lu, M.; Eriksson, C.; Eriksson, E. Tetracycline Repressor, TetR, Rather than the TetR–Mammalian Cell Transcription Factor Fusion Derivatives, Regulates Inducible Gene Expression in Mammalian Cells. Hum. Gene Ther. 1998, 9, 1939–1950. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, B.; Li, G.; Yin, H.; Kuret, J. Tau Aggregation and Toxicity in a Cell Culture Model of Tauopathy. J. Biol. Chem. 2007, 282, 16454–16464. [Google Scholar] [CrossRef]
- Crowe, A.; Henderson, M.J.; Anderson, J.; Titus, S.A.; Zakharov, A.; Simeonov, A.; Buist, A.; Delay, C.; Moechars, D.; Trojanowski, J.Q.; et al. Compound Screening in Cell-Based Models of Tau Inclusion Formation: Comparison of Primary Neuron and HEK293 Cell Assays. J. Biol. Chem. 2020, 295, 4001–4013. [Google Scholar] [CrossRef]
- Natale, C.; Barzago, M.M.; Diomede, L. Caenorhabditis Elegans Models to Investigate the Mechanisms Underlying Tau Toxicity in Tauopathies. Brain Sci 2020, 10, 838. [Google Scholar] [CrossRef]
- Zanier, E.R.; Bertani, I.; Sammali, E.; Pischiutta, F.; Chiaravalloti, M.A.; Vegliante, G.; Masone, A.; Corbelli, A.; Smith, D.H.; Menon, D.K.; et al. Induction of a Transmissible Tau Pathology by Traumatic Brain Injury. Brain 2018, 141, 2685–2699. [Google Scholar] [CrossRef]
- Zanier, E.R.; Barzago, M.M.; Vegliante, G.; Romeo, M.; Restelli, E.; Bertani, I.; Natale, C.; Colnaghi, L.; Colombo, L.; Russo, L.; et al. C. Elegans Detects Toxicity of Traumatic Brain Injury Generated Tau. Neurobiol. Dis. 2021, 153, 105330. [Google Scholar] [CrossRef]
- Stoilova, T.; Colombo, L.; Forloni, G.; Tagliavini, F.; Salmona, M. A New Face for Old Antibiotics: Tetracyclines in Treatment of Amyloidoses. J. Med. Chem. 2013, 56, 5987–6006. [Google Scholar] [CrossRef] [PubMed]
- Diomede, L.; Rognoni, P.; Lavatelli, F.; Romeo, M.; del Favero, E.; Cantù, L.; Ghibaudi, E.; di Fonzo, A.; Corbelli, A.; Fiordaliso, F.; et al. A Caenorhabditis Elegans-Based Assay Recognizes Immunoglobulin Light Chains Causing Heart Amyloidosis. Blood 2014, 123, 3543–3552. [Google Scholar] [CrossRef] [PubMed]
- Giorgino, T.; Mattioni, D.; Hassan, A.; Milani, M.; Mastrangelo, E.; Barbiroli, A.; Verhelle, A.; Gettemans, J.; Barzago, M.M.; Diomede, L.; et al. Nanobody Interaction Unveils Structure, Dynamics and Proteotoxicity of the Finnish-Type Amyloidogenic Gelsolin Variant. Biochim. Biophys. Acta (BBA) -Mol. Basis Dis. 2019, 1865, 648–660. [Google Scholar] [CrossRef] [PubMed]
- Zeinolabediny, Y.; Caccuri, F.; Colombo, L.; Morelli, F.; Romeo, M.; Rossi, A.; Schiarea, S.; Ciaramelli, C.; Airoldi, C.; Weston, R.; et al. HIV-1 Matrix Protein P17 Misfolding Forms Toxic Amyloidogenic Assemblies That Induce Neurocognitive Disorders. Sci. Rep. 2017, 7, 10313. [Google Scholar] [CrossRef] [PubMed]
- Medina, L.; González-Lizárraga, F.; Dominguez-Meijide, A.; Ploper, D.; Parrales, V.; Sequeira, S.; Cima-Omori, M.-S.; Zweckstetter, M.; Del Bel, E.; Michel, P.P.; et al. Doxycycline Interferes With Tau Aggregation and Reduces Its Neuronal Toxicity. Front Aging Neurosci. 2021, 13, 635760. [Google Scholar] [CrossRef] [PubMed]
- Pagliuso, A.; Tham, T.N.; Allemand, E.; Robertin, S.; Dupuy, B.; Bertrand, Q.; Bécavin, C.; Koutero, M.; Najburg, V.; Nahori, M.-A.; et al. An RNA-Binding Protein Secreted by a Bacterial Pathogen Modulates RIG-I Signaling. Cell Host Microbe 2019, 26, 823–835.e11. [Google Scholar] [CrossRef]
- Topalidou, I.; Cattin-Ortolá, J.; Hummer, B.; Asensio, C.S.; Ailion, M. EIPR1 Controls Dense-Core Vesicle Cargo Retention and EARP Complex Localization in Insulin-Secreting Cells. Mol. Biol. Cell 2020, 31, 59–79. [Google Scholar] [CrossRef]
- Perni, M.; Aprile, F.A.; Casford, S.; Mannini, B.; Sormanni, P.; Dobson, C.M.; Vendruscolo, M. Delivery of Native Proteins into C. Elegans Using a Transduction Protocol Based on Lipid Vesicles. Sci. Rep. 2017, 7, 15045. [Google Scholar] [CrossRef]
- Perni, M.; Mannini, B.; Xu, C.K.; Kumita, J.R.; Dobson, C.M.; Chiti, F.; Vendruscolo, M. Exogenous Misfolded Protein Oligomers Can Cross the Intestinal Barrier and Cause a Disease Phenotype in C. Elegans. Sci. Rep. 2021, 11, 14391. [Google Scholar] [CrossRef]
- Limbocker, R.; Staats, R.; Chia, S.; Ruggeri, F.S.; Mannini, B.; Xu, C.K.; Perni, M.; Cascella, R.; Bigi, A.; Sasser, L.R.; et al. Squalamine and Its Derivatives Modulate the Aggregation of Amyloid-β and α-Synuclein and Suppress the Toxicity of Their Oligomers. Front Neurosci. 2021, 15, 680026. [Google Scholar] [CrossRef]
- Ward, J.E.; Ren, R.; Toraldo, G.; SooHoo, P.; Guan, J.; O’Hara, C.; Jasuja, R.; Trinkaus-Randall, V.; Liao, R.; Connors, L.H.; et al. Doxycycline Reduces Fibril Formation in a Transgenic Mouse Model of AL Amyloidosis. Blood 2011, 118, 6610–6617. [Google Scholar] [CrossRef] [PubMed]
- Gautieri, A.; Beeg, M.; Gobbi, M.; Rigoldi, F.; Colombo, L.; Salmona, M. The Anti-Amyloidogenic Action of Doxycycline: A Molecular Dynamics Study on the Interaction with Aβ42. Int. J. Mol. Sci. 2019, 20, 4641. [Google Scholar] [CrossRef] [PubMed]
- Forloni, G.; Colombo, L.; Girola, L.; Tagliavini, F.; Salmona, M. Anti-Amyloidogenic Activity of Tetracyclines: Studies in Vitro. FEBS Lett. 2001, 487, 404–407. [Google Scholar] [CrossRef]
- Diomede, L.; Cassata, G.; Fiordaliso, F.; Salio, M.; Ami, D.; Natalello, A.; Doglia, S.M.; De Luigi, A.; Salmona, M. Tetracycline and Its Analogues Protect Caenorhabditis Elegans from β Amyloid-Induced Toxicity by Targeting Oligomers. Neurobiol. Dis. 2010, 40, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Balducci, C.; Santamaria, G.; La Vitola, P.; Brandi, E.; Grandi, F.; Viscomi, A.R.; Beeg, M.; Gobbi, M.; Salmona, M.; Ottonello, S.; et al. Doxycycline Counteracts Neuroinflammation Restoring Memory in Alzheimer’s Disease Mouse Models. Neurobiol. Aging 2018, 70, 128–139. [Google Scholar] [CrossRef]
- González-Lizárraga, F. Repurposing Doxycycline for Synucleinopathies: Remodelling of α-Synuclein Oligomers towards Non-Toxic Parallel Beta-Sheet Structured Species. Sci. Rep. 2017, 7, 41755. [Google Scholar] [CrossRef]
- Saivin, S.; Houin, G. Clinical Pharmacokinetics of Doxycycline and Minocycline. Clin-Pharmacokinet 1988, 15, 355–366. [Google Scholar] [CrossRef]
- Kaufman, S.K.; Sanders, D.W.; Thomas, T.L.; Ruchinskas, A.; Vaquer-Alicea, J.; Sharma, A.M.; Miller, T.M.; Diamond, M.I. Tau Prion Strains Dictate Patterns of Cell Pathology, Progression Rate, and Regional Vulnerability in Vivo. Neuron 2016, 92, 796–812. [Google Scholar] [CrossRef]
- Porta-de-la-Riva, M.; Fontrodona, L.; Villanueva, A.; Cerón, J. Basic Caenorhabditis Elegans Methods: Synchronization and Observation. J. Vis. Exp. 2012, 4019. [Google Scholar] [CrossRef]
- Morelli, F.; Romeo, M.; Barzago, M.M.; Bolis, M.; Mattioni, D.; Rossi, G.; Tagliavini, F.; Bastone, A.; Salmona, M.; Diomede, L. V363I and V363A Mutated Tau Affect Aggregation and Neuronal Dysfunction Differently in C. Elegans. Neurobiol. Dis. 2018, 117, 226–234. [Google Scholar] [CrossRef]
- Diomede, L.; Di Fede, G.; Romeo, M.; Bagnati, R.; Ghidoni, R.; Fiordaliso, F.; Salio, M.; Rossi, A.; Catania, M.; Paterlini, A.; et al. Expression of A2V-Mutated Aβ in Caenorhabditis Elegans Results in Oligomer Formation and Toxicity. Neurobiol. Dis. 2014, 62, 521–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Natale, C.; Barzago, M.M.; Colnaghi, L.; De Luigi, A.; Orsini, F.; Fioriti, L.; Diomede, L. A Combined Cell-Worm Approach to Search for Compounds Counteracting the Toxicity of Tau Oligomers In Vivo. Int. J. Mol. Sci. 2022, 23, 11277. https://doi.org/10.3390/ijms231911277
Natale C, Barzago MM, Colnaghi L, De Luigi A, Orsini F, Fioriti L, Diomede L. A Combined Cell-Worm Approach to Search for Compounds Counteracting the Toxicity of Tau Oligomers In Vivo. International Journal of Molecular Sciences. 2022; 23(19):11277. https://doi.org/10.3390/ijms231911277
Chicago/Turabian StyleNatale, Carmina, Maria Monica Barzago, Luca Colnaghi, Ada De Luigi, Franca Orsini, Luana Fioriti, and Luisa Diomede. 2022. "A Combined Cell-Worm Approach to Search for Compounds Counteracting the Toxicity of Tau Oligomers In Vivo" International Journal of Molecular Sciences 23, no. 19: 11277. https://doi.org/10.3390/ijms231911277