
Peppers in Diet: Genome-Wide Transcriptome and Metabolome Changes in Drosophila melanogaster
 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Phytochemical Content
2.2. Food Intake and Phenotypic Profile of Flies on the Habanero-Pepper Diet
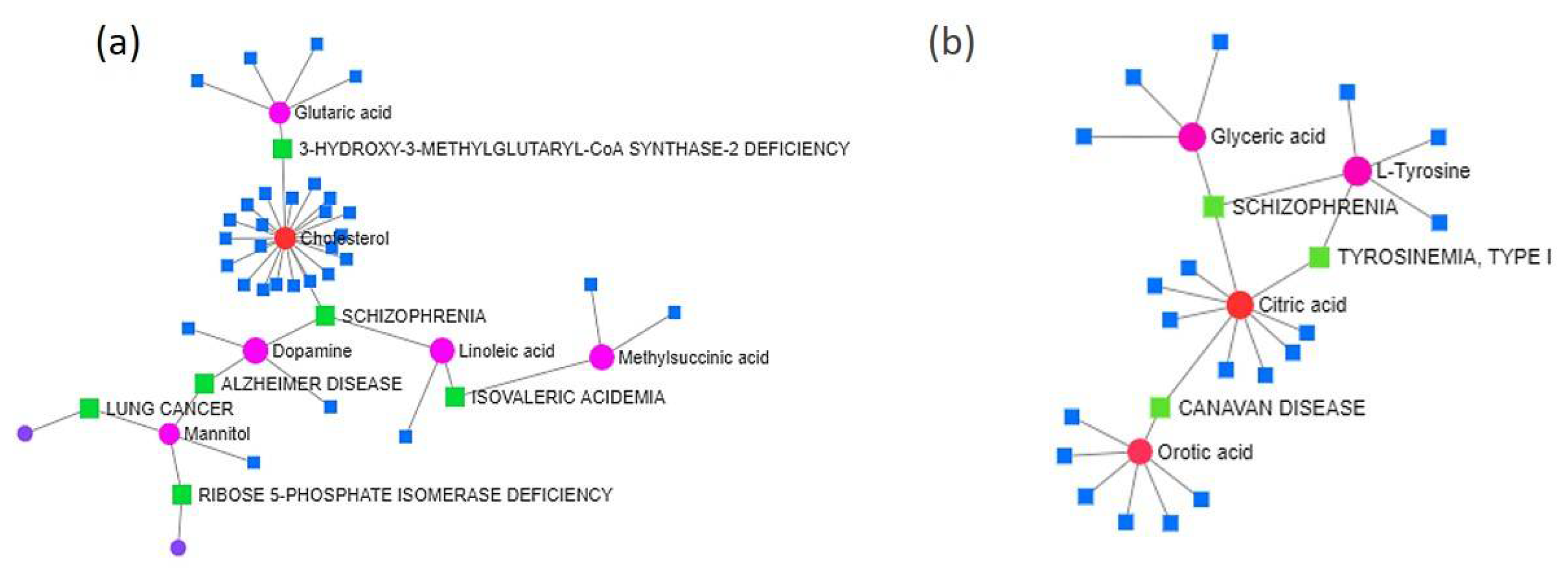
2.3. Metabolome Analysis of Flies on the Habanero-Pepper Diet
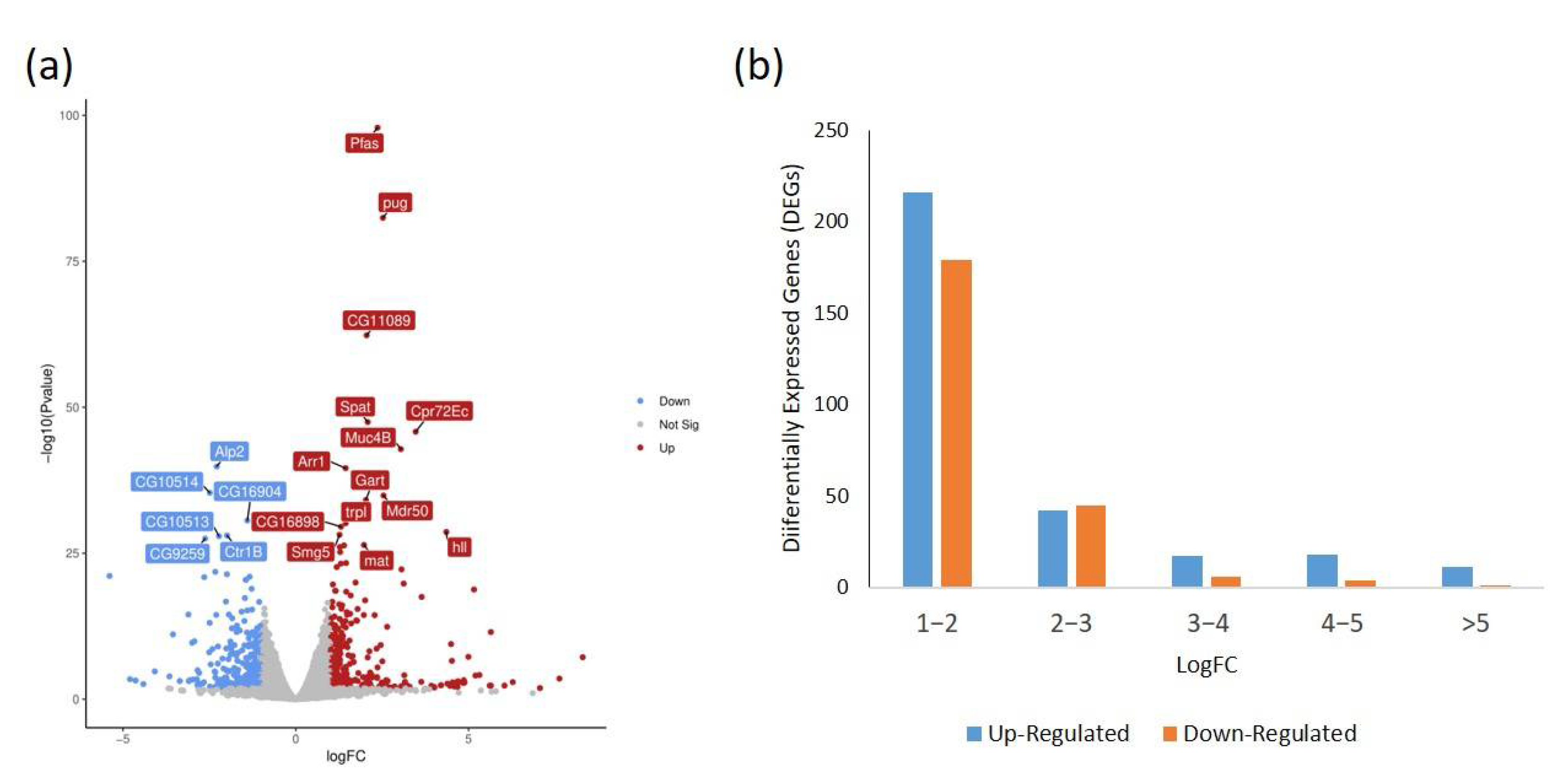
2.4. Transcriptome Analysis of Flies on the Habanero-Pepper Diet
2.5. Functional Annotation and Classification of DEGs
2.6. Pathway Analysis of DEGs
2.7. Functional Network Analysis of DEGs
2.8. DEGs in Response to Capsaicin
2.9. Integrated Metabolomic and Transcriptomic Analyses
3. Discussion
4. Materials and Methods
4.1. Drosophila Stocks and Cultures
4.2. Quantification of Phytochemicals from Habanero Pepper
4.3. Experimental Diets: Control and Habanero-Pepper
4.4. CAFE Assay
4.5. Body Weight Measurement
4.6. Triglycerides (TG) Measurement
4.7. Glucose Measurement
4.8. Metabolome Analysis
4.9. Total RNA Extraction and RNA-Seq Library Construction
4.10. Sequencing Analysis and Identification of Differentially Expressed Genes (DEGs)
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Carpéné, C.; Les, F.; Cásedas, G.; Peiro, C.; Fontaine, J.; Chaplin, A.; Mercader, J.; López, V. Resveratrol anti-obesity effects: Rapid inhibition of adipocyte glucose utilization. Antioxidants 2019, 8, 74. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Zhu, X.; Chen, T.; Peng, B.; Zhang, L.; Liu, X.; Cao, Y.; Chen, Y. Hypolipidemic and anti-obesity effect of nobiletin and its role in prevention of fatty liver in high fat diet-fed rats. Shipin Kexue/Food Sci. 2018, 39, 213–220. [Google Scholar]
- Li, F.; Gao, C.; Yan, P.; Zhang, M.; Wang, Y.; Hu, Y.; Wu, X.; Wang, X.; Sheng, J. EGCG reduces obesity and white adipose tissue gain partly through AMPK activation in mice. Front. Pharmacol. 2018, 9, 1366. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Suo, Y.; Zhang, J.; Zou, Q.; Tan, X.; Yuan, T.; Liu, Z.; Liu, X. Lycopene supplementation attenuates western diet-induced body weight gain through increasing the expressions of thermogenic/mitochondrial functional genes and improving insulin resistance in the adipose tissue of obese mice. J. Nutr. Biochem. 2019, 69, 63–72. [Google Scholar] [CrossRef]
- Andrew, R.; Izzo, A.A. Principles of pharmacological research of nutraceuticals. Br. J. Pharmacol. 2017, 174, 1177. [Google Scholar] [CrossRef]
- Baenas, N.; Belović, M.; Ilic, N.; Moreno, D.; García-Viguera, C. Industrial use of pepper (Capsicum annum L.) derived products: Technological benefits and biological advantages. Food Chem. 2019, 274, 872–885. [Google Scholar] [CrossRef]
- Kraft, K.H.; Brown, C.H.; Nabhan, G.P.; Luedeling, E.; Ruiz, J.D.J.L.; d’Eeckenbrugge, G.C.; Hijmans, R.J.; Gepts, P. Multiple lines of evidence for the origin of domesticated chili pepper, Capsicum annuum, in Mexico. Proc. Natl. Acad. Sci. USA 2014, 111, 6165–6170. [Google Scholar] [CrossRef]
- Nimmakayala, P.; Lopez-Ortiz, C.; Shahi, B.; Abburi, V.L.; Natarajan, P.; Kshetry, A.O.; Shinde, S.; Davenport, B.; Stommel, J.; Reddy, U.K. Exploration into natural variation for genes associated with fruit shape and size among Capsicum chinense collections. Genomics 2021, 113, 3002–3014. [Google Scholar] [CrossRef]
- Richards, B.L.; Whittle, S.L.; Buchbinder, R. Neuromodulators for pain management in rheumatoid arthritis. Cochrane Database Syst. Rev. 2012, 1, CD008921. [Google Scholar] [CrossRef]
- Singh, U.; Bernstein, J.A. Intranasal capsaicin in management of nonallergic (vasomotor) rhinitis. Capsaicin A Ther. Mol. 2014, 68, 147–170. [Google Scholar]
- Perla, V.; Nadimi, M.; Reddy, R.; Hankins, G.R.; Nimmakayala, P.; Harris, R.T.; Valluri, J.; Sirbu, C.; Reddy, U.K. Effect of ghost pepper on cell proliferation, apoptosis, senescence and global proteomic profile in human renal adenocarcinoma cells. PLoS ONE 2018, 13, e0206183. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-P.; Dong, F.-X.; Chai, X.; Zhu, S.; Zhang, B.-L.; Gao, D.-S. Role of autophagy in capsaicin-induced apoptosis in U251 glioma cells. Cell. Mol. Neurobiol. 2016, 36, 737–743. [Google Scholar] [CrossRef]
- Xu, W.; Liu, J.; Ma, D.; Yuan, G.; Lu, Y.; Yang, Y. Capsaicin reduces Alzheimer-associated tau changes in the hippocampus of type 2 diabetes rats. PLoS ONE 2017, 12, e0172477. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Sun, B.-L.; Xiang, Y.; Tian, D.-Y.; Zhu, C.; Li, W.-W.; Liu, Y.-H.; Bu, X.-L.; Shen, L.-L.; Jin, W.-S. Capsaicin consumption reduces brain amyloid-beta generation and attenuates Alzheimer’s disease-type pathology and cognitive deficits in APP/PS1 mice. Transl. Psychiatry 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Zheng, S.; Feng, Q.; Zhang, Q.; Xiao, X. Dietary capsaicin and its anti-obesity potency: From mechanism to clinical implications. Biosci. Rep. 2017, 37, 3. [Google Scholar] [CrossRef] [PubMed]
- Villa-Rivera, M.G.; Ochoa-Alejo, N. Chili Pepper Carotenoids: Nutraceutical Properties and Mechanisms of Action. Molecules 2020, 25, 5573. [Google Scholar] [CrossRef]
- Fontana, L.; Partridge, L. Promoting health and longevity through diet: From model organisms to humans. Cell 2015, 161, 106–118. [Google Scholar] [CrossRef]
- Shih, J.; Hodge, R.; Andrade-Navarro, M.A. Comparison of inter-and intraspecies variation in humans and fruit flies. Genom. Data 2015, 3, 49–54. [Google Scholar] [CrossRef]
- Piya, M.K.; McTernan, P.G.; Kumar, S. Adipokine inflammation and insulin resistance: The role of glucose, lipids and endotoxin. J. Endocrinol. 2013, 216, T1–T15. [Google Scholar] [CrossRef]
- Baenas, N.; Wagner, A.E. Drosophila melanogaster as an alternative model organism in nutrigenomics. Genes Nutr. 2019, 14, 14. [Google Scholar] [CrossRef]
- Evangelakou, Z.; Manola, M.; Gumeni, S.; Trougakos, I.P. Nutrigenomics as a tool to study the impact of diet on aging and age-related diseases: The Drosophila approach. Genes Nutr. 2019, 14, 1–18. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Maleki, M.; Sathyapalan, T.; Jamialahmadi, T.; Sahebkar, A. Obesity and insulin resistance: A review of molecular interactions. Curr. Mol. Med. 2021, 21, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Chow, C.Y.; Reiter, L.T. Etiology of human genetic disease on the fly. Trends Genet. 2017, 33, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.A.; Al-Sakkaf, K.; Shait Mohammed, M.R.; Dallol, A.; Al-Maghrabi, J.; Aldahlawi, A.; Ashoor, S.; Maamra, M.; Ragoussis, J.; Wu, W. Integration of transcriptome and metabolome provides unique insights to pathways associated with obese breast cancer patients. Front. Oncol. 2020, 10, 804. [Google Scholar] [CrossRef] [PubMed]
- de Sá Mendes, N.; de Andrade Gonçalves, É.C.B. The role of bioactive components found in peppers. Trends Food Sci. Technol. 2020, 99, 229–243. [Google Scholar] [CrossRef]
- Whiting, S.; Derbyshire, E.; Tiwari, B. Could capsaicinoids help to support weight management? A systematic review and meta-analysis of energy intake data. Appetite 2014, 73, 183–188. [Google Scholar] [CrossRef]
- Li, Y.; Bai, P.; Wei, L.; Kang, R.; Chen, L.; Zhang, M.; Tan, E.K.; Liu, W. Capsaicin functions as Drosophila ovipositional repellent and causes intestinal dysplasia. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Semaniuk, U.V.; Gospodaryov, D.V.; Strilbytska, O.M.; Kucharska, A.Z.; Sokół-Łętowska, A.; Burdyliuk, N.I.; Storey, K.B.; Bayliak, M.M.; Lushchak, O. Chili pepper extends lifespan in a concentration-dependent manner and confers cold resistance on Drosophila melanogaster cohorts by influencing specific metabolic pathways. Food Funct. 2022, 13, 8313–8328. [Google Scholar] [CrossRef]
- Al-Anzi, B.; Tracey Jr, W.D.; Benzer, S. Response of Drosophila to wasabi is mediated by painless, the fly homolog of mammalian TRPA1/ANKTM1. Curr. Biol. 2006, 16, 1034–1040. [Google Scholar] [CrossRef] [PubMed]
- Baboota, R.K.; Singh, D.P.; Sarma, S.M.; Kaur, J.; Sandhir, R.; Boparai, R.K.; Kondepudi, K.K.; Bishnoi, M. Capsaicin induces “brite” phenotype in differentiating 3T3-L1 preadipocytes. PLoS ONE 2014, 9, e103093. [Google Scholar] [CrossRef]
- Szolcsányi, J. Capsaicin and sensory neurones: A historical perspective. Capsaicin A Ther. Mol. 2014, 68, 1–37. [Google Scholar]
- Chang, A.E.; Vaughan, A.G.; Wilson, R.I. A mechanosensory circuit that mixes opponent channels to produce selectivity for complex stimulus features. Neuron 2016, 92, 888–901. [Google Scholar] [CrossRef] [PubMed]
- Hampel, S.; Eichler, K.; Yamada, D.; Bock, D.D.; Kamikouchi, A.; Seeds, A.M. Distinct subpopulations of mechanosensory chordotonal organ neurons elicit grooming of the fruit fly antennae. Elife 2020, 9, e59976. [Google Scholar] [CrossRef] [PubMed]
- Farhadian, S.F.; Suárez-Fariñas, M.; Cho, C.E.; Pellegrino, M.; Vosshall, L.B. Post-fasting olfactory, transcriptional, and feeding responses in Drosophila. Physiol. Behav. 2012, 105, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Riveron, J.; Boto, T.; Alcorta, E. Transcriptional basis of the acclimation to high environmental temperature at the olfactory receptor organs of Drosophila melanogaster. BMC Genom. 2013, 14, 1–16. [Google Scholar] [CrossRef]
- Chopra, G.; Kaushik, S.; Kain, P. Nutrient Sensing via Gut in Drosophila melanogaster. Int. J. Mol. Sci. 2022, 23, 2694. [Google Scholar] [CrossRef]
- Chatterjee, N.; Perrimon, N. What fuels the fly: Energy metabolism in Drosophila and its application to the study of obesity and diabetes. Sci. Adv. 2021, 7, eabg4336. [Google Scholar] [CrossRef]
- Isabel, G.; Martin, J.-R.; Chidami, S.; Veenstra, J.A.; Rosay, P. AKH-producing neuroendocrine cell ablation decreases trehalose and induces behavioral changes in Drosophila. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2005, 288, R531–R538. [Google Scholar] [CrossRef]
- Gáliková, M.; Diesner, M.; Klepsatel, P.; Hehlert, P.; Xu, Y.; Bickmeyer, I.; Predel, R.; Kühnlein, R.P. Energy homeostasis control in Drosophila adipokinetic hormone mutants. Genetics 2015, 201, 665–683. [Google Scholar] [CrossRef]
- Wat, L.W.; Chao, C.; Bartlett, R.; Buchanan, J.L.; Millington, J.W.; Chih, H.J.; Chowdhury, Z.S.; Biswas, P.; Huang, V.; Shin, L.J. A role for triglyceride lipase brummer in the regulation of sex differences in Drosophila fat storage and breakdown. PLoS Biol. 2020, 18, e3000595. [Google Scholar] [CrossRef]
- Yang, R.; Xue, L.; Zhang, L.; Wang, X.; Qi, X.; Jiang, J.; Yu, L.; Wang, X.; Zhang, W.; Zhang, Q. Phytosterol contents of edible oils and their contributions to estimated phytosterol intake in the Chinese diet. Foods 2019, 8, 334. [Google Scholar] [CrossRef] [PubMed]
- Hannich, J.T.; Umebayashi, K.; Riezman, H. Distribution and functions of sterols and sphingolipids. Cold Spring Harb. Perspect. Biol. 2011, 3, a004762. [Google Scholar] [CrossRef] [PubMed]
- Knittelfelder, O.; Prince, E.; Sales, S.; Fritzsche, E.; Wöhner, T.; Brankatschk, M.; Shevchenko, A. Sterols as dietary markers for Drosophila melanogaster. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2020, 1865, 158683. [Google Scholar] [CrossRef] [PubMed]
- Zanco, B.; Mirth, C.K.; Sgrò, C.M.; Piper, M.D. A dietary sterol trade-off determines lifespan responses to dietary restriction in Drosophila melanogaster females. Elife 2021, 10, e62335. [Google Scholar] [CrossRef] [PubMed]
- Vinci, G.; Xia, X.; Veitia, R.A. Preservation of genes involved in sterol metabolism in cholesterol auxotrophs: Facts and hypotheses. PLoS ONE 2008, 3, e2883. [Google Scholar] [CrossRef]
- Wu, C.; Hwang, S.H.; Jia, Y.; Choi, J.; Kim, Y.-J.; Choi, D.; Pathiraja, D.; Choi, I.-G.; Koo, S.-H.; Lee, S.-J. Olfactory receptor 544 reduces adiposity by steering fuel preference toward fats. J. Clin. Investig. 2017, 127, 4118–4123. [Google Scholar] [CrossRef]
- Xu, H.; Wang, Z.; Zhu, L.; Sui, Z.; Bi, W.; Liu, R.; Bi, K.; Li, Q. Targeted neurotransmitters profiling identifies metabolic signatures in rat brain by LC-MS/MS: Application in insomnia, depression and Alzheimer’s disease. Molecules 2018, 23, 2375. [Google Scholar] [CrossRef]
- Lecis, C.; Segatto, M. Cholesterol homeostasis imbalance and brain functioning: Neurological disorders and behavioral consequences. J. Neurol. Neurol. Disord. 2014, 1, 14. [Google Scholar]
- Sun, Z.; Zhao, L.; Bo, Q.; Mao, Z.; He, Y.; Jiang, T.; Li, Y.; Wang, C.; Li, R. Brain-Specific Oxysterols and Risk of Schizophrenia in Clinical High-Risk Subjects and Patients With Schizophrenia. Front. Psychiatry 2021, 1284. [Google Scholar] [CrossRef]
- Bjerkenstedt, L.; Farde, L.; Terenius, L.; Edman, G.; Venizelos, N.; Wiesel, F.-A. Support for limited brain availability of tyrosine in patients with schizophrenia. Int. J. Neuropsychopharmacol. 2006, 9, 247–255. [Google Scholar] [CrossRef]
- Bilton, D.; Robinson, P.; Cooper, P.; Gallagher, C.; Kolbe, J.; Fox, H.; Jaques, A.; Charlton, B. Inhaled dry powder mannitol in cystic fibrosis: An efficacy and safety study. Eur. Respir. J. 2011, 38, 1071–1080. [Google Scholar] [CrossRef] [PubMed]
- De Jager, R.; Longeval, E.; Klastersky, J. High-Dose Cisplatin With Fluid and Mannitol-Induced Diuresis in Advanced Lung Cancer: A Phase Il Clinical Trial of the EORTC Lung. Cancer Treat. Rep. 1980, 64, 1341–1346. [Google Scholar]
- Rosenbloom, B.E.; Weinreb, N.J. Gaucher disease: A comprehensive review. Crit. Rev.™ Oncog. 2013, 18, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Do, J.; McKinney, C.; Sharma, P.; Sidransky, E. Glucocerebrosidase and its relevance to Parkinson disease. Mol. Neurodegener. 2019, 14, 1–16. [Google Scholar] [CrossRef]
- Davis, M.Y.; Trinh, K.; Thomas, R.E.; Yu, S.; Germanos, A.A.; Whitley, B.N.; Sardi, S.P.; Montine, T.J.; Pallanck, L.J. Glucocerebrosidase deficiency in Drosophila results in α-synuclein-independent protein aggregation and neurodegeneration. PLoS Genet. 2016, 12, e1005944. [Google Scholar] [CrossRef] [Green Version]
- Prentki, M.; Madiraju, S.M. Glycerolipid metabolism and signaling in health and disease. Endocr. Rev. 2008, 29, 647–676. [Google Scholar] [CrossRef]
- Baas-Espinola, F.M.; Castro-Concha, L.A.; Vázquez-Flota, F.A.; Miranda-Ham, M.L. Capsaicin synthesis requires in situ phenylalanine and valine formation in in vitro maintained placentas from Capsicum chinense. Molecules 2016, 21, 799. [Google Scholar] [CrossRef]
- Montoliu, I.; Scherer, M.; Beguelin, F.; DaSilva, L.; Mari, D.; Salvioli, S.; Martin, F.-P.J.; Capri, M.; Bucci, L.; Ostan, R. Serum profiling of healthy aging identifies phospho-and sphingolipid species as markers of human longevity. Aging (Albany NY) 2014, 6, 9. [Google Scholar] [CrossRef] [PubMed]
- Canfield, C.-A.; Bradshaw, P.C. Amino acids in the regulation of aging and aging-related diseases. Transl. Med. Aging 2019, 3, 70–89. [Google Scholar] [CrossRef]
- Alves, A.; Bassot, A.; Bulteau, A.-L.; Pirola, L.; Morio, B. Glycine metabolism and its alterations in obesity and metabolic diseases. Nutrients 2019, 11, 1356. [Google Scholar] [CrossRef]
- Chanchay, P.; Vongsangnak, W.; Thancharoen, A.; Sriboonlert, A. Reconstruction of insect hormone pathways in an aquatic firefly, Sclerotia aquatilis (Coleoptera: Lampyridae), using RNA-seq. PeerJ 2019, 7, e7428. [Google Scholar] [CrossRef] [PubMed]
- Nimmakayala, P.; Abburi, V.L.; Abburi, L.; Alaparthi, S.B.; Cantrell, R.; Park, M.; Choi, D.; Hankins, G.; Malkaram, S.; Reddy, U.K. Linkage disequilibrium and population-structure analysis among Capsicum annuum L. cultivars for use in association mapping. Mol. Genet. Genom. 2014, 289, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Lozano, M.; Haynes, J.; Lopez-Ortiz, C.; Natarajan, P.; Peña-Garcia, Y.; Nimmakayala, P.; Stommel, J.; Alaparthi, S.B.; Sirbu, C.; Balagurusamy, N. Effect of Pepper-Containing Diets on the Diversity and Composition of Gut Microbiome of Drosophila melanogaster. Int. J. Mol. Sci. 2020, 21, 945. [Google Scholar] [CrossRef] [PubMed]
- Diegelmann, S.; Jansen, A.; Jois, S.; Kastenholz, K.; Escarcena, L.V.; Strudthoff, N.; Scholz, H. The CApillary FEeder assay measures food intake in Drosophila melanogaster. JoVE (J. Vis. Exp.) 2017, 121, e55024. [Google Scholar] [CrossRef]
- He, J.; Tan, A.M.X.; Ng, S.Y.; Rui, M.; Yu, F. Cannabinoids modulate food preference and consumption in Drosophila melanogaster. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Heinrichsen, E.T.; Zhang, H.; Robinson, J.E.; Ngo, J.; Diop, S.; Bodmer, R.; Joiner, W.J.; Metallo, C.M.; Haddad, G.G. Metabolic and transcriptional response to a high-fat diet in Drosophila melanogaster. Mol. Metab. 2014, 3, 42–54. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef]
- Liu, T.; Salguero, P.; Petek, M.; Martinez-Mira, C.; Balzano-Nogueira, L.; Ramšak, Ž.; McIntyre, L.; Gruden, K.; Tarazona, S.; Conesa, A. PaintOmics 4: New tools for the integrative analysis of multi-omics datasets supported by multiple pathway databases. Nucleic Acids Res. 2022, 50, W551–W559. [Google Scholar] [CrossRef]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv Prepr. 2013, arXiv:1303.3997. [Google Scholar]
- Li, Q.; Zaim, S.R.; Aberasturi, D.; Berghout, J.; Li, H.; Vitali, F.; Kenost, C.; Zhang, H.H.; Lussier, Y.A. iDEG: A single-subject method utilizing local estimates of dispersion to impute differential expression between two transcriptomes. bioRxiv 2018, 405332. [Google Scholar]
- Götz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Symbol | Differential Expression | Annotation | Human Orthologue | Function in Humans |
|---|---|---|---|---|---|
| FBgn0086693 | iav | [UP] | Encodes a protein involved in the response to different stimuli such as startle, heat, or sound | TRPV1 | TRPV1 Activation in response to heat |
| FBgn0036181 | Muc68Ca | [UP] | Expressed in embryonic/larval midgut; embryonic/larval salivary gland | Muc2 | Protective role for gastrointestinal tissues |
| FBgn0053300 | Muc30E | [UP] | Expressed in amnioserosa; embryonic/larval salivary gland | ||
| FBgn0031273 | CG2839 | [DOWN] | Predicted to enable signaling receptor activity | Reg3g | Modulate intestinal hormones and microbiome |
| FBgn0051661 | CG31661 | [DOWN] | Predicted to enable aspartic-type endopeptidase activity | PGC1-α | Fatty acid metabolism |
| FBgn0004552 | Akh | [UP] | Adipokinetic hormone (Akh) encodes a peptide hormone secreted by the corpora cardiaca | Lipid metabolism | |
| FBgn0263593 | Lpin | [UP] | Encodes a protein that plays a central role in fat body function and energy metabolism | Lpin2 | Lipid and energy metabolism |
| FBgn0033782 | sug | [DOWN] | Encodes a transcription factor that regulates the expression of insulin-like peptides and genes involved in lipid and carbohydrate metabolism | ||
| FBgn0034140 | Lst | [UP] | Encodes a peptide hormone produced by endocrine corpora cardiaca cells during starvation | ||
| FBgn0287184 | FASN3 | [DOWN] | Fatty acid synthase | FASN | Fatty acid metabolism |
| FBgn0051148 | Gba1a | [DOWN] | Enables glucosylceramidase activity, involved in adult locomotory behavior | GBA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopez-Ortiz, C.; Edwards, M.; Natarajan, P.; Pacheco-Valenciana, A.; Nimmakayala, P.; Adjeroh, D.A.; Sirbu, C.; Reddy, U.K. Peppers in Diet: Genome-Wide Transcriptome and Metabolome Changes in Drosophila melanogaster. Int. J. Mol. Sci. 2022, 23, 9924. https://doi.org/10.3390/ijms23179924
Lopez-Ortiz C, Edwards M, Natarajan P, Pacheco-Valenciana A, Nimmakayala P, Adjeroh DA, Sirbu C, Reddy UK. Peppers in Diet: Genome-Wide Transcriptome and Metabolome Changes in Drosophila melanogaster. International Journal of Molecular Sciences. 2022; 23(17):9924. https://doi.org/10.3390/ijms23179924
Chicago/Turabian StyleLopez-Ortiz, Carlos, Mary Edwards, Purushothaman Natarajan, Armando Pacheco-Valenciana, Padma Nimmakayala, Donald A. Adjeroh, Cristian Sirbu, and Umesh K. Reddy. 2022. "Peppers in Diet: Genome-Wide Transcriptome and Metabolome Changes in Drosophila melanogaster" International Journal of Molecular Sciences 23, no. 17: 9924. https://doi.org/10.3390/ijms23179924