
Atomic Simulation of the Binding of JAK1 and JAK2 with the Selective Inhibitor Ruxolitinib
 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
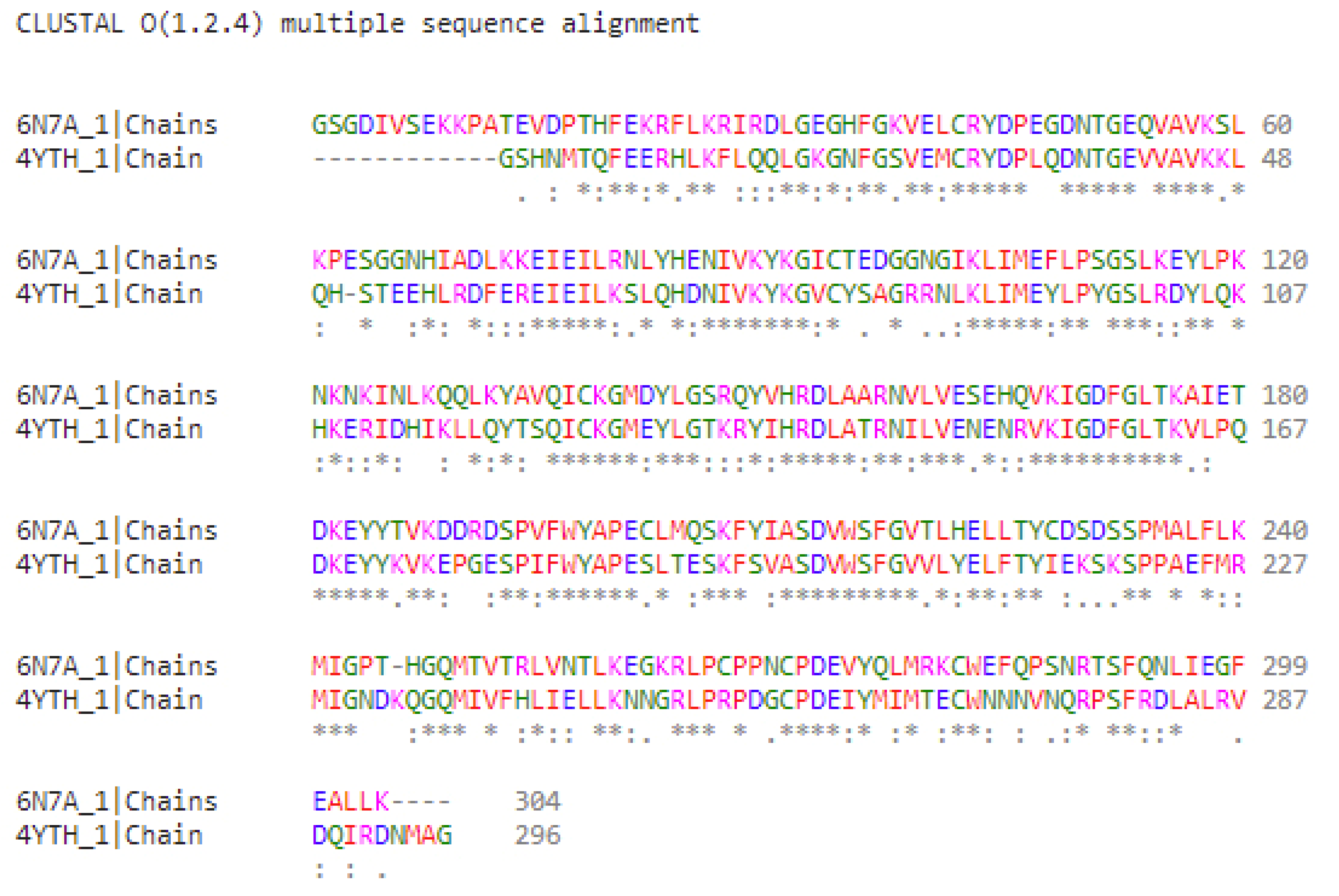
2.1. Characterization of the “Grotto” of Ligand Binding for JAK Isoforms
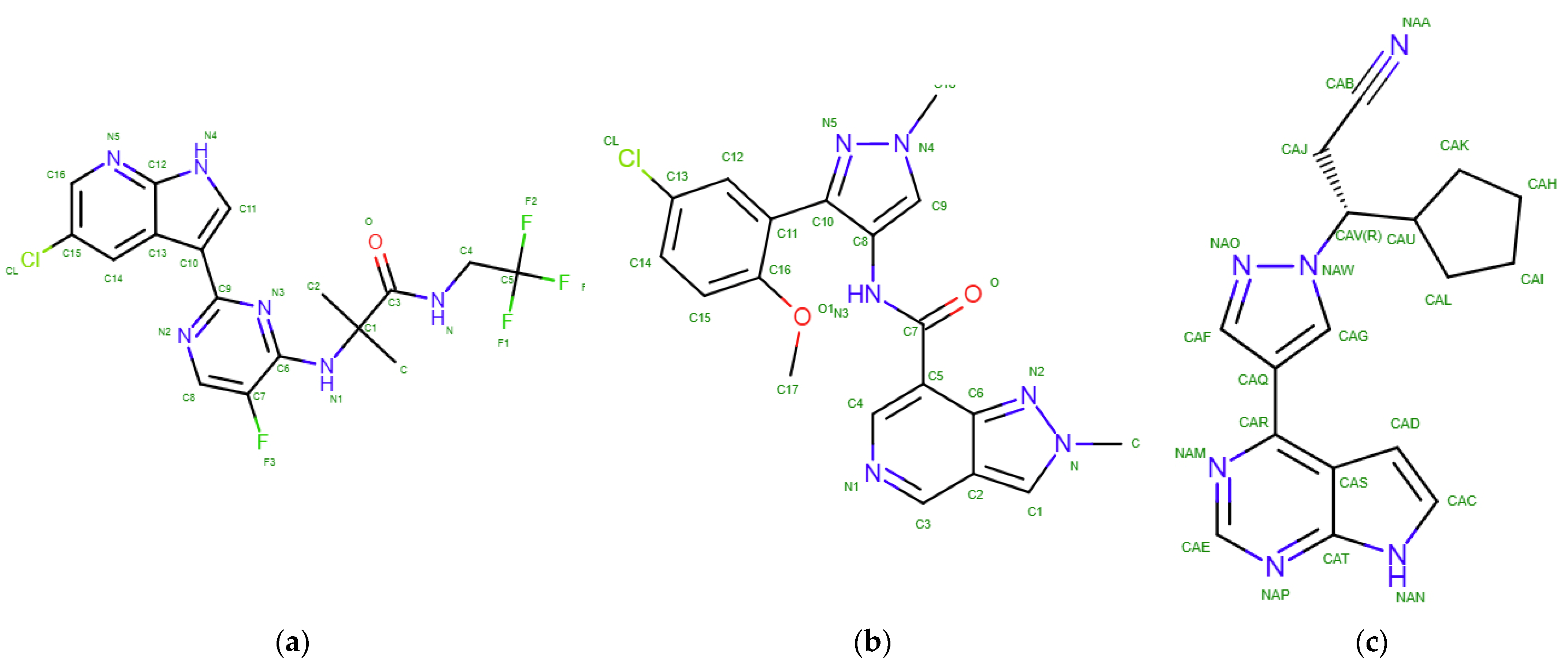
2.2. Results of Molecular Docking
3. Discussion
4. Materials and Methods
4.1. Objects of Study
4.2. Molecular Docking
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sacquin-Mora, S. Fold and Flexibility: What Can Proteins’ Mechanical Properties Tell Us about Their Folding Nucleus? J. R. Soc. Interface 2015, 12, 20150876. [Google Scholar] [CrossRef] [PubMed]
- Khanna, S.; Jaiswal, K.S.; Gupta, B. Managing Rheumatoid Arthritis with Dietary Interventions. Front. Nutr. 2017, 4, 52. [Google Scholar] [CrossRef] [PubMed]
- Malorgio, A.; Malorgio, M.; Benedetti, M.; Casarosa, S.; Cannataro, R. High Intensity Resistance Training as Intervention Method to Knee Osteoarthritis. Sports Med. Health Sci. 2021, 3, 46–48. [Google Scholar] [CrossRef]
- Sandstad, J.; Stensvold, D.; Hoff, M.; Nes, B.M.; Arbo, I.; Bye, A. The Effects of High Intensity Interval Training in Women with Rheumatic Disease: A Pilot Study. Eur. J. Appl. Physiol. 2015, 115, 2081–2089. [Google Scholar] [CrossRef]
- de Jong, Z.; Munneke, M.; Zwinderman, A.H.; Kroon, H.M.; Ronday, K.H.; Lems, W.F.; Dijkmans, B.A.C.; Breedveld, F.C.; Vlieland, T.P.M.V.; Hazes, J.M.W.; et al. Long Term High Intensity Exercise and Damage of Small Joints in Rheumatoid Arthritis. Ann. Rheum. Dis. 2004, 63, 1399–1405. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, D.B.; Willis, L.H.; Slentz, C.A.; Hoselton, A.; Kelly, L.; Huebner, J.L.; Kraus, V.B.; Moss, J.; Muehlbauer, M.J.; Spielmann, G.; et al. Ten Weeks of High-Intensity Interval Walk Training Is Associated with Reduced Disease Activity and Improved Innate Immune Function in Older Adults with Rheumatoid Arthritis: A Pilot Study. Arthritis Res. Ther. 2018, 20, 127. [Google Scholar] [CrossRef]
- Fraser, D.A.; Thoen, J.; Reseland, J.E.; Førre, O.; Kjeldsen-Kragh, J. Decreased CD4+ Lymphocyte Activation and Increased Interleukin-4 Production in Peripheral Blood of Rheumatoid Arthritis Patients after Acute Starvation. Clin. Rheumatol. 1999, 18, 394–401. [Google Scholar] [CrossRef]
- Kjeldsen-Kragh, J.; Haugen, M.; Borchgrevink, C.F.; Laerum, E.; Eek, M.; Mowinkel, P.; Hovi, K.; Førre, O. Controlled Trial of Fasting and One-Year Vegetarian Diet in Rheumatoid Arthritis. Lancet Lond. Engl. 1991, 338, 899–902. [Google Scholar] [CrossRef]
- Michalsen, A.; Riegert, M.; Lüdtke, R.; Bäcker, M.; Langhorst, J.; Schwickert, M.; Dobos, G.J. Mediterranean Diet or Extended Fasting’s Influence on Changing the Intestinal Microflora, Immunoglobulin A Secretion and Clinical Outcome in Patients with Rheumatoid Arthritis and Fibromyalgia: An Observational Study. BMC Complement. Altern. Med. 2005, 5, 22. [Google Scholar] [CrossRef]
- Hafström, I.; Ringertz, B.; Spångberg, A.; von Zweigbergk, L.; Brannemark, S.; Nylander, I.; Rönnelid, J.; Laasonen, L.; Klareskog, L. A Vegan Diet Free of Gluten Improves the Signs and Symptoms of Rheumatoid Arthritis: The Effects on Arthritis Correlate with a Reduction in Antibodies to Food Antigens. Rheumatol. Oxf. Engl. 2001, 40, 1175–1179. [Google Scholar] [CrossRef] [Green Version]
- McDougall, J.; Bruce, B.; Spiller, G.; Westerdahl, J.; McDougall, M. Effects of a Very Low-Fat, Vegan Diet in Subjects with Rheumatoid Arthritis. J. Altern. Complement. Med. 2002, 8, 71–75. [Google Scholar] [CrossRef]
- Abkevich, V.I.; Gutin, A.M.; Shakhnovich, E.I. Specific Nucleus as the Transition State for Protein Folding: Evidence from the Lattice Model. Biochemistry 1994, 33, 10026–10036. [Google Scholar] [CrossRef] [PubMed]
- Dobson, C.M. Protein Folding and Misfolding. Nature 2003, 426, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Faísca, P.F.N. The Nucleation Mechanism of Protein Folding: A Survey of Computer Simulation Studies. J. Phys. Condens. Matter 2009, 21, 373102. [Google Scholar] [CrossRef] [PubMed]
- Kmiecik, S.; Kolinski, A. Folding Pathway of the B1 Domain of Protein G Explored by Multiscale Modeling. Biophys. J. 2008, 94, 726–736. [Google Scholar] [CrossRef]
- Muñoz, V. Conformational Dynamics and Ensembles in Protein Folding. Annu. Rev. Biophys. Biomol. Struct. 2007, 36, 395–412. [Google Scholar] [CrossRef]
- Efimov, A.V. A Novel Super-Secondary Structure of β-Proteins A Triple-Strand Corner. FEBS Lett. 1992, 298, 261–265. [Google Scholar] [CrossRef]
- Efimov, A.V. A Novel Super-Secondary Structure of Proteins and the Relation between the Structure and the Amino Acid Sequence. FEBS Lett. 1984, 166, 33–38. [Google Scholar] [CrossRef]
- Efimov, A.V. Structure of Coiled Beta-Beta-Hairpins and Beta-Beta-Corners. FEBS Lett. 1991, 284, 288–292. [Google Scholar] [CrossRef]
- Chothia, C.; Janin, J. Orthogonal Packing of Beta-Pleated Sheets in Proteins. Biochemistry 1982, 21, 3955–3965. [Google Scholar] [CrossRef]
- Boshkova, E.A.; Efimov, A.V. Structures Closed into Cycles in Proteins Containing 3β-Corners. Biochem. Biokhimiia 2010, 75, 1258–1263. [Google Scholar] [CrossRef] [PubMed]
- Hammarström, A.; Berndt, K.D.; Sillard, R.; Adermann, K.; Otting, G. Solution Structure of a Naturally-Occurring Zinc-Peptide Complex Demonstrates That the N-Terminal Zinc-Binding Module of the Lasp-1 LIM Domain Is an Independent Folding Unit. Biochemistry 1996, 35, 12723–12732. [Google Scholar] [CrossRef] [PubMed]
- Drew, E.D.; Janes, R.W. 2StrucCompare: A Webserver for Visualizing Small but Noteworthy Differences between Protein Tertiary Structures through Interrogation of the Secondary Structure Content. Nucleic Acids Res. 2019, 47, W477–W481. [Google Scholar] [CrossRef] [PubMed]
- Heinig, M.; Frishman, D. STRIDE: A Web Server for Secondary Structure Assignment from Known Atomic Coordinates of Proteins. Nucleic Acids Res. 2004, 32 (Suppl. 2), W500–W502. [Google Scholar] [CrossRef]
- Arnittali, M.; Rissanou, A.N.; Amprazi, M.; Kokkinidis, M.; Harmandaris, V. Structure and Thermal Stability of WtRop and RM6 Proteins through All-Atom Molecular Dynamics Simulations and Experiments. Int. J. Mol. Sci. 2021, 22, 5931. [Google Scholar] [CrossRef]
- Pérez-Alvarado, G.C.; Miles, C.; Michelsen, J.W.; Louis, H.A.; Winge, D.R.; Beckerle, M.C.; Summers, M.F. Structure of the Carboxy-Terminal LIM Domain from the Cysteine Rich Protein CRP. Nat. Struct. Biol. 1994, 1, 388–398. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High Performance Molecular Simulations through Multi-Level Parallelism from Laptops to Supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef]
- Schmid, N.; Eichenberger, A.P.; Choutko, A.; Riniker, S.; Winger, M.; Mark, A.E.; van Gunsteren, W.F. Definition and Testing of the GROMOS Force-Field Versions 54A7 and 54B7. Eur. Biophys. J. 2011, 40, 843. [Google Scholar] [CrossRef]
- Nandel, F.S.; Garla, R. Conformational Behavior of Stereo Regular Substituted Polyglycolides Is Side Chain Dependent. J. Biophys. Chem. 2011, 2, 285–299. [Google Scholar] [CrossRef]
- Bussi, G.; Donadio, D.; Parrinello, M. Canonical Sampling through Velocity Rescaling. J. Chem. Phys. 2007, 126, 014101. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, H.J.C.; Postma, J.P.M.; van Gunsteren, W.F.; DiNola, A.; Haak, J.R. Molecular Dynamics with Coupling to an External Bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef]
- Hess, B.; Bekker, H.; Berendsen, H.; Fraaije, J. LINCS: A Linear Constraint Solver for Molecular Simulations. J. Comput. Chem. 1997, 18, 1463–1472. [Google Scholar] [CrossRef]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A Smooth Particle Mesh Ewald Method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef]
- Daura, X.; Gademann, K.; Jaun, B.; Seebach, D.; van Gunsteren, W.F.; Mark, A.E. Peptide Folding: When Simulation Meets Experiment. Angew. Chem. Int. Ed. 1999, 38, 236–240. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| “Grotto” | SASA, Å2 | HB * | Nonpolar a.a. (%) | Polar a.a. (%) | Charged a.a. (%) | Nonpolar “Cross-Elements” Contacts | Nonpolar “Inside-Elements” Contacts | Distance ** of “Cross-Elements” Contacts (Å ± SD) | Distance of “Inside-Elements” Contacts (Å ± SD) |
|---|---|---|---|---|---|---|---|---|---|
| JAK1 (865–1154) | 29,255.336 | 189 | 36.8 | 30.9 | 31.6 | 310 | 43 | 8.28 ± 1.37 | 7.80 ± 1.54 |
| JAK2 (842-1130) | 31,252.359 | 198 | 38.4 | 30.4 | 30.4 | 333 | 38 | 8.25 ± 1.40 | 7.67 ± 1.62 |
| Ligand | JAK1 (6N7A), kcal/mol | Locus of “Grotto” | HB * | Hydrophobic Bonds ** | JAK2 (4YTH), kcal/mol | Locus of “Grotto” | HB | Hydrophobic Bonds |
|---|---|---|---|---|---|---|---|---|
| Decernotinib | −8.7 | ARG879, LEU881, GLY884, GLY962 | 1 | 10 | −8.7 | LEU855, GLY858, ARG980, LEU983 | 0 | 13 |
| Ligand KEV | −8.7 | ARG879, LEU881, GLY884, GLY962 | 1 | 15 | −8.6 | LEU855, GLY858, ARG980, LEU983 | 1 | 12 |
| Ruxolitinib | −8.3 | ARG879, LEU881, GLY884, GLY962 | 0 | 16 | −8.0 | LEU855, GLY858, ARG980, LEU983 | 0 | 11 |
| Ligand | IUPAC (Formula) | Class | Mw, Da | Donor HB * | Acceptor HB | TPSA **, Å2 | SC 3* | Selectivity | IC50, Nm 4* |
|---|---|---|---|---|---|---|---|---|---|
| Decernotinib | (2R)-2-methyl-2-[[2-(1H-pyrrolo [2,3-b]pyridin-3-yl)pyrimidin-4-yl]amino]-N-(2,2,2-trifluoroethyl)butanamide | Pyrimidine | 392.4 | 3 | 8 | 95.6 | 0 | JAK3 | 2.5 ±0.7 |
| KEV | N-[3-(5-chloro-2-methoxyphenyl)-1-methyl-1H-pyrazol-4-yl]-2-methyl-2H-pyrazolo [4,3-c]pyridine-7-carboxamide(C19H17ClN6O2) | Pyrazole | 396.8 | 1 | 5 | 86.9 | 0 | JAK1 | N/A |
| Ruxolitinib | (3R)-3-cyclopentyl-3-[4-(7H-pyrrolo [2,3-d]pyrimidin-4-yl)pyrazol-1-yl]propanenitrile | Pyrazole | 306.4 | 1 | 4 | 83.2 | 1 | JAK1 and JAK2 | 3.3 ± 1.2 (JAK1)2.8 ± 1.2 (JAK2) |
| Target | PDB ID | Uniprot ID | Resolution, Å | Ligand | Method | Sequence Length, a.a. | Native Ligand | Binding Affinity with Native Ligand, nM |
|---|---|---|---|---|---|---|---|---|
| JAK1 | 6N7A | P23458 | 1.33 | Ruxolitinib | X-ray | 304 | KEV | 4.3 |
| JAK2 | 4YTH | O60674 | 2.04 | Ruxolitinib | X-ray | 296 | Decernotinib | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kondratyev, M.; Rudnev, V.R.; Nikolsky, K.S.; Stepanov, A.A.; Petrovsky, D.V.; Kulikova, L.I.; Kopylov, A.T.; Malsagova, K.A.; Kaysheva, A.L. Atomic Simulation of the Binding of JAK1 and JAK2 with the Selective Inhibitor Ruxolitinib. Int. J. Mol. Sci. 2022, 23, 10466. https://doi.org/10.3390/ijms231810466
Kondratyev M, Rudnev VR, Nikolsky KS, Stepanov AA, Petrovsky DV, Kulikova LI, Kopylov AT, Malsagova KA, Kaysheva AL. Atomic Simulation of the Binding of JAK1 and JAK2 with the Selective Inhibitor Ruxolitinib. International Journal of Molecular Sciences. 2022; 23(18):10466. https://doi.org/10.3390/ijms231810466
Chicago/Turabian StyleKondratyev, Maxim, Vladimir R. Rudnev, Kirill S. Nikolsky, Alexander A. Stepanov, Denis V. Petrovsky, Liudmila I. Kulikova, Arthur T. Kopylov, Kristina A. Malsagova, and Anna L. Kaysheva. 2022. "Atomic Simulation of the Binding of JAK1 and JAK2 with the Selective Inhibitor Ruxolitinib" International Journal of Molecular Sciences 23, no. 18: 10466. https://doi.org/10.3390/ijms231810466