
Targeting the Maladaptive Effects of Binge Drinking on Circadian Gene Expression
 ,
,
Abstract
:1. Introduction
2. Results
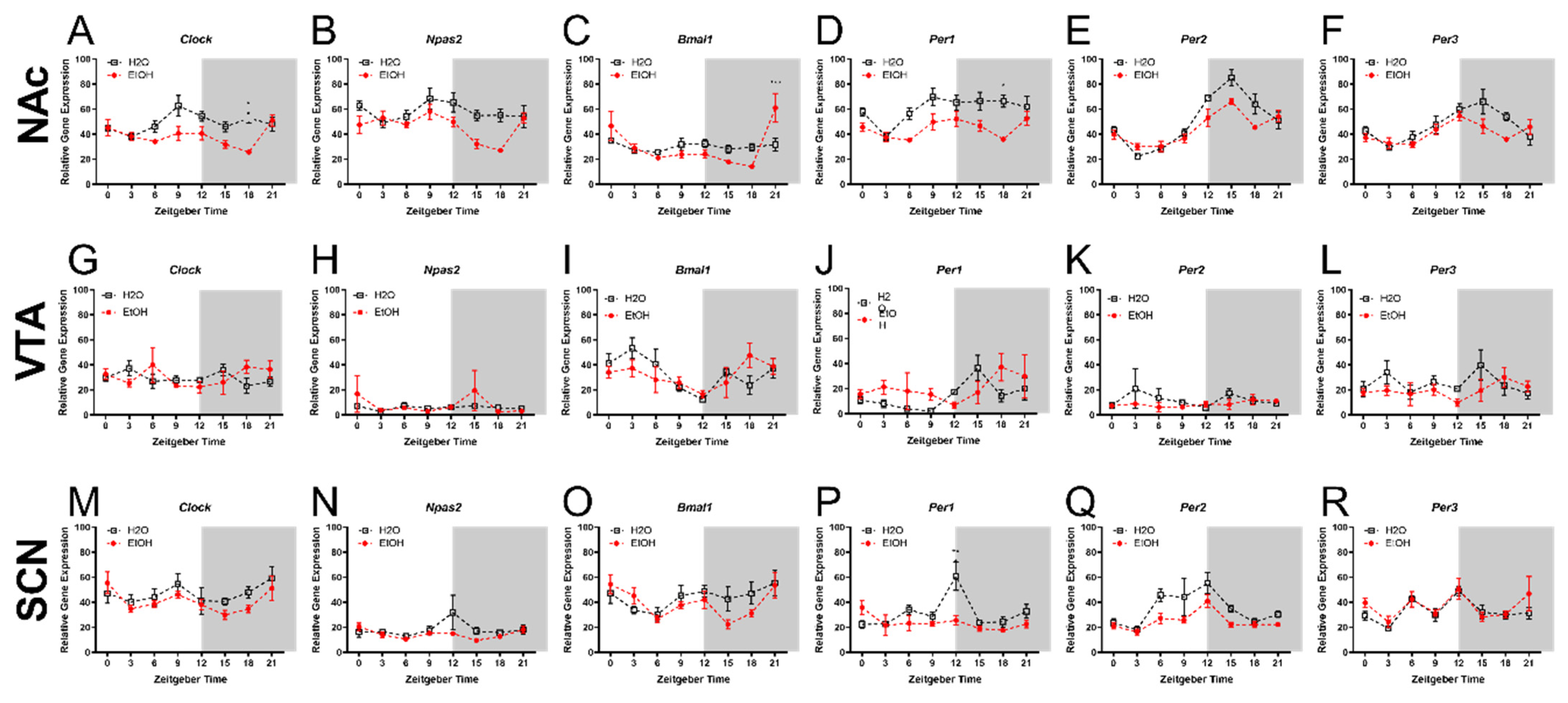
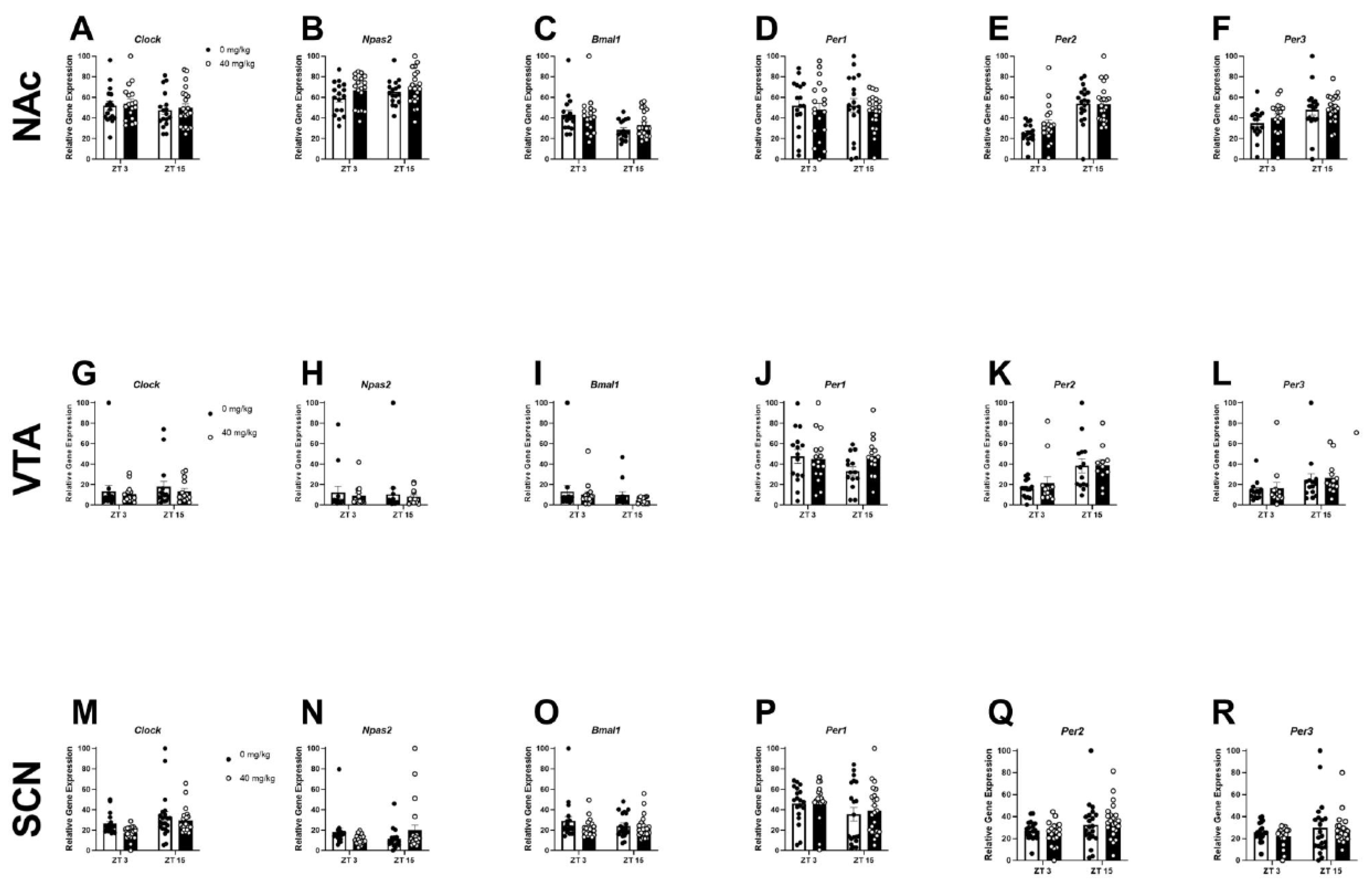
2.1. Experiment 1: Chronic Binge-like Ethanol Drinking Reduces NAc and SCN Expression of Circadian Genes
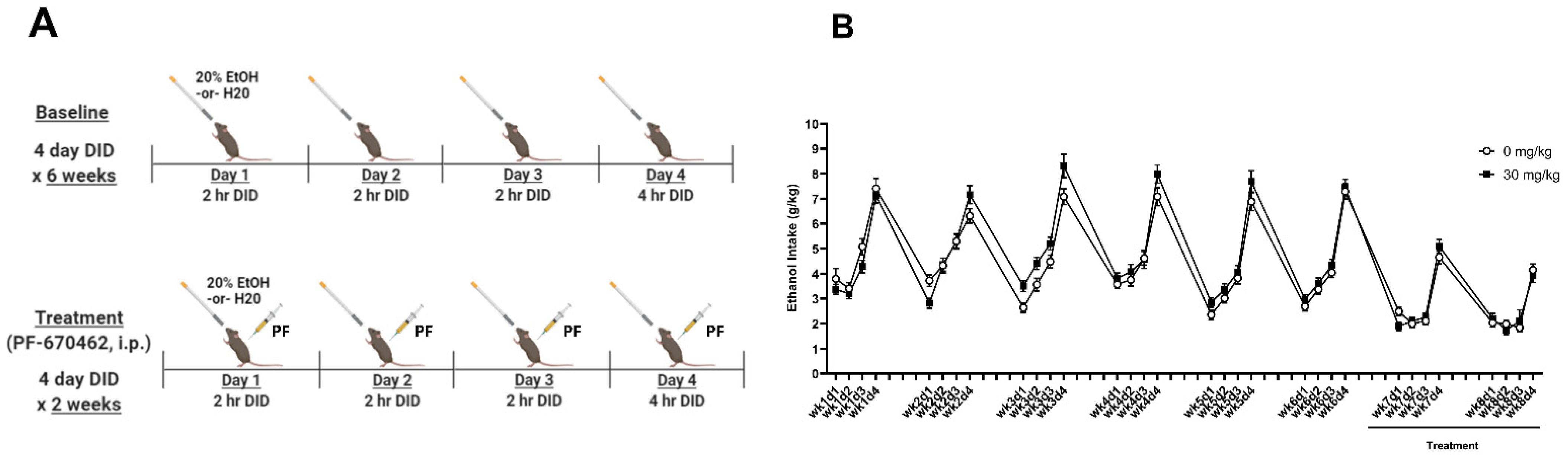
2.2. Experiment 2A: PF-670462 Did Not Reduce Chronic Binge-like Ethanol Intake
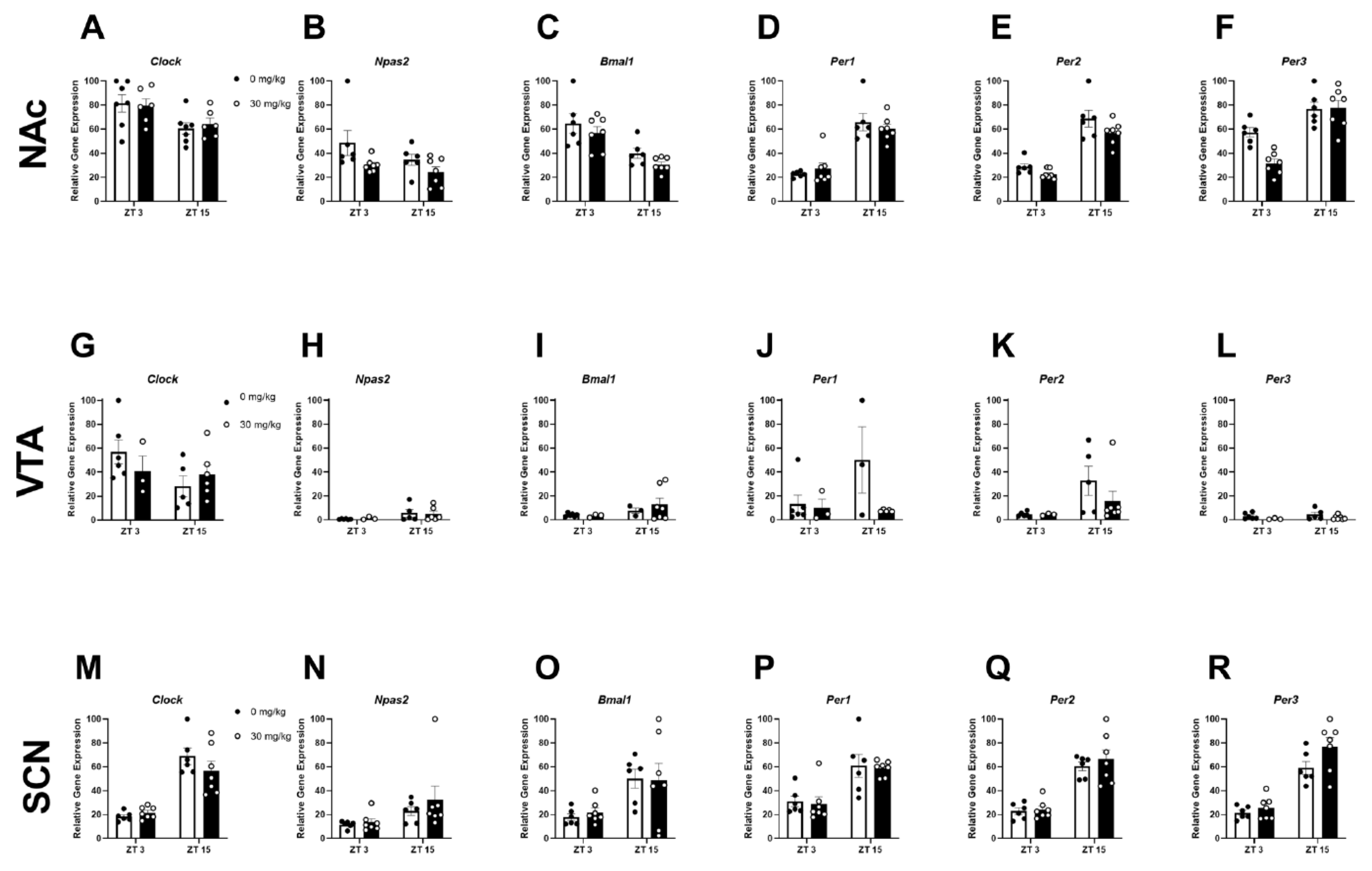
2.3. Experiment 2B: Inhibiting CK1E/D Reduces NAc Circadian Gene Expression
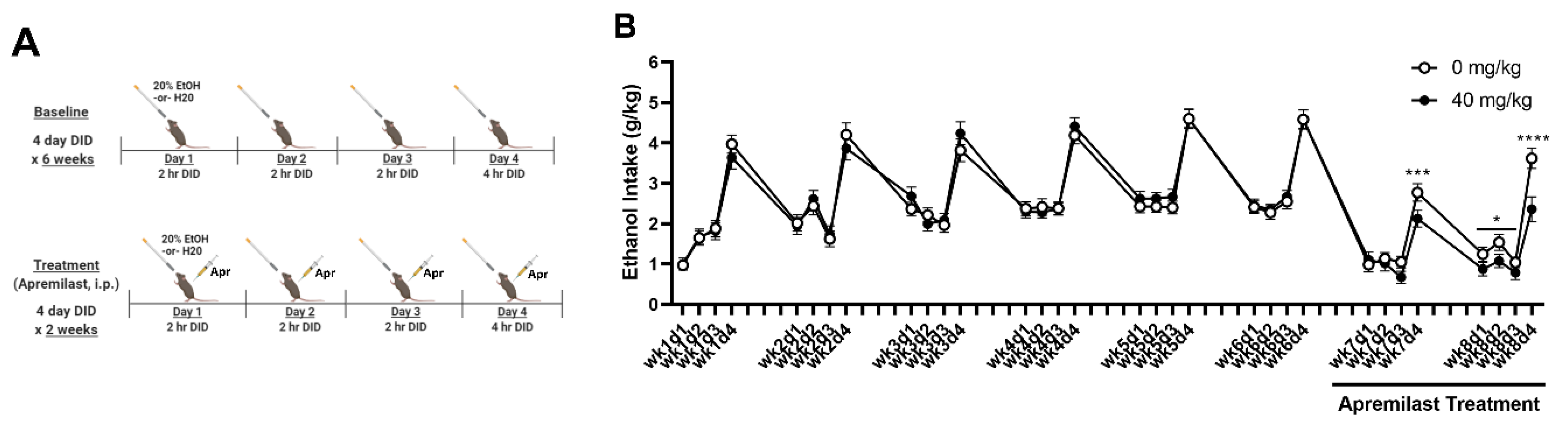
2.4. Experiment 3A: Apremilast Reduces Ethanol Intake in iHDID-1 Mice with a History of Chronic Binge-like Drinking
2.5. Experiment 3B: Apremilast Treatment Did Not Ameliorate the Effects of Ethanol on Circadian Gene Expression
3. Discussion
4. Methods and Materials
4.1. Animals
4.2. Drugs
4.3. Experiment 1: Determining Effects of Chronic Binge-like Drinking on Circadian Gene Expression in Brain Regions Important for AUDs
4.4. Experiment 2: Determine Whether Inhibition of CK1e/d (via Administration of PF-670462) Reduces Ethanol Intake and Rescues Ethanol-Induced Circadian Gene Disruptions
4.5. Experiment 3: Determine Whether Inhibition of PDE4 (via Administration Apremilast) Reduces Ethanol Intake and Rescues Ethanol-Induced Circadian Gene Disruptions
4.6. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carvalho, A.F.; Heilig, M.; Perez, A.; Probst, C.; Rehm, J. Alcohol use disorders. Lancet 2019, 394, 781–792. [Google Scholar] [CrossRef]
- Rehm, J. The risks associated with alcohol use and alcoholism. Alcohol Res. Health 2011, 34, 135–143. [Google Scholar]
- Landolt, H.-P.; Roth, C.; Dijk, D.-J.; Borbely, A.A. Late-Afternoon Ethanol Intake Affects Nocturnal Sleep and the Sleep EEG in Middle-Aged Men. J. Clin. Psychopharmacol. 1996, 16, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Danel, T.; Libersa, C.; Touitou, Y. The effect of alcohol consumption on the circadian control of human core body temperature is time dependent. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 281, R52–R55. [Google Scholar] [CrossRef]
- Rupp, T.L.; Acebo, C.; Carskadon, M.A. Evening Alcohol Suppresses Salivary Melatonin in Young Adults. Chronobiol. Int. 2007, 24, 463–470. [Google Scholar] [CrossRef]
- McCarthy, M.J.; Fernandes, M.; Kranzler, H.R.; Covault, J.M.; Welsh, D.K. Circadian Clock Period Inversely Correlates with Illness Severity in Cells from Patients with Alcohol Use Disorders. Alcohol. Clin. Exp. Res. 2013, 37, 1304–1310. [Google Scholar] [CrossRef]
- Spanagel, R.; Rosenwasser, A.M.; Schumann, G.; Sarkar, D.K. Alcohol Consumption and the Body’s Biological Clock. Alcohol. Clin. Exp. Res. 2005, 29, 1550–1557. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, J.S.; Hong, H.-K.; Ko, C.H.; McDearmon, E.L. The genetics of mammalian circadian order and disorder: Implications for physiology and disease. Nat. Rev. Genet. 2008, 9, 764–775. [Google Scholar] [CrossRef]
- Ozburn, A.R.; Falcon, E.; Twaddle, A.; Nugent, A.L.; Gillman, A.G.; Spencer, S.M.; Arey, R.N.; Mukherjee, S.; Lyons-Weiler, J.; Self, D.W.; et al. Direct Regulation of Diurnal Drd3 Expression and Cocaine Reward by NPAS2. Biol. Psychiatry 2015, 77, 425–433. [Google Scholar] [CrossRef]
- Pilorz, V.; Helfrich-Förster, C.; Oster, H. The role of the circadian clock system in physiology. Pflügers Arch. Eur. J. Physiol. 2018, 470, 227–239. [Google Scholar] [CrossRef]
- Rijo-Ferreira, F.; Takahashi, J.S. Genomics of circadian rhythms in health and disease. Genome Med. 2019, 11, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozburn, A.R.; Falcon, E.; Mukherjee, S.; Gillman, A.; Arey, R.; Spencer, S.; McClung, C.A. The role of clock in ethanol-related behaviors. Neuropsychopharmacology 2013, 38, 2393–2400. [Google Scholar] [CrossRef] [PubMed]
- Reppert, S.M.; Weaver, D.R. Molecular analysis of mammalian circadian rhythms. Annu. Rev. Physiol. 2001, 63, 647–676. [Google Scholar] [CrossRef]
- Sharma, R.; Puckett, H.; Kemerling, M.; Parikh, M.; Sahota, P.; Thakkar, M. Antisense-Induced Downregulation of Clock Genes in the Shell Region of the Nucleus Accumbens Reduces Binge Drinking in Mice. Alcohol. Clin. Exp. Res. 2021, 45, 530–542. [Google Scholar] [CrossRef] [PubMed]
- Sjöholm, L.K.; Kovanen, L.; Saarikoski, S.T.; Schalling, M.; Lavebratt, C.; Partonen, T. CLOCK is suggested to associate with comorbid alcohol use and depressive disorders. J. Circadian Rhythm. 2010, 8, 1. [Google Scholar] [CrossRef]
- De Zavalia, N.; Schoettner, K.; Goldsmith, J.A.; Solis, P.; Ferraro, S.; Parent, G.; Amir, S. Bmal1 in the striatum influences alcohol intake in a sexually dimorphic manner. Commun. Biol. 2021, 4, 1227. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Chen, R.; Lee, Y.; Yoo, S.; Lee, C. Essential roles of CKIdelta and CKIepsilon in the mammalian circadian clock. Proc. Natl. Acad. Sci. USA 2009, 106, 21359–21364. [Google Scholar] [CrossRef]
- Perreau-Lenz, S.; Vengeliene, V.; Noori, H.R.; Merlo-Pich, E.V.; Corsi, M.A.; Corti, C.; Spanagel, R. Inhibition of the casein-kinase-1-ε/δ/ prevents relapse-like alcohol drinking. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2012, 37, 2121–2131. [Google Scholar] [CrossRef]
- Vitaterna, M.H.; Ko, C.H.; Chang, A.M.; Buhr, E.D.; Fruechte, E.M.; Schook, A.; Antoch, M.P.; Turek, F.W.; Takahashi, J.S. The mouse Clock mutation reduces circadian pacemaker amplitude and enhances efficacy of resetting stimuli and phase-response curve amplitude. Proc. Natl. Acad. Sci. USA 2006, 103, 9327–9332. [Google Scholar] [CrossRef]
- Atkinson, S.E.; Maywood, E.S.; Chesham, J.E.; Wozny, C.; Colwell, C.S.; Hastings, M.H.; Williams, S.R. Cyclic AMP Signaling Control of Action Potential Firing Rate and Molecular Circadian Pacemaking in the Suprachiasmatic Nucleus. J. Biol. Rhythm. 2011, 26, 210–220. [Google Scholar] [CrossRef]
- O’Neill, J.S.; Maywood, E.S.; Chesham, J.E.; Takahashi, J.S.; Hastings, M.H. cAMP-Dependent Signaling as a Core Component of the Mammalian Circadian Pacemaker. Science 2008, 320, 949–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Yoo, S.-H.; Park, Y.-S.; Kim, K.-H.; Wei, S.; Buhr, E.; Ye, Z.-Y.; Pan, H.-L.; Takahashi, J.S. Identification of diverse modulators of central and peripheral circadian clocks by high-throughput chemical screening. Proc. Natl. Acad. Sci. USA 2012, 109, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Blednov, Y.A.; Benavidez, J.M.; Black, M.; Harris, R.A. Inhibition of phosphodiesterase 4 reduces ethanol intake and preference in C57BL/6J mice. Front. Neurosci. 2014, 8, 129. [Google Scholar] [CrossRef]
- Blednov, Y.A.; Black, M.; Benavidez, J.M.; Stamatakis, E.E.; Harris, R.A. PPAR Agonists: II. Fenofibrate and Tesaglitazar Alter Behaviors Related to Voluntary Alcohol Consumption. Alcohol. Clin. Exp. Res. 2016, 40, 563–571. [Google Scholar] [CrossRef]
- Grigsby, K.B.; Mangieri, R.A.; Roberts, A.J.; Lopez, M.F.; Tran, A.; Firsick, E.J.; Townsley, K.G.; Beneze, A.; Bess, J.; Eisenstein, T.K.; et al. The FDA-approved drug apremilast suppresses alcohol intake: Clinical and pre-clinical validation. bioRxiv 2021. [Google Scholar] [CrossRef]
- Logrip, M.L. Phosphodiesterase regulation of alcohol drinking in rodents. Alcohol 2015, 49, 795–802. [Google Scholar] [CrossRef]
- Ozburn, A.R.; Metten, P.; Potretzke, S.; Townsley, K.G.; Blednov, Y.A.; Crabbe, J.C. Effects of Pharmacologically Targeting Neuroimmune Pathways on Alcohol Drinking in Mice Selectively Bred to Drink to Intoxication. Alcohol. Clin. Exp. Res. 2020, 44, 553–566. [Google Scholar] [CrossRef]
- Barkley-Levenson, A.M.; Crabbe, J.C. High Drinking in the Dark Mice: A genetic model of drinking to intoxication. Alcohol 2014, 48, 217–223. [Google Scholar] [CrossRef]
- Thiele, T.E.; Navarro, M. “Drinking in the dark” (DID) procedures: A model of binge-like ethanol drinking in non-dependent mice. Alcohol 2014, 48, 235–241. [Google Scholar] [CrossRef]
- Sprow, G.M.; Thiele, T.E. The neurobiology of binge-like ethanol drinking: Evidence from rodent models. Physiol. Behav. 2012, 106, 325–331. [Google Scholar] [CrossRef]
- Banach, E.; Pawlak, J.; Kapelski, P.; Szczepankiewicz, A.; Rajewska-Rager, A.; Skibinska, M.; Czerski, P.; Twarowska-Hauser, J.; Dmitrzak-Weglarz, M. Clock genes polymorphisms in male bipolar patients with comorbid alcohol abuse. J. Affect. Disord. 2018, 241, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Gulick, D.; Gamsby, J.J. Racing the clock: The role of circadian rhythmicity in addiction across the lifespan. Pharmacol. Ther. 2018, 188, 124–139. [Google Scholar] [CrossRef] [PubMed]
- Parekh, P.K.; Ozburn, A.R.; McClung, C.A. Circadian clock genes: Effects on dopamine, reward and addiction. Alcohol 2015, 49, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Grippo, R.M.; Purohit, A.M.; Zhang, Q.; Zweifel, L.S.; Güler, A.D. Direct Midbrain Dopamine Input to the Suprachiasmatic Nucleus Accelerates Circadian Entrainment. Curr. Biol. 2017, 27, 2465–2475.e2463. [Google Scholar] [CrossRef]
- Sidor, M.M.; Spencer, S.M.; Dzirasa, K.; Parekh, P.K.; Tye, K.M.; Warden, M.R.; Arey, R.N.; Enwright, J.F.; Jacobsen, J.P.R.; Kumar, S.; et al. Daytime spikes in dopaminergic activity drive rapid mood-cycling in mice. Mol. Psychiatry 2015, 20, 1406–1419. [Google Scholar] [CrossRef]
- Koike, N.; Yoo, S.-H.; Huang, H.-C.; Kumar, V.; Lee, C.; Kim, T.-K.; Takahashi, J.S. Transcriptional Architecture and Chromatin Landscape of the Core Circadian Clock in Mammals. Science 2012, 338, 349–354. [Google Scholar] [CrossRef]
- Takahashi, J.S. Molecular components of the circadian clock in mammals. Diabetes Obes. Metab. 2015, 17, 6–11. [Google Scholar] [CrossRef]
- Erickson, E.K.; Grantham, E.K.; Warden, A.S.; Harris, R.A. Neuroimmune signaling in alcohol use disorder. Pharm. Biochem. Behav. 2019, 177, 34–60. [Google Scholar] [CrossRef]
- Avchalumov, Y.; Mandyam, C.D. Synaptic Plasticity and its Modulation by Alcohol. Brain Plast. 2020, 6, 103–111. [Google Scholar] [CrossRef]
- Karadayian, A.G.; Lombardi, P.; Bustamante, J.; Lores-Arnaiz, S. Alcohol hangover effects on brain cortex non-synaptic mitochondria and synaptosomes bioenergetics. Alcohol 2019, 77, 113–123. [Google Scholar] [CrossRef]
- Huang, M.C.; Ho, C.W.; Chen, C.H.; Liu, S.C.; Chen, C.C.; Leu, S.J. Reduced expression of circadian clock genes in male alcoholic patients. Alcohol. Clin. Exp. Res. 2010, 34, 1899–1904. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.P.; Kuhn, P.; Advis, J.P.; Sarkar, D.K. Chronic ethanol consumption impairs the circadian rhythm of pro-opiomelanocortin and period genes mRNA expression in the hypothalamus of the male rat. J. Neurochem. 2004, 88, 1547–1554. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Bilbao, A.; Laucht, M.; Henriksson, R.; Yakovleva, T.; Ridinger, M.; Desrivieres, S.; Clarke, T.K.; Lourdusamy, A.; Smolka, M.N.; et al. Effects of the Circadian Rhythm Gene Period 1 (Per1) on Psychosocial Stress-Induced Alcohol Drinking. Am. J. Psychiatry 2011, 168, 1090–1098. [Google Scholar] [CrossRef]
- Crum, R.M.; Storr, C.L.; Chan, Y.F.; Ford, D.E. Sleep Disturbance and Risk for Alcohol-Related Problems. Am. J. Psychiatry 2004, 161, 1197–1203. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Mozhui, K.; Li, Z.; Mulligan, M.K.; Ingels, J.F.; Zhou, X.; Hori, R.T.; Chen, H.; Cook, M.N.; Williams, R.W.; et al. A promoter polymorphism in the Per3 gene is associated with alcohol and stress response. Transl. Psychiatry 2012, 2, e73. [Google Scholar] [CrossRef] [PubMed]
- Zghoul, T.; Abarca, C.; Sanchis-Segura, C.; Albrecht, U.; Schumann, G.; Spanagel, R. Ethanol self-administration and reinstatement of ethanol-seeking behavior in Per1Brdm1mutant mice. Psychopharmacology 2007, 190, 13–19. [Google Scholar] [CrossRef]
- Roybal, K.; Theobold, D.; Graham, A.; DiNieri, J.A.; Russo, S.J.; Krishnan, V.; Chakravarty, S.; Peevey, J.; Oehrlein, N.; Birnbaum, S.; et al. Mania-like behavior induced by disruption of CLOCK. Proc. Natl. Acad. Sci. USA 2007, 104, 6406–6411. [Google Scholar] [CrossRef]
- Mukherjee, S.; Coque, L.; Cao, J.-L.; Kumar, J.; Chakravarty, S.; Asaithamby, A.; Graham, A.; Gordon, E.; Enwright, J.F.; DiLeone, R.J.; et al. Knockdown of Clock in the Ventral Tegmental Area Through RNA Interference Results in a Mixed State of Mania and Depression-Like Behavior. Biol. Psychiatry 2010, 68, 503–511. [Google Scholar] [CrossRef]
- Rizk, A.A.; Jenkins, B.W.; Al-Sabagh, Y.; Hamidullah, S.; Reitz, C.J.; Rasouli, M.; Martino, T.A.; Khokhar, J.Y. The Impact of Sex, Circadian Disruption, and the ClockΔ19/Δ19 Genotype on Alcohol Drinking in Mice. Genes 2022, 13, 701. [Google Scholar] [CrossRef]
- Zhang, R.; Wiers, C.E.; Manza, P.; Tomasi, D.; Shokri-Kojori, E.; Kerich, M.; Almira, E.; Schwandt, M.; Diazgranados, N.; Momenan, R.; et al. Severity of alcohol use disorder influences sex differences in sleep, mood, and brain functional connectivity impairments. Brain Commun. 2022, 4, fcac127. [Google Scholar] [CrossRef]
- Fama, R.; Le Berre, A.-P.; Sullivan, E.V. Alcohol’s unique effects on cognition in women: A 2020 (re)view to envision future research and treatment. Alcohol Res. Curr. Rev. 2020, 40, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Madeira, M.D.; Lieberman, A.R. Sexual dimorphism in the mammalian limbic system. Prog. Neurobiol. 1995, 45, 275–333. [Google Scholar] [CrossRef]
- Forlano, P.M.; Woolley, C.S. Quantitative analysis of pre- and postsynaptic sex differences in the nucleus accumbens. J. Comp. Neurol. 2010, 518, 1330–1348. [Google Scholar] [CrossRef]
- Campi, K.L.; Jameson, C.E.; Trainor, B.C. Sexual Dimorphism in the Brain of the Monogamous California Mouse (Peromyscus californicus). Brain Behav. Evol. 2013, 81, 236–249. [Google Scholar] [CrossRef] [PubMed]
- Pozhidayeva, D.y.Y.; Farris, S.P.; Goeke, C.M.; Firsick, E.J.; Townsley, K.G.; Guizzetti, M.; Ozburn, A.R. Chronic Chemogenetic Stimulation of the Nucleus Accumbens Produces Lasting Reductions in Binge Drinking and Ameliorates Alcohol-Related Morphological and Transcriptional Changes. Brain Sci. 2020, 10, 109. [Google Scholar] [CrossRef] [PubMed]
- Townsley, K.G.; Borrego, M.B.; Ozburn, A.R. Effects of chemogenetic manipulation of the nucleus accumbens core in male C57BL/6J mice. Alcohol 2021, 91, 21–27. [Google Scholar] [CrossRef]
- Cassataro, D.; Bergfeldt, D.; Malekian, C.; Van Snellenberg, J.X.; Thanos, P.K.; Fishell, G.; Sjulson, L. Reverse Pharmacogenetic Modulation of the Nucleus Accumbens Reduces Ethanol Consumption in a Limited Access Paradigm. Neuropsychopharmacology 2014, 39, 283–290. [Google Scholar] [CrossRef]
- Manber, R.; Armitage, R. Sex, Steroids, and Sleep: A Review. Sleep 1999, 22, 540–541. [Google Scholar] [CrossRef]
- Zhou, W.; Cunningham, K.A.; Thomas, M.L. Estrogen regulation of gene expression in the brain: A possible mechanism altering the response to psychostimulants in female rats. Mol. Brain Res. 2002, 100, 75–83. [Google Scholar] [CrossRef]
- Takahashi, J.S. Transcriptional architecture of the mammalian circadian clock. Nat. Rev. Genet. 2017, 18, 164–179. [Google Scholar] [CrossRef]
- Martin-Burgos, B.; Wang, W.; William, I.; Tir, S.; Mohammad, I.; Javed, R.; Smith, S.; Cui, Y.; Arzavala, J.; Mora, D.; et al. Methods for Detecting PER2:LUCIFERASE Bioluminescence Rhythms in Freely Moving Mice. J. Biol. Rhythm. 2022, 37, 78–93. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.-H.; Yamazaki, S.; Lowrey, P.L.; Shimomura, K.; Ko, C.H.; Buhr, E.D.; Siepka, S.M.; Hong, H.-K.; Oh, W.J.; Yoo, O.J.; et al. PERIOD2::LUCIFERASE real-time reporting of circadian dynamics reveals persistent circadian oscillations in mouse peripheral tissues. Proc. Natl. Acad. Sci. USA 2004, 101, 5339–5346. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, J.S.; Best, K.; Belknap, J.K.; Finn, D.A.; Crabbe, J.C. Evaluation of a simple model of ethanol drinking to intoxication in C57BL/6J mice. Physiol. Behav. 2005, 84, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Arey, R.; McClung, C.A. An inhibitor of casein kinase 1 ε/δ partially normalizes the manic-like behaviors of the ClockΔ19 mouse. Behav. Pharm. 2012, 23, 392–396. [Google Scholar] [CrossRef] [PubMed]
- Wager, T.T.; Chandrasekaran, R.Y.; Bradley, J.; Rubitski, D.; Berke, H.; Mente, S.; Butler, T.; Doran, A.; Chang, C.; Fisher, K.; et al. Casein Kinase 1δ/ε Inhibitor PF-5006739 Attenuates Opioid Drug-Seeking Behavior. ACS Chem. Neurosci. 2014, 5, 1253–1265. [Google Scholar] [CrossRef]
- Gamsby, J.J.; Templeton, E.L.; Bonvini, L.A.; Wang, W.; Loros, J.J.; Dunlap, J.C.; Green, A.I.; Gulick, D. The circadian Per1 and Per2 genes influence alcohol intake, reinforcement, and blood alcohol levels. Behav. Brain Res. 2013, 249, 15–21. [Google Scholar] [CrossRef]
- Walker, L.C.; Lawrence, A.J. Investigational drug therapies in phase I and phase II clinical trials for alcohol use disorders. Expert Opin. Investig. Drugs 2018, 27, 677–690. [Google Scholar] [CrossRef]
- Blednov, Y.A.; Da Costa, A.J.; Tarbox, T.; Ponomareva, O.; Messing, R.O.; Harris, R.A. Apremilast Alters Behavioral Responses to Ethanol in Mice: I. Reduced Consumption and Preference. Alcohol. Clin. Exp. Res. 2018, 42, 926–938. [Google Scholar] [CrossRef]
- Vendruscolo, L.F.; Estey, D.; Goodell, V.; Macshane, L.G.; Logrip, M.L.; Schlosburg, J.E.; McGinn, M.A.; Zamora-Martinez, E.R.; Belanoff, J.K.; Hunt, H.J.; et al. Glucocorticoid receptor antagonism decreases alcohol seeking in alcohol-dependent individuals. J. Clin. Investig. 2015, 125, 3193–3197. [Google Scholar] [CrossRef]
- Hasler, B.P.; Bruce, S.; Scharf, D.; Ngari, W.; Clark, D.B. Circadian misalignment and weekend alcohol use in late adolescent drinkers: Preliminary evidence. Chronobiol. Int. 2019, 36, 796–810. [Google Scholar] [CrossRef]
- Adinoff, B.; Krebaum, S.R.; Chandler, P.A.; Ye, W.; Brown, M.B.; Williams, M.J. Dissection of Hypothalamic-Pituitary-Adrenal Axis Pathology in 1-Month-Abstinent Alcohol-Dependent Men, Part 2: Response to Ovine Corticotropin-Releasing Factor and Naloxone. Alcohol. Clin. Exp. Res. 2005, 29, 528–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kidd, T.; Carvalho, L.A.; Steptoe, A. The relationship between cortisol responses to laboratory stress and cortisol profiles in daily life. Biol. Psychol. 2014, 99, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Kakihana, R.; Moore, J.A. Circadian rhythm of corticosterone in mice: The effect of chronic consumption of alcohol. Psychopharmacologia 1976, 46, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Savarese, A.M.; Ozburn, A.R.; Metten, P.; Schlumbohm, J.P.; Hack, W.R.; LeMoine, K.; Hunt, H.; Hausch, F.; Bauder, M.; Crabbe, J.C. Targeting the Glucocorticoid Receptor Reduces Binge-Like Drinking in High Drinking in the Dark (HDID-1) Mice. Alcohol. Clin. Exp. Res. 2020, 44, 1025–1036. [Google Scholar] [CrossRef] [PubMed]
- Savarese, A.M.; Grigsby, K.B.; Jensen, B.E.; Borrego, M.B.; Finn, D.A.; Crabbe, J.C.; Ozburn, A.R. Corticosterone Levels and Glucocorticoid Receptor Gene Expression in High Drinking in the Dark Mice and Their Heterogeneous Stock (HS/NPT) Founder Line. Front. Behav. Neurosci. 2022, 16, 821859. [Google Scholar] [CrossRef]
- McCulley, W.D.; Ascheid, S.; Crabbe, J.C.; Rosenwasser, A.M. Selective breeding for ethanol-related traits alters circadian phenotype. Alcohol 2013, 47, 187–194. [Google Scholar] [CrossRef]
- Crabbe, J.C.; Metten, P.; Belknap, J.K.; Spence, S.E.; Cameron, A.J.; Schlumbohm, J.P.; Huang, L.C.; Barkley-Levenson, A.M.; Ford, M.M.; Phillips, T.J. Progress in a replicated selection for elevated blood ethanol concentrations in HDID mice. Genes Brain Behav. 2014, 13, 236–246. [Google Scholar] [CrossRef]
- Crabbe, J.C.; Metten, P.; Rhodes, J.S.; Yu, C.-H.; Brown, L.L.; Phillips, T.J.; Finn, D.A. A Line of Mice Selected for High Blood Ethanol Concentrations Shows Drinking in the Dark to Intoxication. Biol. Psychiatry 2009, 65, 662–670. [Google Scholar] [CrossRef]
- Jensen, B.E.; Townsley, K.G.; Grigsby, K.B.; Metten, P.; Chand, M.; Uzoekwe, M.; Tran, A.; Firsick, E.; LeBlanc, K.; Crabbe, J.C.; et al. Ethanol-Related Behaviors in Mouse Lines Selectively Bred for Drinking to Intoxication. Brain Sci. 2021, 11, 189. [Google Scholar] [CrossRef]
- Falcon, E.; Ozburn, A.; Mukherjee, S.; Roybal, K.; McClung, C.A. Differential Regulation of the Period Genes in Striatal Regions following Cocaine Exposure. PLoS ONE 2013, 8, e66438. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Descriptive Statistics for Experiment 1 | |||||||
|---|---|---|---|---|---|---|---|
| NAc | Main effects | Clock | Npas2 | Bmal1 | Per1 | Per2 | Per3 |
| Fluid Type | F(1, 80) = 21.61 | F(1, 80) = 18.01 | F(1, 80) = 0.03 | F(1, 80) = 34.67 | F(1, 80) = 7.03 | F(1, 80) = 5.28 | |
| p-value | **** | **** | ns | **** | ** | * | |
| ZT | F(7, 80) = 2.72 | F(7, 80) = 3.22 | F(7, 80) = 7.33 | F(7, 80) = 3.72 | F(7, 80) = 27.64 | F(7, 80) = 6.68 | |
| p-value | * | ** | **** | ** | **** | **** | |
| Interaction | F(7, 80) = 2.83 | F(7, 80) = 1.73 | F(7, 80) = 4.92 | F(7, 80) = 1.37 | F(7, 80) = 2.74 | F(7, 80) = 1.73 | |
| p-value | * | ns | *** | ns | * | ns | |
| VTA | Main effects | Clock | Npas2 | Bmal1 | Per1 | Per2 | Per3 |
| Fluid Type | F(1, 74) = 0.10 | F(1, 74) = 0.25 | F(1, 74) = 0.08 | F(1,70) = 1.68 | F(1,73) = 1.11 | F(1, 74) = 1.85 | |
| p-value | ns | ns | ns | ns | ns | ns | |
| ZT | F(7, 74) = 0.35 | F(7, 74) = 0.86 | F(7, 74) = 1.67 | F(7,70) = 1.62 | F(7,73) = 0.44 | F(7, 74) = 0.85 | |
| p-value | ns | ns | ns | ns | ns | ns | |
| Interaction | F(7, 74) = 1.47 | F(7, 74) = 0.50 | F(7, 74) = 1.22 | F(7,70) = 1.62 | F(7,73) = 0.44 | F(7, 74) = 0.89 | |
| p-value | ns | ns | ns | ns | ns | ns | |
| SCN | Main effects | Clock | Npas2 | Bmal1 | Per1 | Per2 | Per3 |
| Fluid Type | F(1, 76) = 2.78 | F(1, 74) = 2,74 | F(1, 76) = 1.71 | F(1, 74) = 9.08 | F(1, 76) = 12.22 | F(1, 75) = 2.08 | |
| p-value | ns | ns | ns | ** | *** | ns | |
| ZT | F(7, 76) = 2.27 | F(7, 74) = 1.32 | F(7, 76) = 3.15 | F(7, 74) = 3.96 | F(7, 76) = 5.87 | F(7, 80) = 4.98 | |
| p-value | * | ns | ** | *** | **** | *** | |
| Interaction | F(7, 76) = 0.48 | F(7, 76) = 0.93 | F(7, 80) = 1.01 | F(7, 74) = 3.66 | F(7, 76) = 0.84 | F(7, 80) = 0.78 | |
| p-value | ns | ns | ns | ** | ns | ns | |
| Gene | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| 18s | ACCGCAGCTAGGAATAATGGA | GCCTCAGTTCCGAAAACCA |
| Clock | CAGAACAGTACCCAGAGTGCT | CACCACCTGACCCATAAGCAT |
| Npas2 | GACACTGGAGTCCAGACGCAA | AATGTATACAGGGTGCGCCAAA |
| Bmal1 | CCAAGAAAGTATGGACACAGACAAA | GCATTCTTGATCCTTCCTTGGT |
| Per1 | CTCTGTGCTGAAGCAAGACCG | TCATCAGAGTGGCCAGGATCTT |
| Per2 | GAGTGTGTGCAGCGGCTTAG | GTAGGGTGTCATGCGGAAGG |
| Per3 | GTCCATCTGGAGAATGATAGAGCG | GCTTCAGCACCTCCTCTCGAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grigsby, K.; Ledford, C.; Batish, T.; Kanadibhotla, S.; Smith, D.; Firsick, E.; Tran, A.; Townsley, K.; Reyes, K.-A.V.; LeBlanc, K.; et al. Targeting the Maladaptive Effects of Binge Drinking on Circadian Gene Expression. Int. J. Mol. Sci. 2022, 23, 11084. https://doi.org/10.3390/ijms231911084
Grigsby K, Ledford C, Batish T, Kanadibhotla S, Smith D, Firsick E, Tran A, Townsley K, Reyes K-AV, LeBlanc K, et al. Targeting the Maladaptive Effects of Binge Drinking on Circadian Gene Expression. International Journal of Molecular Sciences. 2022; 23(19):11084. https://doi.org/10.3390/ijms231911084
Chicago/Turabian StyleGrigsby, Kolter, Courtney Ledford, Tanvi Batish, Snigdha Kanadibhotla, Delaney Smith, Evan Firsick, Alexander Tran, Kayla Townsley, Kaylee-Abril Vasquez Reyes, Katherine LeBlanc, and et al. 2022. "Targeting the Maladaptive Effects of Binge Drinking on Circadian Gene Expression" International Journal of Molecular Sciences 23, no. 19: 11084. https://doi.org/10.3390/ijms231911084