
Examination of the TIGIT-CD226-CD112-CD155 Immune Checkpoint Network during a Healthy Pregnancy
 , , ,
, , ,
Abstract
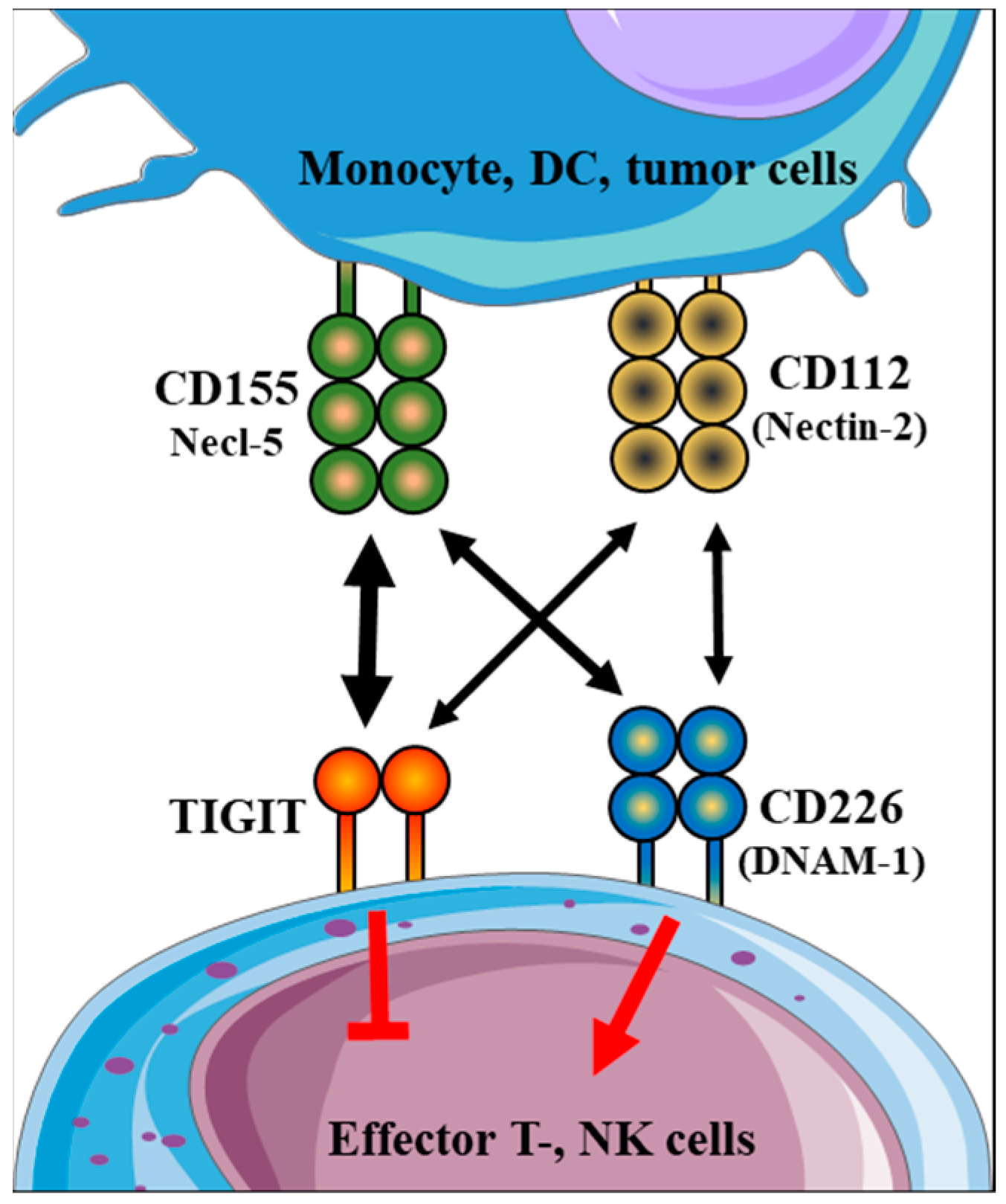
:1. Introduction
2. Results
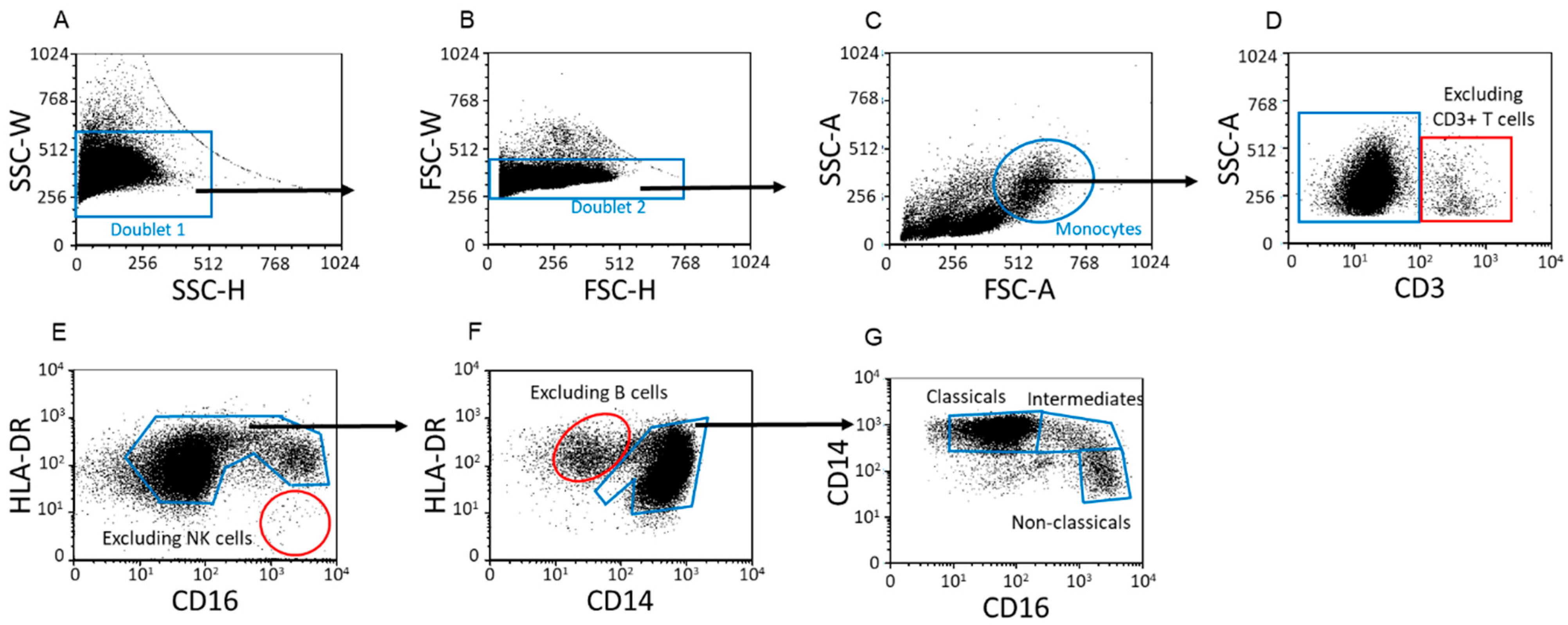
2.1. Distribution of Peripheral Lymphocyte and Monocyte Subpopulations throughout Healthy Pregnancy and in Non-Pregnant Women
2.2. Immune Checkpoint Receptor Expression by Different Immune Cell Subpopulations throughout Healthy Pregnancy and in Non-Pregnant Women
2.3. Immune Checkpoint Ligand Expression by Different Immune Cell Subpopulations throughout Healthy Pregnancy and in Non-Pregnant Women
2.4. Comparing the Percentage of TIGIT−/CD226+ and TIGIT−/CD226− Subpopulations throughout Healthy Pregnancy and in Non-Pregnant Women
2.5. Intracellular Perforin and Granzyme B Content by Immune Cell Subpopulations throughout Healthy Pregnancy and in Non-Pregnant Women
2.6. Circulating CD226, CD112 and CD155 Levels throughout Healthy Pregnancy and in Non-Pregnant Women
2.7. Relationship between the Surface CD226 Receptor Expression by CD4+T and NKT Cell Subpopulations and the Serum Level of CD226 throughout Pregnancy and in Non-Pregnant Women
3. Discussion
4. Materials and Methods
4.1. Participants and Sample Collection
4.2. Lymphocyte Separation, Cryopreservation, and Thawing
4.3. Flow Cytometric Staining and Analysis
4.4. Intracellular Staining
4.5. Enzyme-Linked Immunosorbent Assay (ELISA)
4.6. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Willoughby, J.; Griffiths, J.; Tews, I.; Cragg, M.S. OX40: Structure and function—What questions remain? Mol. Immunol. 2017, 83, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Remedios, K.A.; Zirak, B.; Sandoval, P.M.; Lowe, M.M.; Boda, D.; Henley, E.; Bhattrai, S.; Scharschmidt, T.C.; Liao, W.; Naik, H.B.; et al. The TNFRSF members CD27 and OX40 coordinately limit TH17 differentiation in regulatory T cells. Sci. Immunol. 2018, 3, eaau2042. [Google Scholar] [CrossRef] [PubMed]
- Söderström, L.; Tarnawski, L.; Olofsson, P.S. CD137: A checkpoint regulator involved in atherosclerosis. Atherosclerosis 2018, 272, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Miko, E.; Meggyes, M.; Doba, K.; Barakonyi, A.; Szereday, L. Immune checkpoint molecules in reproductive immunology. Front. Immunol. 2019, 10, 846. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Harden, K.; Gonzalez, L.C.; Francesco, M.; Chiang, E.; Irving, B.; Tom, I.; Ivelja, S.; Refino, C.J.; Clark, H.; et al. The surface protein TIGIT suppresses T cell activation by promoting the generation of mature immunoregulatory dendritic cells. Nat. Immunol. 2009, 10, 48–57. [Google Scholar] [CrossRef]
- Stanietsky, N.; Simic, H.; Arapovic, J.; Toporik, A.; Levy, O.; Novik, A.; Levine, Z.; Beiman, M.; Dassa, L.; Achdout, H.; et al. The interaction of TIGIT with PVR and PVRL2 inhibits human NK cell cytotoxicity. Proc. Natl. Acad. Sci. USA 2009, 106, 17858–17863. [Google Scholar] [CrossRef]
- Joller, N.; Hafler, J.P.; Brynedal, B.; Kassam, N.; Spoerl, S.; Levin, S.D.; Sharpe, A.H.; Kuchroo, V.K. Cutting edge: TIGIT has T cell-intrinsic inhibitory functions. J. Immunol. 2011, 186, 1338–1342. [Google Scholar] [CrossRef]
- Amiezer, M.; Phan, T.G. Disentangling Tfr cells from Treg cells and Tfh cells: How to untie the Gordian knot. Eur. J. Immunol. 2016, 46, 1101–1104. [Google Scholar] [CrossRef]
- Linsley, P.S.; Greene, J.A.L.; Brady, W.; Bajorath, J.; Ledbetter, J.A.; Peach, R. Human B7-1 (CD80) and B7-2 (CD86) bind with similar avidities but distinct kinetics to CD28 and CTLA-4 receptors. Immunity 1994, 1, 793–801. [Google Scholar] [CrossRef]
- He, W.; Zhang, H.; Han, F.; Chen, X.; Lin, R.; Wang, W.; Qiu, H.; Zhuang, Z.; Liao, Q.; Zhang, W.; et al. CD155T/TIGIT Signaling Regulates CD8+ T-cell Metabolism and Promotes Tumor Progression in Human Gastric Cancer. Cancer Res. 2017, 77, 6375–6388. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Qi, G.; Miller, J.S.; Zheng, S.G. CD226: An Emerging Role in Immunologic Diseases. Front. Cell Dev. Biol. 2020, 8, 564. [Google Scholar] [CrossRef] [PubMed]
- Nabekura, T.; Kanaya, M.; Shibuya, A.; Fu, G.; Gascoigne, N.R.J.; Lanier, L.L. Costimulatory molecule DNAM-1 is essential for optimal differentiation of memory natural killer cells during mouse cytomegalovirus infection. Immunity 2014, 40, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Johnston, R.J.; Comps-Agrar, L.; Hackney, J.; Yu, X.; Huseni, M.; Yang, Y.; Park, S.; Javinal, V.; Chiu, H.; Irving, B.; et al. The immunoreceptor TIGIT regulates antitumor and antiviral CD8+ T cell effector function. Cancer Cell 2014, 26, 923–937. [Google Scholar] [CrossRef] [PubMed]
- Mendelsohn, C.L.; Wimmer, E.; Racaniello, V.R. Cellular receptor for poliovirus: Molecular cloning, nucleotide sequence, and expression of a new member of the immunoglobulin superfamily. Cell 1989, 56, 855–865. [Google Scholar] [CrossRef]
- Bevelacqua, V.; Bevelacqua, Y.; Candido, S.; Skarmoutsou, E.; Amoroso, A.; Guarneri, C.; Strazzanti, A.; Gangemi, P.; Mazzarino, M.C.; D’Amico, F.; et al. Nectin like -5 overexpression correlates with the malignant phenotype in cutaneous melanoma. Oncotarget 2012, 3, 882–892. [Google Scholar] [CrossRef]
- Nakai, R.; Maniwa, Y.; Tanaka, Y.; Nishio, W.; Yoshimura, M.; Okita, Y.; Ohbayashi, C.; Satoh, N.; Ogita, H.; Takai, Y.; et al. Overexpression of Necl-5 correlates with unfavorable prognosis in patients with lung adenocarcinoma. Cancer Sci. 2010, 101, 1326–1330. [Google Scholar] [CrossRef]
- Nishiwada, S.; Sho, M.; Yasuda, S.; Shimada, K.; Yamato, I.; Akahori, T.; Kinoshita, S.; Nagai, M.; Konishi, N.; Nakajima, Y. Nectin-4 expression contributes to tumor proliferation, angiogenesis and patient prognosis in human pancreatic cancer. J. Exp. Clin. Cancer Res. 2015, 34, 30. [Google Scholar] [CrossRef]
- Matsumine, A.; Atsumi, S.; Toyoda, H.; Niimi, R.; Iino, T.; Sudo, A. Prognostic significance of CD155 mRNA expression in soft tissue sarcomas. Oncol. Lett. 2013, 5, 1771–1776. [Google Scholar]
- Szereday, L.; Nagy, D.U.; Csiszar, B.; Kevey, D.; Feik, T.; Meggyes, M. Examination of the tigit, cd226, cd112, and cd155 immune checkpoint molecules in peripheral blood mononuclear cells in women diagnosed with early-onset preeclampsia. Biomedicines 2021, 9, 1608. [Google Scholar] [CrossRef]
- Molfetta, R.; Milito, N.D.; Zitti, B.; Lecce, M.; Fionda, C.; Cippitelli, M.; Santoni, A.; Paolini, R. The Ubiquitin-proteasome pathway regulates Nectin2/CD112 expression and impairs NK cell recognition and killing. Eur. J. Immunol. 2019, 49, 873–883. [Google Scholar] [CrossRef]
- Son, Y.S.; Lee, B.N.R.; Choi, Y.J.; Jeon, S.A.; Kim, J.H.; Lee, H.K.; Kwon, S.M.; Cho, J.Y. Nectin-2 (CD112) is expressed on outgrowth endothelial cells and regulates cell proliferation and angiogenic function. PLoS ONE 2016, 11, e0163301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandai, K.; Rikitake, Y.; Mori, M.; Takai, Y. Nectins and nectin-like molecules in development and disease. In Current Topics in Developmental Biology; Academic Press Inc.: Cambridge, MA, USA, 2015; Volume 112, pp. 197–231. [Google Scholar]
- Huang, K.; Lui, W.Y. Nectins and nectin-like molecules (Necls): Recent findings and their role and regulation in spermatogenesis. Semin. Cell Dev. Biol. 2016, 59, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Paniccia, A.; Schulick, A.C.; Chen, W.; Koenig, M.R.; Byers, J.T.; Yao, S.; Bevers, S.; Edil, B.H. Identification of CD112R as a novel checkpoint for human T cells. J. Exp. Med. 2016, 213, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Ho, D.W.H.; Tsui, Y.M.; Chan, L.K.; Sze, K.M.F.; Zhang, X.; Cheu, J.W.S.; Chiu, Y.T.; Lee, J.M.F.; Chan, A.C.Y.; Cheung, E.T.Y.; et al. Single-cell RNA sequencing shows the immunosuppressive landscape and tumor heterogeneity of HBV-associated hepatocellular carcinoma. Nat. Commun. 2021, 12, 3684. [Google Scholar] [CrossRef]
- Whelan, S.; Ophir, E.; Kotturi, M.F.; Levy, O.; Ganguly, S.; Leung, L.; Vaknin, I.; Kumar, S.; Dassa, L.; Hansen, K.; et al. PVRIG and PVRL2 are induced in cancer and inhibit CD8+ T-cell function. Cancer Immunol. Res. 2019, 7, 257–268. [Google Scholar] [CrossRef]
- Kamrani, A.; Soltani-Zangbar, M.S.; Shiri, S.; Yousefzadeh, Y.; Pourakbari, R.; Aghebati-Maleki, L.; Mehdizadeh, A.; Danaii, S.; Jadidi-Niaragh, F.; Yousefi, B.; et al. TIGIT and CD155 as Immune-Modulator Receptor and Ligand on CD4+ T cells in Preeclampsia Patients. Immunol. Investig. 2022, 51, 1023–1038. [Google Scholar] [CrossRef]
- Meggyes, M.; Nagy, D.U.; Szigeti, B.; Csiszar, B.; Sandor, B.; Tamas, P.; Szereday, L. Investigation of mucosal-associated invariant T (MAIT) cells expressing immune checkpoint receptors (TIGIT and CD226) in early-onset preeclampsia. Eur. J. Obstet. Gynecol. Reprod. Biol. 2020, 252, 373–381. [Google Scholar] [CrossRef]
- Zhang, T.; Zhao, Y.; Cheung, W.C.; Gan, Y.H.; Huang, L.; Li, M.; Leung, K.T.; Chung, P.W.; Wang, C.C.; Laird, S.; et al. Serial changes in two immune checkpoint receptors and ligands, Tim-3/Gal-9 and PD-1/PD-L1 in peripheral blood prior to miscarriage: Comparison with pregnancies resulting in a live birth. Am. J. Reprod. Immunol. 2022, 87, e13524. [Google Scholar] [CrossRef]
- Hu, X.H.; Li, Z.H.; Muyayalo, K.P.; Wang, L.L.; Liu, C.Y.; Mor, G.; Liao, A.H. A newly intervention strategy in preeclampsia: Targeting PD-1/Tim-3 signaling pathways to modulate the polarization of decidual macrophages. FASEB J. 2022, 36, e22073. [Google Scholar] [CrossRef]
- Wang, W.; Zhao, Y.; Zhou, X.; Sung, N.; Chen, L.; Zhang, X.; Ma, D.; Zhu, P.; Kwak-Kim, J. Dynamic changes in regulatory T cells during normal pregnancy, recurrent pregnancy loss, and gestational diabetes. J. Reprod. Immunol. 2022, 150, 103492. [Google Scholar] [CrossRef]
- Meggyes, M.; Nagy, D.U.; Szereday, L. Investigation of the PD-1 and PD-L1 Immune Checkpoint Molecules Throughout Healthy Human Pregnancy and in Nonpregnant Women. J. Clin. Med. 2020, 9, 2536. [Google Scholar] [CrossRef] [PubMed]
- Meggyes, M.; Miko, E.; Polgar, B.; Bogar, B.; Farkas, B.; Illes, Z.; Szereday, L. Peripheral blood TIM-3 Positive NK and CD8+ T cells throughout pregnancy: TIM-3/Galectin-9 interaction and its possible role during pregnancy. PLoS ONE 2014, 9, e92371. [Google Scholar] [CrossRef] [PubMed]
- Kühnert, M.; Strohmeier, R.; Stegmüller, M.; Halberstadt, E. Changes in lymphocyte subsets during normal pregnancy. Eur. J. Obstet. Gynecol. Reprod. Biol. 1998, 76, 147–151. [Google Scholar] [CrossRef]
- Coulam, C.B.; Silverfield, J.C.; Kazmar, R.E.; Fathman, C.G. T-Lymphocyte Subsets During Pregnancy and the Menstrual Cycle. Am. J. Reprod. Immunol. 1983, 4, 88–90. [Google Scholar] [CrossRef]
- Pflitsch, C.; Feldmann, C.N.; Richert, L.; Hagen, S.; Diemert, A.; Goletzke, J.; Hecher, K.; Jazbutyte, V.; Renné, T.; Arck, P.C.; et al. In-depth characterization of monocyte subsets during the course of healthy pregnancy. J. Reprod. Immunol. 2020, 141, 103151. [Google Scholar] [CrossRef]
- Ziegler, S.M.; Feldmann, C.N.; Hagen, S.H.; Richert, L.; Barkhausen, T.; Goletzke, J.; Jazbutyte, V.; Martrus, G.; Salzberger, W.; Renné, T.; et al. Innate immune responses to toll-like receptor stimulation are altered during the course of pregnancy. J. Reprod. Immunol. 2018, 128, 30–37. [Google Scholar] [CrossRef]
- Kawanaka, N.; Yamamura, M.; Aita, T.; Morita, Y.; Okamoto, A.; Kawashima, M.; Iwahashi, M.; Ueno, A.; Ohmoto, Y.; Makino, H. CD14+, CD16+ blood monocytes and joint inflammation in rheumatoid arthritis. Arthritis Rheum. 2002, 46, 2578–2586. [Google Scholar] [CrossRef]
- Castaño, D.; García, L.F.; Rojas, M. Increased frequency and cell death of CD16+ monocytes with Mycobacterium tuberculosis infection. Tuberculosis 2011, 91, 348–360. [Google Scholar] [CrossRef]
- Patel, A.A.; Zhang, Y.; Fullerton, J.N.; Boelen, L.; Rongvaux, A.; Maini, A.A.; Bigley, V.; Flavell, R.A.; Gilroy, D.W.; Asquith, B.; et al. The fate and lifespan of human monocyte subsets in steady state and systemic inflammation. J. Exp. Med. 2017, 214, 1913–1923. [Google Scholar] [CrossRef]
- Zawada, A.M.; Rogacev, K.S.; Rotter, B.; Winter, P.; Marell, R.R.; Fliser, D.; Heine, G.H. SuperSAGE evidence for CD14++CD16+ monocytes as a third monocyte subset. Blood 2011, 118, e50–e61. [Google Scholar] [CrossRef]
- Wong, K.L.; Tai, J.J.Y.; Wong, W.C.; Han, H.; Sem, X.; Yeap, W.H.; Kourilsky, P.; Wong, S.C. Gene expression profiling reveals the defining features of the classical, intermediate, and nonclassical human monocyte subsets. Blood 2011, 118, e16–e31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, N.; Sugaya, M.; Suga, H.; Oka, T.; Kawaguchi, M.; Miyagaki, T.; Fujita, H.; Inozume, T.; Sato, S. Increased Soluble CD226 in Sera of Patients with Cutaneous T-Cell Lymphoma Mediates Cytotoxic Activity against Tumor Cells via CD155. J. Investig. Dermatol. 2017, 137, 1766–1773. [Google Scholar] [CrossRef] [PubMed]
- Miko, E.; Barakonyi, A.; Meggyes, M.; Szereday, L. The Role of Type I and Type II NKT Cells in Materno-Fetal Immunity. Biomedicines 2021, 9, 1901. [Google Scholar] [CrossRef] [PubMed]
- Hanasoge Somasundara, A.V.; Moss, M.A.; Feigman, M.J.; Chen, C.; Cyrill, S.L.; Ciccone, M.F.; Trousdell, M.C.; Vollbrecht, M.; Li, S.; Kendall, J.; et al. Parity-induced changes to mammary epithelial cells control NKT cell expansion and mammary oncogenesis. Cell Rep. 2021, 37, 110099. [Google Scholar] [CrossRef]
- Circulating NK and NKT Cells in The Diagnosis and Treatment of Immunological Causes of Female Infertility—Retrospective Data Analysis from The Tertiary Clinical Center—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/33823601/ (accessed on 13 June 2022).











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-Pregnant | 1st Trimester | 2nd Trimester | 3rd Trimester | Significant p-Values | |
|---|---|---|---|---|---|
| CD3+ T cells | 61.27 ± 12.54 | 67.32 ± 9.79 | 68.44 ± 8.08 | 69.24 ± 7.62 | |
| CD4+ T cells | 31.11 ± 9.01 | 36.98 ± 9.80 | 35.98 ± 7.81 | 40.45 ± 7.54 | <0.01 NP vs. 3rd |
| CD4+ T cells in CD3+ T cells | 50.53 ± 12.14 | 54.22 ± 9.44 | 52.71 ± 7.88 | 58.19 ± 8.45 | |
| CD8+ T cells | 25.30 ± 9.01 | 24.32 ± 5.65 | 26.91 ± 4.91 | 24.27 ± 5.50 | |
| CD8+ T cells in CD3+ T cells | 41.18 ± 10.77 | 36.70 ± 8.95 | 39.02 ± 7.06 | 34.97 ± 7.82 | |
| NK cells | 19.03 ± 9.77 | 15.36 ± 7.75 | 12.94 ± 5.18 | 12.93 ± 5.08 | |
| NKdim cells | 17.74 ± 9.12 | 14.00 ± 7.51 | 11.61 ± 5.16 | 11.97 ± 4.94 | |
| NKbright cells | 1.53 ± 2.24 | 1.36 ± 0.74 | 1.34 ± 0.58 | 1.02 ± 0.45 | |
| NKT cells | 6.84 ± 5.84 | 5.51 ± 4.17 | 5.25 ± 3.38 | 4.83 ± 3.63 | |
| Classical monocytes | 89.10 ± 4.37 | 91.35 ± 3.35 | 92.19 ± 2.13 | 93.42 ± 2.52 | <0.03 NP vs. 2nd <0.01 NP vs. 3rd |
| Intermediate monocytes | 2.84 ± 1.56 | 2.80 ± 1.38 | 2.87 ± 1.25 | 2.47 ± 1.13 | |
| Non-classical monocytes | 7.99 ± 3.32 | 5.76 ± 2.64 | 4.82 ± 1.41 | 3.99 ± 1.77 | <0.01 NP vs. 2nd <0.01 NP vs. 3rd |
| Non-Pregnant | 1st Trimester | 2nd Trimester | 3rd Trimester | |
|---|---|---|---|---|
| No. of women | 20 | 26 | 28 | 29 |
| Age (years) | 29.85 (22–43) | 32.77 (18–41) | 32.54 (18–43) | 32.10 (23–44) |
| Gestation age at sampling (weeks) | - | 13.96 ± 2.44 | 24.96 ± 1.82 | 30.86 ± 3.77 |
| Gestation age at birth (weeks) | - | 39.05 ± 1.39 | 38.65 ± 1.87 | 39.34 ± 0.81 |
| Mean of gravidity | - | 0.76 | 0.77 | 0.89 |
| Mean of parity | - | 1.15 | 1.08 | 1.41 |
| Antigen | Fluorochrome | Clone | Isotype | Company | Cat. No. |
|---|---|---|---|---|---|
| CD112 | PE | R2.525 | Mouse IgG1, κ | BD Biosciences | 551057 |
| CD14 | FITC | M5E2 | Mouse IgG2a, κ | BD Biosciences | 555397 |
| CD155 | APC | SKII.4 | Mouse IgG1, κ | Biolegend | 337618 |
| CD16 | PerCp-Cy5.5 | 3G8 | Mouse BALB/c x DBA/2, | BD Biosciences | 560717 |
| CD3 | BV510 | UCHT1 | Mouse BALB/c IgG1, κ | BD Biosciences | 563109 |
| CD4 | FITC | RPA-T4 | Mouse IgG1, κ | BD Biosciences | 555346 |
| CD8 | APC-H7 | SK1 | Mouse BALB/c IgG1, κ | BD Biosciences | 560179 |
| CD56 | PerCp Cy5.5 | B159 | Mouse IgG1, κ | BD Biosciences | 560842 |
| CD56 | APC | B159 | Mouse IgG1, κ | BD Biosciences | 555518 |
| CD226 | BV421 | DX11 | Mouse BALB/c IgG1, κ | BD Biosciences | 742493 |
| Granzyme B | FITC | GB11 | Mouse BALB/c IgG1, κ | BD Biosciences | 560211 |
| HLA-DR | APC-H7 | G46-6 | Mouse IgG2a, κ | BD Biosciences | 561358 |
| NKG2D | PE-Cy7 | 1D11 | Mouse RBF/ DnJ IgG1, κ | BD Biosciences | 562365 |
| Perforin | PE-Cy7 | dG9 | Mouse IgG2b, κ | Biolegend | 308126 |
| TIGIT | PE | A1553G | Mouse IgG2a, κ | Biolegend | 372704 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meggyes, M.; Nagy, D.U.; Feik, T.; Boros, A.; Polgar, B.; Szereday, L. Examination of the TIGIT-CD226-CD112-CD155 Immune Checkpoint Network during a Healthy Pregnancy. Int. J. Mol. Sci. 2022, 23, 10776. https://doi.org/10.3390/ijms231810776
Meggyes M, Nagy DU, Feik T, Boros A, Polgar B, Szereday L. Examination of the TIGIT-CD226-CD112-CD155 Immune Checkpoint Network during a Healthy Pregnancy. International Journal of Molecular Sciences. 2022; 23(18):10776. https://doi.org/10.3390/ijms231810776
Chicago/Turabian StyleMeggyes, Matyas, David U. Nagy, Timoteus Feik, Akos Boros, Beata Polgar, and Laszlo Szereday. 2022. "Examination of the TIGIT-CD226-CD112-CD155 Immune Checkpoint Network during a Healthy Pregnancy" International Journal of Molecular Sciences 23, no. 18: 10776. https://doi.org/10.3390/ijms231810776