

Aerobic Exercise Prevents Arterial Stiffness and Attenuates Hyperexcitation of Sympathetic Nerves in Perivascular Adipose Tissue of Mice after Transverse Aortic Constriction

Abstract
:1. Introduction
2. Results
2.1. The Developmental Course of Heart Failure after TAC Was Delayed by Exercise
2.2. The Remodeling of ECM after TAC Was Prevented by Exercise
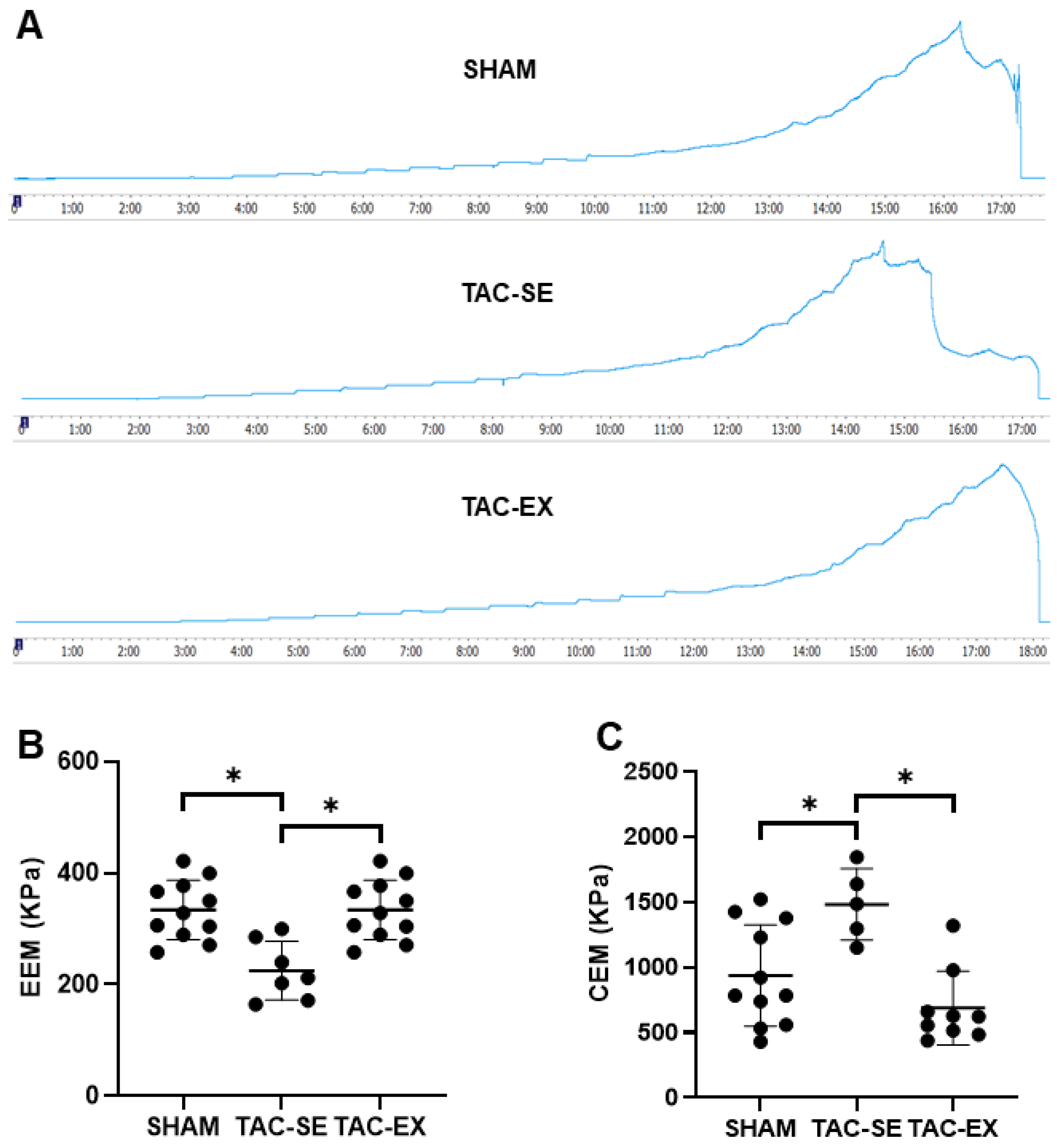
2.3. The Thoracic Arterial Stiffness after TAC Were Prevented by Exercise
2.4. Hyperexcitation of Sympathetic Nerves within PVAT after TAC Was Attenuated by Exercise
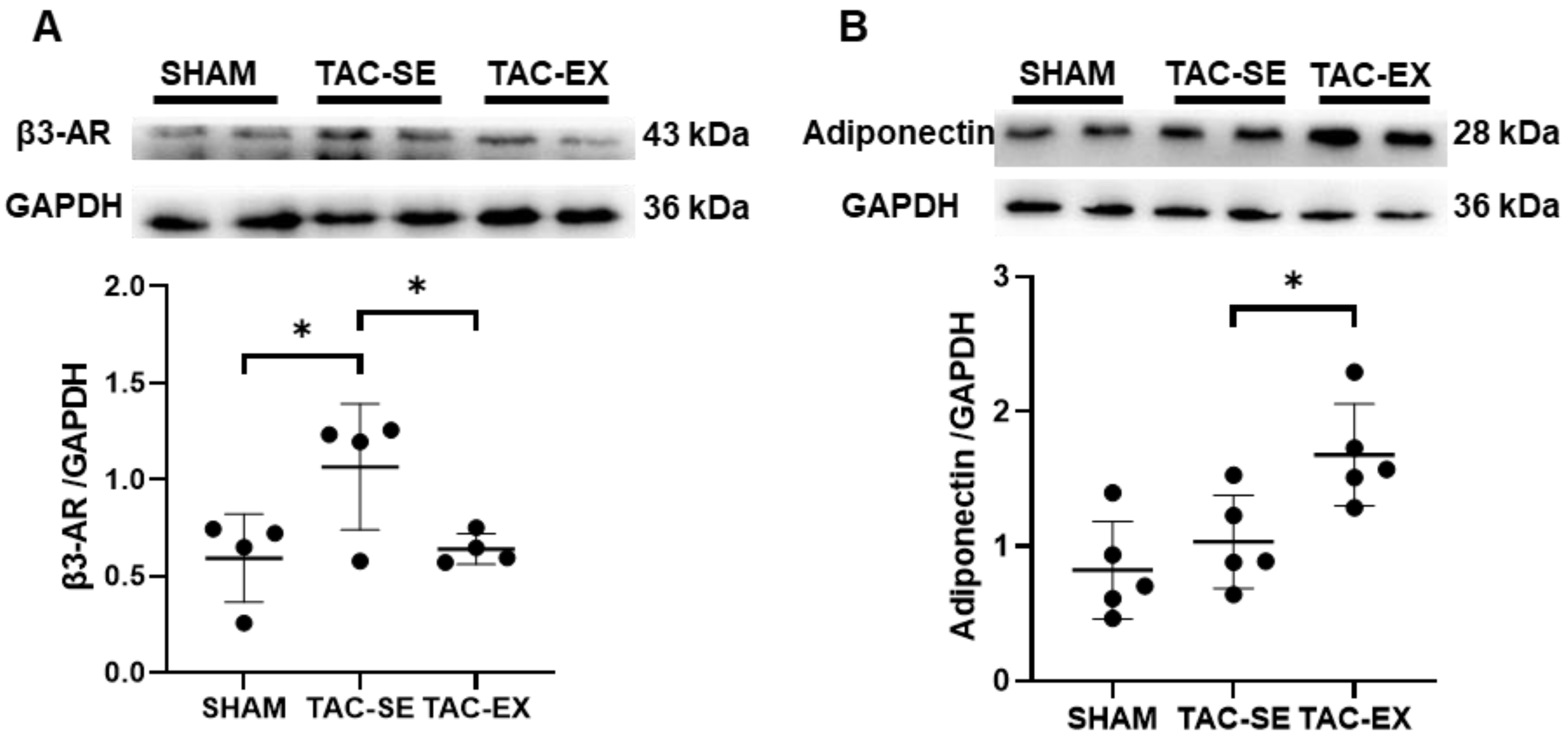
2.5. β3-Adrenergic Receptor (β3-AR) and Adiponectin within PVAT after TAC Was Influenced by Exercise
3. Discussion
4. Materials and Methods
4.1. Animal Care and Experimental Design
4.2. Establishment of Transverse Aortic Constriction Model
4.3. Exercise Protocol
4.4. Echocardiography
4.5. Histological Assessments
4.6. Mechanical Stiffness Testing
4.7. Enzyme Linked Immunosorbent Assay (ELISA)
4.8. Western Blotting
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mitchell, G.F.; Hwang, S.-J.; Vasan, R.S.; Larson, M.G.; Pencina, M.J.; Hamburg, N.M.; Vita, J.A.; Levy, D.; Benjamin, E.J. Arterial Stiffness and Cardiovascular Events. Circulation 2010, 121, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Marti, C.N.; Gheorghiade, M.; Kalogeropoulos, A.P.; Georgiopoulou, V.V.; Quyyumi, A.A.; Butler, J. Endothelial dysfunction, arterial stiffness, and heart failure. J. Am. Coll. Cardiol. 2012, 60, 1455–1469. [Google Scholar] [CrossRef] [PubMed]
- Chirinos, J.A.; Segers, P.; Hughes, T.; Townsend, R. Large.e-Artery Stiffness in Health and Disease: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 2019, 74, 1237–1263. [Google Scholar] [CrossRef] [PubMed]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef] [PubMed]
- Fleenor, B.S.; Carlini, N.A.; Ouyang, A.; Harber, M.P. Perivascular adipose tissue-mediated arterial stiffening in aging and disease: An emerging translational therapeutic target? Pharmacol. Res. 2022, 178, 106150. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.-Y.; Qu, S.-L.; Xiong, W.-H.; Rom, O.; Chang, L.; Jiang, Z.-S. Perivascular adipose tissue (PVAT) in atherosclerosis: A double-edged sword. Cardiovasc. Diabetol. 2018, 17, 134. [Google Scholar] [CrossRef] [PubMed]
- Szasz, T.; Bomfim, G.F.; Webb, R.C. The influence of perivascular adipose tissue on vascular homeostasis. Vasc. Health Risk Manag. 2013, 9, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Y.; Anderson, A.S. The sympathetic nervous system and heart failure. Cardiol. Clin. 2014, 32, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Grassi, G.; Biffi, A.; Seravalle, G.; Trevano, F.Q.; Dell’Oro, R.; Corrao, G.; Mancia, G. Sympathetic Neural Overdrive in the Obese and Overweight State. Hypertension 2019, 74, 349–358. [Google Scholar] [CrossRef]
- Nardone, M.; Floras, J.S.; Millar, P.J. Sympathetic neural modulation of arterial stiffness in humans. Am. J. Physiol. Heart Circ. Physiol. 2020, 319, H1338–H1346. [Google Scholar] [CrossRef] [PubMed]
- Saxton, S.N.; Ryding, K.E.; Aldous, R.G.; Withers, S.B.; Ohanian, J.; Heagerty, A.M. Role of Sympathetic Nerves and Adipocyte Catecholamine Uptake in the Vasorelaxant Function of Perivascular Adipose Tissue. Arter. Thromb Vasc. Biol 2018, 38, 880–891. [Google Scholar] [CrossRef] [PubMed]
- Sabbatini, A.R.; Fontana, V.; Laurent, S.; Moreno, H. An update on the role of adipokines in arterial stiffness and hypertension. J. Hypertens. 2015, 33, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Sowka, A.; Dobrzyn, P. Role of Perivascular Adipose Tissue-Derived Adiponectin in Vascular Homeostasis. Cells 2021, 10, 1485. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.S.; Walker, S.; Smart, N.A.; Piepoli, M.F.; Warren, F.C.; Ciani, O.; Whellan, D.; O’Connor, C.; Keteyian, S.J.; Coats, A.; et al. Impact of Exercise Rehabilitation on Exercise Capacity and Quality-of-Life in Heart Failure. J. Am. Coll. Cardiol. 2019, 73, 1430–1443. [Google Scholar] [CrossRef] [PubMed]
- Lopes, S.; Afreixo, V.; Teixeira, M.; Garcia, C.; Leitão, C.; Gouveia, M.; Figueiredo, D.; Alves, A.J.; Polonia, J.; Oliveira, J.; et al. Exercise training reduces arterial stiffness in adults with hypertension: A systematic review and meta-analysis. J. Hypertens. 2021, 39, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Park, J.B.; Lee, S.Y.; Han, S.K.; Ryu, P.D. Exercise training normalizes elevated firing rate of hypothalamic presympathetic neurons in heart failure rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019, 316, R110–R120. [Google Scholar] [CrossRef] [PubMed]
- Leggio, M.; Fusco, A.; Loreti, C.; Limongelli, G.; Bendini, M.G.; Mazza, A.; Coraci, D.; Padua, L. Effects of exercise training in heart failure with preserved ejection fraction: An updated systematic literature review. Heart Fail. Rev. 2020, 25, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, R.; Kasahara, Y.; Ikeo, T.; Asaki, K.; Sato, K.; Matsui, T.; Iwanuma, S.; Ohashi, N.; Hashiguchi, T. Effects of different intensities and durations of aerobic exercise training on arterial stiffness. J. Phys. Ther. Sci. 2020, 32, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.; Liu, Y.; Xu, Y.; Xu, D. Association Between Arterial Stiffness and Heart Failure With Preserved Ejection Fraction. Front. Cardiovasc. Med. 2021, 8, 707162. [Google Scholar] [CrossRef]
- Ouyang, A.; Olver, T.D.; Emter, C.A.; Fleenor, B.S. Chronic exercise training prevents coronary artery stiffening in aortic-banded miniswine: Role of perivascular adipose-derived advanced glycation end products. J. Appl. Physiol. 2019, 127, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Saxton, S.N.; Toms, L.K.; Aldous, R.G.; Withers, S.B.; Ohanian, J.; Heagerty, A.M. Restoring Perivascular Adipose Tissue Function in Obesity Using Exercise. Cardiovasc. Drugs Ther. 2021, 35, 1291–1304. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, M.J.; Romero, F.; Roa, J.; Rodriguez-Nunez, I. Exercise training to reduce sympathetic nerve activity in heart failure patients. A systematic review and meta-analysis. Braz. J. Phys. Ther. 2018, 22, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, A.; Ahmed, K.; Zhang, Y. β-Adrenergic Receptor Desensitization/Down-Regulation in Heart Failure: A Friend or Foe? Front. Cardiovasc. Med. 2022, 9, 925692. [Google Scholar] [CrossRef] [PubMed]
- Quattrocelli, M.; Wintzinger, M.; Miz, K.; Panta, M.; Prabakaran, A.D.; Barish, G.D.; Chandel, N.S.; McNally, E.M. Intermittent prednisone treatment in mice promotes exercise tolerance in obesity through adiponectin. J. Exp. Med. 2022, 219. [Google Scholar] [CrossRef] [PubMed]
- Margaritis, M.; Antonopoulos, A.S.; Digby, J.; Lee, R.; Reilly, S.; Coutinho, P.; Shirodaria, C.; Sayeed, R.; Petrou, M.; De Silva, R.; et al. Interactions between vascular wall and perivascular adipose tissue reveal novel roles for adiponectin in the regulation of endothelial nitric oxide synthase function in human vessels. Circulation 2013, 127, 2209–2221. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Polaki, V.; Chen, S.; Bihl, J.C. Exercise Improves Endothelial Function Associated with Alleviated Inflammation and Oxidative Stress of Perivascular Adipose Tissue in Type 2 Diabetic Mice. Oxidative Med. Cell. Longev. 2020, 2020, 8830537. [Google Scholar] [CrossRef]
- DeVallance, E.R.; Branyan, K.W.; Olfert, I.M.; Pistilli, E.E.; Bryner, R.W.; Kelley, E.E.; Frisbee, J.C.; Chantler, P.D. Chronic stress induced perivascular adipose tissue impairment of aortic function and the therapeutic effect of exercise. Exp. Physiol. 2021, 106, 1343–1358. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Da, Q.; Cao, S.; Yan, K.; Shi, Z.; Miao, Q.; Li, C.; Hu, L.; Sun, S.; Wu, W.; et al. HINT1 (Histidine Triad Nucleotide-Binding Protein 1) Attenuates Cardiac Hypertrophy Via Suppressing HOXA5 (Homeobox A5) Expression. Circulation 2021, 144, 638–654. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Zhong, G.; Li, J.; Pan, J.; Zhao, Y.; Song, H.; Sun, W.; Jin, X.; Li, Y.; Du, R.; et al. Targeting E3 Ubiquitin Ligase WWP1 Prevents Cardiac Hypertrophy Through Destabilizing DVL2 via Inhibition of K27-Linked Ubiquitination. Circulation 2021, 144, 694–711. [Google Scholar] [CrossRef]
- Au-deAlmeida, A.C.; Au-van Oort, R.J.; Au-Wehrens, X.H.T. Transverse Aortic Constriction in Mice. JoVE 2010, e1729. [Google Scholar] [CrossRef]
- Fernando, P.; Bonen, A.; Hoffman-Goetz, L. Predicting submaximal oxygen consumption during treadmill running in mice. Can. J. Physiol. Pharmacol. 1993, 71, 854–857. [Google Scholar] [CrossRef] [PubMed]
- Høydal, M.A.; Wisløff, U.; Kemi, O.J.; Ellingsen, O. Running speed and maximal oxygen uptake in rats and mice: Practical implications for exercise training. Eur. J. Cardiovasc. Prev. Rehabil. Off. J. Eur. Soc. Cardiol. Work. Groups Epidemiol. Prev. Card. Rehabil. Exerc. Physiol. 2007, 14, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Wang, G.; Zheng, N.; Cheng, W.; Ouyang, K.; Lin, H.; Liao, Y.; Liu, J. HIMF (Hypoxia-Induced Mitogenic Factor)-IL (Interleukin)-6 Signaling Mediates Cardiomyocyte-Fibroblast Crosstalk to Promote Cardiac Hypertrophy and Fibrosis. Hypertension 2019, 73, 1058–1070. [Google Scholar] [CrossRef] [PubMed]
- Malek Mohammadi, M.; Abouissa, A.; Heineke, J. A surgical mouse model of neonatal pressure overload by transverse aortic constriction. Nat. Protoc. 2021, 16, 775–790. [Google Scholar] [CrossRef] [PubMed]
- Clavel, M.A.; Côté, N.; Mathieu, P.; Dumesnil, J.G.; Audet, A.; Pépin, A.; Couture, C.; Fournier, D.; Trahan, S.; Pagé, S.; et al. Paradoxical low-flow, low-gradient aortic stenosis despite preserved left ventricular ejection fraction: New insights from weights of operatively excised aortic valves. Eur. Heart J. 2014, 35, 2655–2662. [Google Scholar] [CrossRef]
- Chang, Z.S.; Xia, J.B.; Wu, H.Y.; Peng, W.T.; Jiang, F.Q.; Li, J.; Liang, C.Q.; Zhao, H.; Park, K.S.; Song, G.H.; et al. Forkhead box O3 protects the heart against paraquat-induced aging-associated phenotypes by upregulating the expression of antioxidant enzymes. Aging Cell 2019, 18, e12990. [Google Scholar] [CrossRef] [PubMed]
- Nishida, N.; Aoki, H.; Ohno-Urabe, S.; Nishihara, M.; Furusho, A.; Hirakata, S.; Hayashi, M.; Ito, S.; Yamada, H.; Hirata, Y.; et al. High Salt Intake Worsens Aortic Dissection in Mice: Involvement of IL (Interleukin)-17A-Dependent ECM (Extracellular Matrix) Metabolism. Arter. Thromb Vasc. Biol. 2020, 40, 189–205. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Feng, B.; Ma, X.; Sun, K.; Xu, G.; Zhou, Y. Dapagliflozin improves left ventricular remodeling and aorta sympathetic tone in a pig model of heart failure with preserved ejection fraction. Cardiovasc. Diabetol. 2019, 18, 107. [Google Scholar] [CrossRef] [PubMed]
- Murano, I.; Barbatelli, G.; Giordano, A.; Cinti, S. Noradrenergic parenchymal nerve fiber branching after cold acclimatisation correlates with brown adipocyte density in mouse adipose organ. J. Anat. 2009, 214, 171–178. [Google Scholar] [CrossRef]
- Ouyang, A.; Garner, T.B.; Fleenor, B.S. Hesperidin reverses perivascular adipose-mediated aortic stiffness with aging. Exp. Gerontol. 2017, 97, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.C.; Su, Y.H.; Jiang, F.Q.; Xia, J.B.; Wu, H.Y.; Chang, Z.S.; Peng, W.T.; Song, G.H.; Park, K.S.; Kim, S.K.; et al. CpG oligodeoxynucleotide preconditioning improves cardiac function after myocardial infarction via modulation of energy metabolism and angiogenesis. J. Cell. Physiol. 2018, 233, 4245–4257. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SHAM (n = 15) | TAC-SE (n = 12) | TAC-EX (n = 15) | |
|---|---|---|---|
| Body Weight (g) | 27.43 ± 2.09 | 26.65 ± 1.69 | 26.78 ± 1.90 |
| PVAT (mg) | 8.68 ± 1.83 | 7.31 ± 1.99 | 10.02 ± 2.29 * |
| PVAT (mg)/Body Weight (g) | 0.318 ± 0.064 | 0.269 ± 0.076 | 0.375 ± 0.071 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, N.; Xia, J.; Wang, C.; Zhou, J.; Huang, J.; Hu, M.; Liao, J. Aerobic Exercise Prevents Arterial Stiffness and Attenuates Hyperexcitation of Sympathetic Nerves in Perivascular Adipose Tissue of Mice after Transverse Aortic Constriction. Int. J. Mol. Sci. 2022, 23, 11189. https://doi.org/10.3390/ijms231911189
Shi N, Xia J, Wang C, Zhou J, Huang J, Hu M, Liao J. Aerobic Exercise Prevents Arterial Stiffness and Attenuates Hyperexcitation of Sympathetic Nerves in Perivascular Adipose Tissue of Mice after Transverse Aortic Constriction. International Journal of Molecular Sciences. 2022; 23(19):11189. https://doi.org/10.3390/ijms231911189
Chicago/Turabian StyleShi, Niujin, Jingbo Xia, Chaoge Wang, Jie Zhou, Junhao Huang, Min Hu, and Jingwen Liao. 2022. "Aerobic Exercise Prevents Arterial Stiffness and Attenuates Hyperexcitation of Sympathetic Nerves in Perivascular Adipose Tissue of Mice after Transverse Aortic Constriction" International Journal of Molecular Sciences 23, no. 19: 11189. https://doi.org/10.3390/ijms231911189