
Zearalenone Promotes LPS-Induced Oxidative Stress, Endoplasmic Reticulum Stress, and Accelerates Bovine Mammary Epithelial Cell Apoptosis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. ZEA and LPS Reduce MAC-T Cells Viability
2.2. ZEA and LPS Increase LDH in MAC-T Cells
2.3. Combination of ZEA and LPS Causes the Oxidative Stress Response of MAC-T Cells
2.4. The Combination of ZEA and LPS Reduces the Mitochondrial Membrane Potential of MAC-T Cells
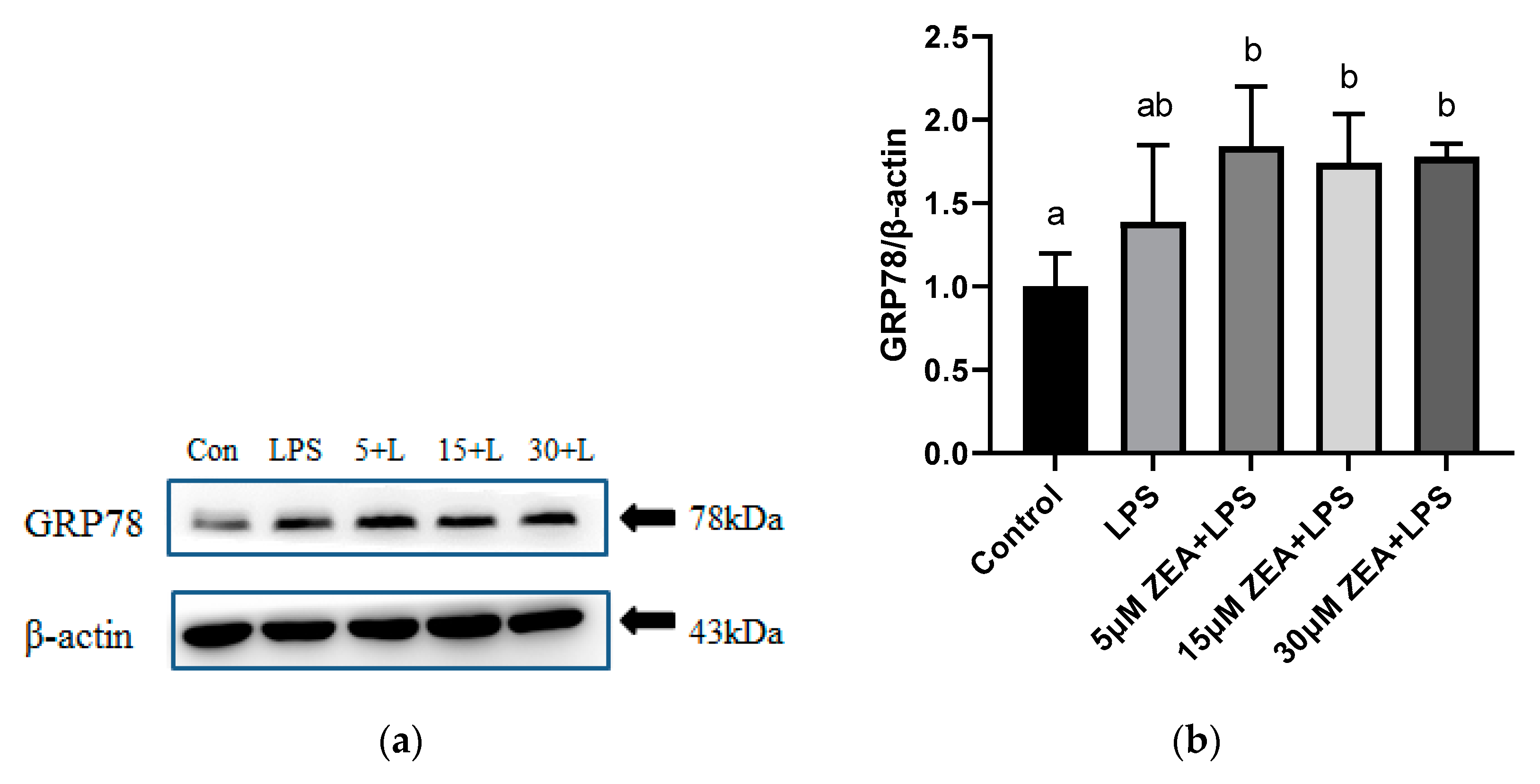
2.5. Combined Treatment of ZEA and LPS Induces ER Stress in MAC-T Cells
2.6. The Combined Effect of ZEA and LPS Accelerates the Apoptosis of MAC-T Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture
4.3. Cell Treatment
4.4. Cell Viability Assay
4.5. Lactate Dehydrogenase Assay
4.6. Antioxidant Enzymes Activities and Malondialdehyde (MDA) Levels
4.7. Measurement of ROS Production
4.8. Mitochondrial Membrane Potential Assay
4.9. Apoptosis Detection by Flow Cytometry
4.10. Western Blotting
4.11. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Winkler, J.; Kersten, S.; Meyer, U.; Stinshoff, H.; Locher, L.; Rehage, J.; Wrenzycki, C.; Engelhardt, U.H.; Dänicke, S. Diagnostic opportunities for evaluation of the exposure of dairy cows to the mycotoxins deoxynivalenol (DON) and zearalenone (ZEN): Reliability of blood plasma, bile and follicular fluid as indicators. J. Anim. Physiol. Anim. Nutr. 2014, 99, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Zada, S.; Alam, S.; Ayoubi, S.A.; Shakeela, Q.; Nisa, S.; Niaz, Z.; Khan, I.; Ahmed, W.; Bibi, Y.; Ahmed, S.; et al. Biological Transformation of Zearalenone by Some Bacterial Isolates Associated with Ruminant and Food Samples. Toxins 2021, 13, 712. [Google Scholar] [CrossRef]
- Hueza, I.; Raspantini, P.; Raspantini, L.; Latorre, A.; Górniak, S. Zearalenone, an Estrogenic Mycotoxin, Is an Immunotoxic Compound. Toxins 2014, 6, 1080–1095. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Tarasiuk, M.; Zielonka, Ł.; Dąbrowski, M.; Nicpoń, J.; Baranowski, M.; Gajęcki, M.T. Changes in the metabolic profile and body weight of pre-pubertal gilts during prolonged monotonic exposure to low doses of zearalenone and deoxynivalenol. Toxicon Off. J. Int. Soc. Toxinol. 2017, 125, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Xiao, Z.H.; Liu, M.; Zhang, N.Y.; Sun, L.H. Dietary Silymarin Supplementation Alleviates Zearalenone-Induced Hepatotoxicity and Reproductive Toxicity in Rats. J. Nutr. 2018, 148, 1209. [Google Scholar] [CrossRef]
- Szabó Odor, J.; Szabó, A.; Kócsó, D.; Marosi, K.; Kovács, M. Interaction between the three frequently co-occurring Fusarium mycotoxins in rats. J. Anim. Physiol. Anim. Nutr. 2019, 103, 370–382. [Google Scholar] [CrossRef] [PubMed]
- Debevere, S.; Baere, S.D.; Haesaert, G.; Rychlik, M.; Croubels, S. Development of an UPLC-MS/MS Method for the Analysis of Mycotoxins in Rumen Fluid with and without Maize Silage Emphasizes the Importance of Using Matrix-Matched Calibration. Toxins 2019, 11, 519. [Google Scholar] [CrossRef] [PubMed]
- Fink-Gremmels, J. The role of mycotoxins in the health and performance of dairy cows. Vet. J. 2008, 176, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Seeling, K.; Dnicke, S.; Valenta, H.; Egmond, H.P.V.; Schothorst, R.C.; Jekel, A.A.; Lebzien, P.; Schollenberger, M.; Razzazi-Fazeli, E.; Flachowsky, G. Effects of Fusarium toxin-contaminated wheat and feed intake level on the biotransformation and carry-over of deoxynivalenol in dairy cows. Food Addit. Contam. 2006, 23, 1008–1020. [Google Scholar] [CrossRef]
- Gruber-Dorninger, C.; Faas, J.; Doupovec, B.; Aleschko, M.; Schatzmayr, D. Metabolism of Zearalenone in the Rumen of Dairy Cows with and without Application of a Zearalenone-Degrading Enzyme. Toxins 2021, 13, 84. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.R.; Jin, Y.C.; Zhao, Y.; Shan, A.S.; Zhang, J. Zearalenone induces apoptosis in bovine mammary epithelial cells by activating endoplasmic reticulum stress. J. Dairy Sci. 2019, 102, 10543–10553. [Google Scholar] [CrossRef] [PubMed]
- Bannert, E.; Tesch, T.; Kluess, J.; Frahm, J.; Kersten, S.; Kahlert, S.; Renner, L.; Rothkötter, H.-J.; Dänicke, S. Metabolic and Hematological Consequences of Dietary Deoxynivalenol Interacting with Systemic Escherichia coli Lipopolysaccharide. Toxins 2015, 7, 4773–4796. [Google Scholar] [CrossRef] [PubMed]
- Xia, D.; Yang, L.; Li, Y.; Chen, J.; Wang, W. Melatonin alleviates Ochratoxin A-induced liver inflammation involved intestinal microbiota homeostasis and intestinal microbiota-independent manner. J. Hazard. Mater. 2021, 413, 125239. [Google Scholar] [CrossRef] [PubMed]
- Bannert, E.; Tesch, T.; Kluess, J.; Winkler, J.; Dnicke, S. On the distribution and metabolism of Fusarium -toxins along the gastrointestinal tract of endotoxaemic pigs. Arch. Anim. Nutr. 2018, 72, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Vels, L.; Røntved, C.M.; Bjerring, M.; Ingvartsen, K.L. Cytokine and acute phase protein gene expression in repeated liver biopsies of dairy cows with a lipopolysaccharide-induced mastitis. J. Dairy Sci. 2009, 92, 922–934. [Google Scholar] [CrossRef]
- Kankkunen, P.; Rintahaka, J.; Aalto, A.; Leino, M.; Majuri, M.L.; Alenius, H.; Wolff, H.; Matikainen, S. Trichothecene mycotoxins activate inflammatory response in human macrophages. J. Immunol. 2009, 182, 6418–6425. [Google Scholar] [CrossRef]
- Lei, G.; Zlab, C.; Glab, C.; Lhab, C.; Xmab, C.; Slab, C.; Dlab, C.; Fang, G.; Khab, C. Nontoxic-dose deoxynivalenol aggravates lipopolysaccharides-induced inflammation and tight junction disorder in IPEC-J2 cells through activation of NF-κB and LC3B-ScienceDirect. Food Chem. Toxicol. 2020, 145. [Google Scholar] [CrossRef]
- Prabhakaran, R.; Kalaivani, P.; Huang, R.; Poornima, P.; Padma, V.V.; Dallemer, F.; Natarajan, K. DNA binding, antioxidant, cytotoxicity (MTT, lactate dehydrogenase, NO), and cellular uptake studies of structurally different nickel(II) thiosemicarbazone complexes: Synthesis, spectroscopy, electrochemistry, and X-ray crystallography. J. Biol. Inorg. Chem. 2013, 18, 233–247. [Google Scholar] [CrossRef]
- Thomas, M.G.; Marwood, R.M.; Parsons, A.E.; Parsons, R.B. The effect of foetal bovine serum supplementation upon the lactate dehydrogenase cytotoxicity assay: Important considerations for in vitro toxicity analysis. Toxicol. Vitr. 2015, 30, 300–308. [Google Scholar] [CrossRef]
- Waller, K.P. Mammary gland immunology around parturition. Influence of stress, nutrition and genetics. Oxyg. Transp. Tissue XXXIII 2000, 480, 231–245. [Google Scholar] [CrossRef]
- So, M.; Tian, Z.; Phoon, Y.; Sha, S.; Antoniou, M.; Zhang, J.; Wu, R.; Tan-Un, K. Gene Expression Profile and Toxic Effects in Human Bronchial Epithelial Cells Exposed to Zearalenone. PLoS ONE 2014, 9, e96404. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Fang, H.; Shen, J.; Jin, Y.; Zhang, J. Curcumin Alleviates LPS-Induced Oxidative Stress, Inflammation and Apoptosis in Bovine Mammary Epithelial Cells via the NFE2L2 Signaling Pathway. Toxins 2021, 13, 208. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Youle, R.J. The role of mitochondria in apoptosis. Annu. Rev. Genet. 2009, 43, 95–118. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Anderson, D.; Dhawan, A. Zinc oxide nanoparticles induce oxidative DNA damage and ROS-triggered mitochondria mediated apoptosis in human liver cells (HepG2). Apoptosis 2012, 17, 852–870. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhang, K.H.; Sun, M.H.; Lan, M.; Sun, S.C. Protective Effects of Melatonin against Zearalenone Toxicity on Porcine Embryos In Vitro. Front. Pharmacol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Sang, Y.; Li, W.; Zhang, G. The protective effect of resveratrol against cytotoxicity induced by mycotoxin, zearalenone. Food Funct. 2016, 7, 3703–3715. [Google Scholar] [CrossRef]
- O’Malley, J.; Kumar, R.; Inigo, J.; Yadava, N.; Chandra, D. Mitochondrial Stress Response and Cancer. Trends Cancer 2020, 6, 688–701. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Jiang, M.; Chen, W.; Zhao, T.; Wei, Y. Cancer and ER stress: Mutual crosstalk between autophagy, oxidative stress and inflammatory response. Biomed. Pharmacother. Biomed. Pharmacother. 2019, 118, 109249. [Google Scholar] [CrossRef]
- Yang, X.; Shao, H.; Liu, W.; Gu, W.; Shu, X.; Mo, Y.; Chen, X.; Zhang, Q.; Jiang, M. Endoplasmic reticulum stress and oxidative stress are involved in ZnO nanoparticle-induced hepatotoxicity. Toxicol. Lett. 2015, 234, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Wei, Y.; Lu, Q.; Qin, D.; Hua, J. Melatonin alleviates LPS-inducedendoplasmic reticulum stress and inflammation in spermatogonial stem cells. J. Cell. Physiol. 2020, 236, 3536–3551. [Google Scholar] [CrossRef]
- Banjerdpongchai, R.; Kongtawelert, P.; Khantamat, O.; Srisomsap, C.; Chokchaichamnankit, D.; Subhasitanont, P.; Svasti, J. Mitochondrial and endoplasmic reticulum stress pathways cooperate in zearalenone-induced apoptosis of human leukemic cells. J. Hematol. Oncol. 2010, 30, 50. [Google Scholar] [CrossRef] [PubMed]
- Swank, G.M.; Qi, L.; Xu, D.Z.; Michalsky, M.; Deitch, E.A. Effect of acute-phase and heat-shock stress on apoptosis in intestinal epithelial cells (Caco-2). Crit. Care Med. 1998, 26, 1213. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.M.; Kubota, H.; Okita, M.; Maeda, T. The anti-inflammatory and antioxidant effects of melatonin on LPS-stimulated bovine mammary epithelial cells. PLoS ONE 2017, 12, e0178525. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, T.; Ren, X.; Li, B.; Wang, S. Male reproductive toxicity of zearalenone-meta-analysis with mechanism review. Ecotoxicol. Environ. Saf. 2021, 221, 112457. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Shen, T.; Ding, Q.; Lv, Y.; Li, L.; Huang, K.; Yan, L.; Song, S. Zearalenone induces ROS-mediated mitochondrial damage in porcine IPEC-J2 cells. J. Biochem. Mol. Toxicol. 2017, 31, 10. [Google Scholar] [CrossRef] [PubMed]
- Taranu, I.; Braicu, C.; Marin, D.E.; Pistol, G.C.; Motiu, M.; Balacescu, L.; Beridan Neagoe, I.; Burlacu, R. Exposure to zearalenone mycotoxin alters in vitro porcine intestinal epithelial cells by differential gene expression. Toxicol. Lett. 2015, 232, 310–325. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.R.; Jin, Y.C.; Shan, A.S.; Zhang, J.; Tian, Y. Polydatin Protects Bovine Mammary Epithelial Cells Against Zearalenone-Induced Apoptosis By Inhibiting Oxidative Responses and Endoplasmic Reticulum Stress. Toxins 2021, 13, 121. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yang, S.; Wang, M.; Chen, J.; Li, P. Curcumin inhibits zearalenone-induced apoptosis and oxidative stress in Leydig cells via modulation of the PTEN/Nrf2/Bip signaling pathway. Food Chem. Toxicol. 2020, 141, 111385. [Google Scholar] [CrossRef] [PubMed]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Y.; Jin, Y.; Tian, Y.; Yu, H.; Wang, R.; Qi, H.; Feng, B.; Zhang, J. Zearalenone Promotes LPS-Induced Oxidative Stress, Endoplasmic Reticulum Stress, and Accelerates Bovine Mammary Epithelial Cell Apoptosis. Int. J. Mol. Sci. 2022, 23, 10925. https://doi.org/10.3390/ijms231810925
Fu Y, Jin Y, Tian Y, Yu H, Wang R, Qi H, Feng B, Zhang J. Zearalenone Promotes LPS-Induced Oxidative Stress, Endoplasmic Reticulum Stress, and Accelerates Bovine Mammary Epithelial Cell Apoptosis. International Journal of Molecular Sciences. 2022; 23(18):10925. https://doi.org/10.3390/ijms231810925
Chicago/Turabian StyleFu, Yurong, Yongcheng Jin, Yue Tian, Hao Yu, Ruqi Wang, Huiyu Qi, Bo Feng, and Jing Zhang. 2022. "Zearalenone Promotes LPS-Induced Oxidative Stress, Endoplasmic Reticulum Stress, and Accelerates Bovine Mammary Epithelial Cell Apoptosis" International Journal of Molecular Sciences 23, no. 18: 10925. https://doi.org/10.3390/ijms231810925