
Osteogenic Efficacy of Human Trophoblasts-Derived Conditioned Medium on Mesenchymal Stem Cells
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
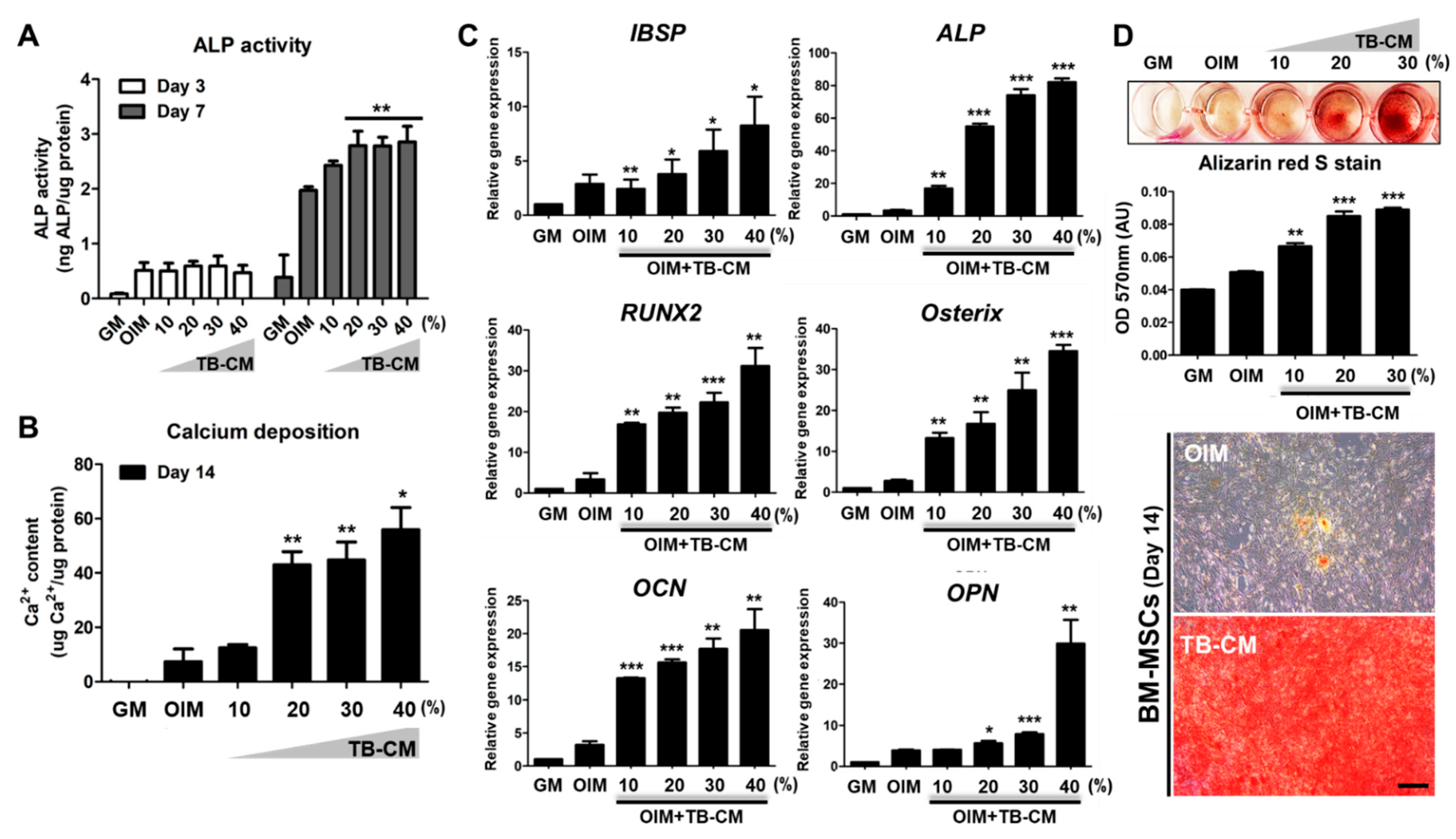
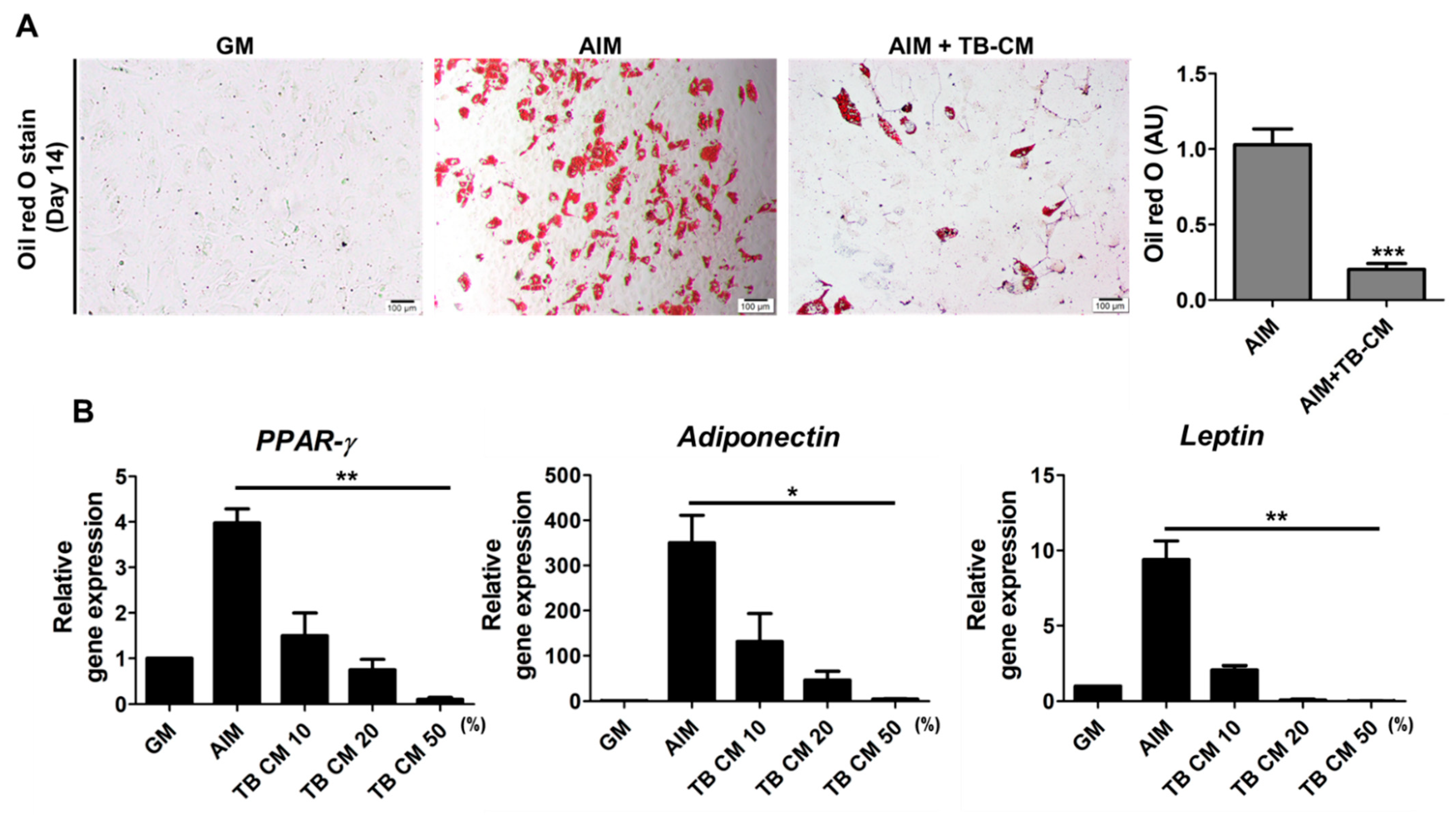
2.1. TB-CM Promotes Osteogenesis but Inhibits Adipogenesis in Human MSCs
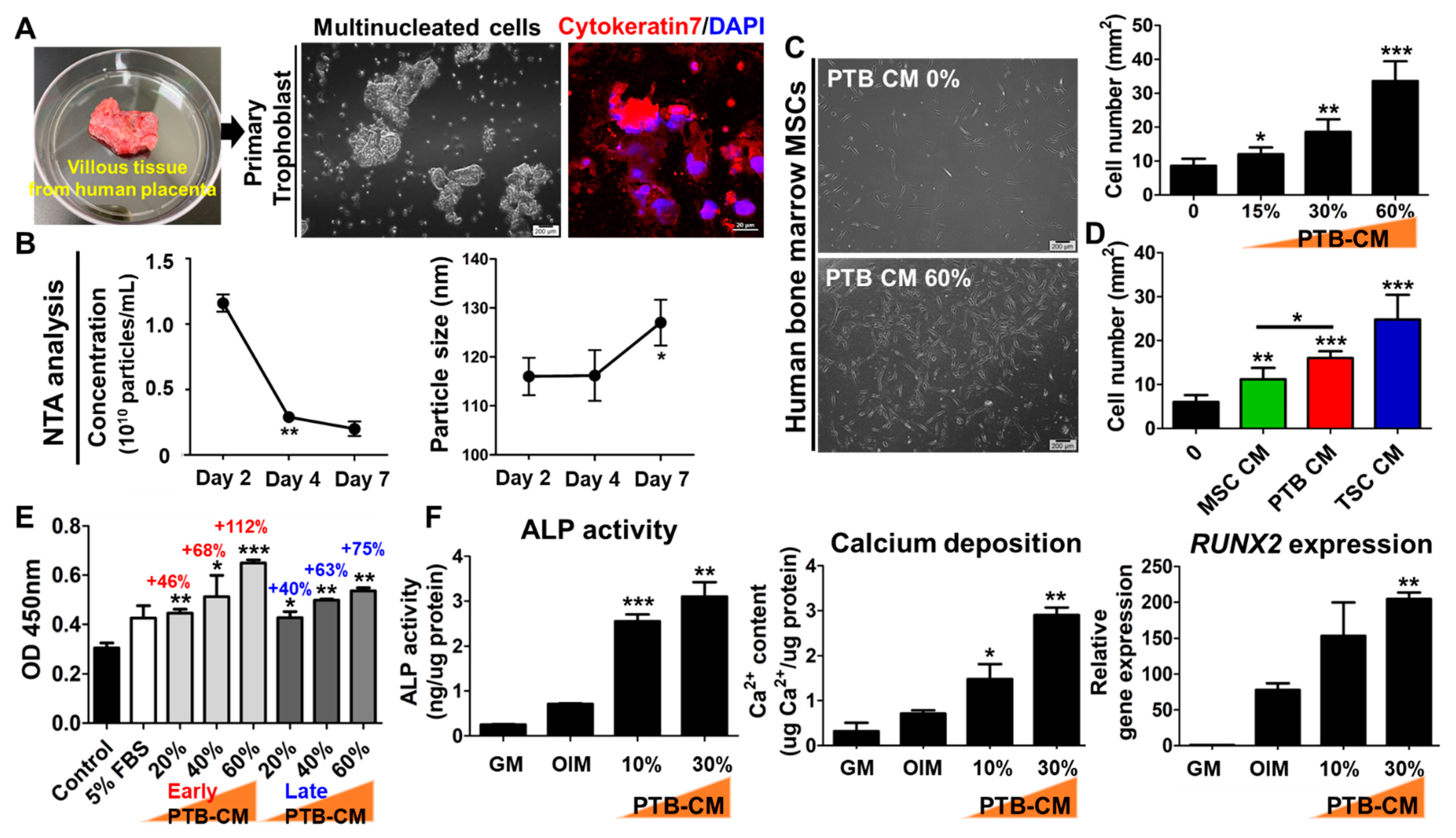
2.2. Human PTB-CM Promotes Proliferation and Osteogenic Differentiation of MSCs
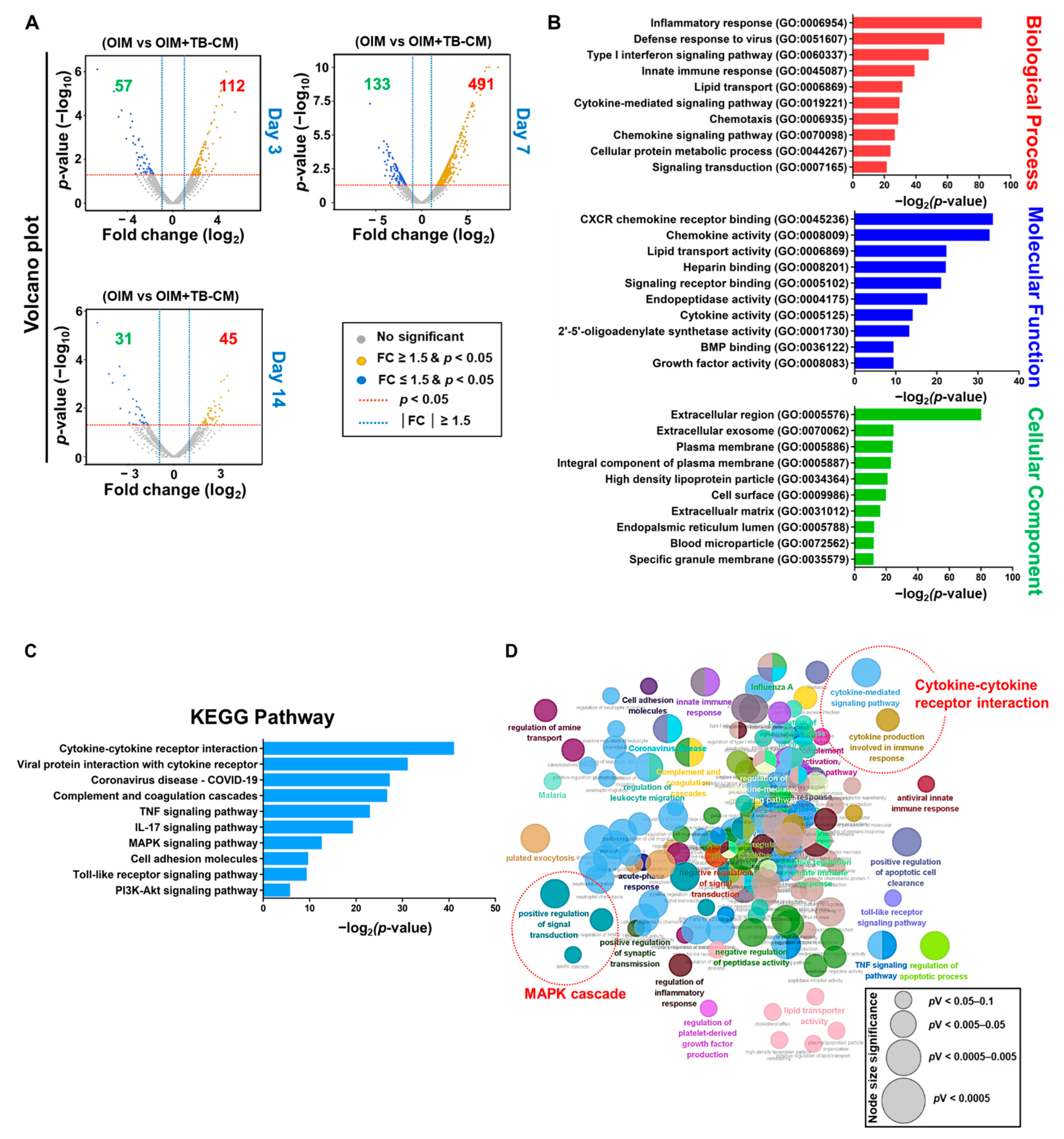
2.3. Transcriptomic Analysis of the Role of TB-CM during Osteogenesis of MSCs
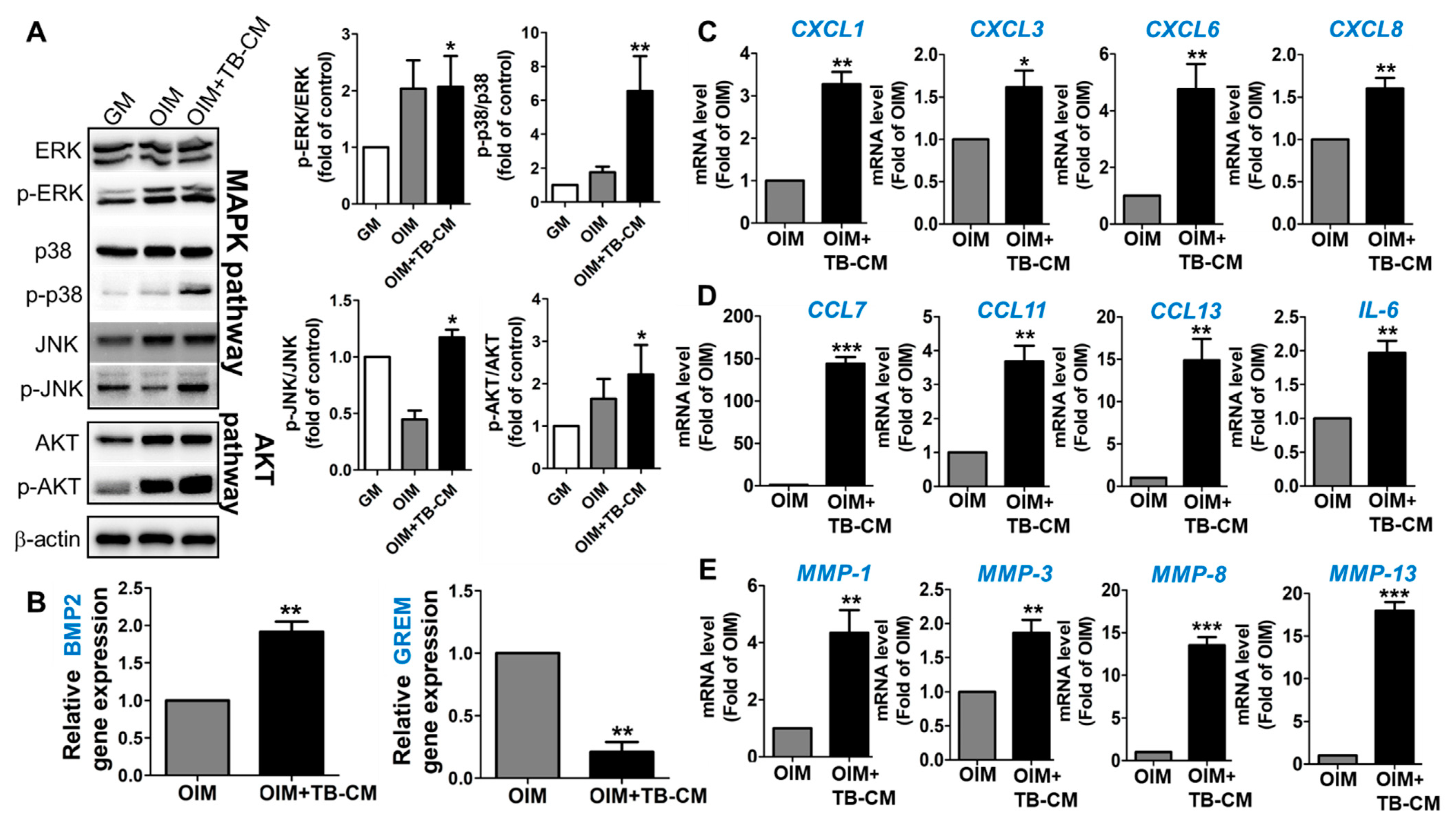
2.4. TB-CM Regulates the MAPK-, BMP2-, and Cytokine-Mediated Cellular Processes during the Osteogenesis of MSCs
3. Discussion
4. Materials and Methods
4.1. Cell Culture Procedure and In Vitro Osteogenic Differentiation
4.2. Preparation of PTB-CM
4.3. Alkaline Phosphatase (ALP) Assay
4.4. Calcium Assay and Alizarin Red S Staining
4.5. CCK8 Assay
4.6. In Vitro Adipogenic Differentiation
4.7. Nanoparticle Tracking Analysis (NTA)
4.8. Immunofluorescence Staining
4.9. RNA Extraction and Quantitative Real-Time RT-PCR
4.10. Western Blot Analysis
4.11. RNA-Seq
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ATCC | American Type Culture Collection |
| ALP | alkaline phosphatase |
| ARE | AU-rich element |
| AUF1 | ARE/poly(U)-binding/degradation factor 1 |
| BMP2 | bone morphogenetic protein 2 |
| CM | conditioned medium |
| DAVID | Database for Annotation, Visualization, and Integrated Discovery |
| DEGs | differentially expressed genes |
| ERK | extracellular signal-regulated kinase |
| ECM | extracellular matrix |
| EVs | extracellular vesicles |
| FBS | fetal bovine serum |
| GO | Gene ontology |
| GREM1 | Gremlin-1 |
| hMSCs | human mesenchymal stem cells |
| IL-17 | interleukin-17 |
| JNK | c-JUN N-terminal kinase |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| MAPK | mitogen-activated protein kinase |
| MMP | matrix metalloproteinase |
| MSCs | mesenchymal stem cells |
| OCN | osteocalcin |
| OPN | osteopontin |
| OSX | osterix |
| OIM | osteogenic induction medium |
| PPAR-g | peroxisome proliferator-activated receptor-gamma |
| PI3K- AKT | phosphatidylinositol 3-kinase (PI3K)-protein kinase B (Akt) |
| PTM-CM | primary trophoblast-derived conditioned medium |
| RNA-seq | RNA-sequencing |
| TBs | trophoblasts |
| TB-CM | trophoblast-derived conditioned medium |
| TGF-β | transforming growth factor-beta |
| TSCs | trophoblast stem cells |
| TNF | tumor necrosis factor |
References
- Albrektsson, T.; Johansson, C. Osteoinduction, osteoconduction and osseointegration. Eur. Spine J. 2001, 10 (Suppl. 2), S96–S101. [Google Scholar]
- Rutkovskiy, A.; Stensløkken, K.-O.; Vaage, I.J. Osteoblast Differentiation at a Glance. Med. Sci. Monit. Basic Res. 2016, 22, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Luvizuto, E.R.; Tangl, S.; Zanoni, G.; Okamoto, T.; Sonoda, C.; Gruber, R.; Okamoto, R. The effect of BMP-2 on the osteoconductive properties of β-tricalcium phosphate in rat calvaria defects. Biomaterials 2011, 32, 3855–3861. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Enggist, L.; Kuffer, A.F.; Buser, D.; Hunziker, E.B. The influence of BMP-2 and its mode of delivery on the osteoconductivity of implant surfaces during the early phase of osseointegration. Biomaterials 2007, 28, 2677–2686. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Harris, M.A.; Rossini, G.; Dunstan, C.R.; Dallas, S.L.; Feng, J.Q.; Mundy, G.R.; Harris, S.E. Bone morphogenetic protein 2 (BMP-2) enhances BMP-3, BMP-4, and bone cell differen-tiation marker gene expression during the induction of mineralized bone matrix formation in cultures of fetal rat calvarial os-teoblasts. Calcif. Tissue Int. 1997, 60, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Long, F. Building strong bones: Molecular regulation of the osteoblast lineage. Nat. Rev. Mol. Cell. Biol. 2011, 13, 27–38. [Google Scholar] [CrossRef] [PubMed]
- James, A.W.; Lachaud, G.; Shen, J.; Asatrian, G.; Nguyen, V.; Zhang, X.; Ting, K.; Soo, C. A Review of the Clinical Side Effects of Bone Morphogenetic Protein-2. Tissue Eng. Part B Rev. 2016, 22, 284–297. [Google Scholar] [CrossRef]
- Chen, Y.-T.; Tsai, M.-J.; Hsieh, N.; Lo, M.-J.; Lee, M.-J.; Cheng, H.; Huang, W.-C. The superiority of conditioned medium derived from rapidly expanded mesenchymal stem cells for neural repair. Stem Cell Res. Ther. 2019, 10, 390. [Google Scholar] [CrossRef]
- Zhong, S.; He, X.; Li, Y.; Lou, X. Conditioned Medium Enhances Osteogenic Differentiation of Induced Pluripotent Stem Cell-Derived Mesenchymal Stem Cells. Tissue Eng. Regen. Med. 2019, 16, 141–150. [Google Scholar] [CrossRef]
- Oh, M.; Lee, J.; Kim, Y.J.; Rhee, W.J.; Park, J.H. Exosomes Derived from Human Induced Pluripotent Stem Cells Ameliorate the Aging of Skin Fibroblasts. Int. J. Mol. Sci. 2018, 19, 1715. [Google Scholar] [CrossRef]
- Shang, F.; Yu, Y.; Liu, S.; Ming, L.; Zhang, Y.; Zhou, Z.; Zhao, J.; Jin, Y. Advancing application of mesenchymal stem cell-based bone tissue regeneration. Bioact. Mater. 2020, 6, 666–683. [Google Scholar] [CrossRef] [PubMed]
- Re, F.; Gabusi, E.; Manferdini, C.; Russo, D.; Lisignoli, G. Bone Regeneration Improves with Mesenchymal Stem Cell Derived Ex-tracellular Vesicles (EVs) Combined with Scaffolds: A Systematic Review. Biology 2021, 10, 579. [Google Scholar] [CrossRef]
- Benavides-Castellanos, M.P.; Garzón-Orjuela, N.; Linero, I. Effectiveness of mesenchymal stem cell-conditioned medium in bone regeneration in animal and human models: A systematic review and meta-analysis. Cell Regen. 2020, 9, 5. [Google Scholar] [CrossRef]
- Ross, C.; Boroviak, T.E. Origin and function of the yolk sac in primate embryogenesis. Nat. Commun. 2020, 11, 3760. [Google Scholar] [CrossRef]
- Mukherjee, I.; Dhar, R.; Singh, S.; Sharma, J.B.; Nag, T.C.; Mridha, A.R.; Jaiswal, P.; Biswas, S.; Karmakar, S. Oxidative stress-induced impairment of trophoblast function causes preeclampsia through the unfolded protein response pathway. Sci. Rep. 2021, 11, 18415. [Google Scholar] [CrossRef]
- Karahoda, R.; Zaugg, J.; Fuenzalida, B.; Kallol, S.; Moser-Haesing, R.; Staud, F.; Albrecht, C. Trophoblast Differentiation Affects Crucial Nutritive Functions of Placental Mem-brane Transporters. Front. Cell Dev. Biol. 2022, 10, 820286. [Google Scholar] [CrossRef]
- Ouyang, Y.; Bayer, A.; Chu, T.; Tyurin, V.A.; Kagan, V.E.; Morelli, A.E.; Coyne, C.B.; Sadovsky, Y. Isolation of human trophoblastic extracellular vesicles and characterization of their cargo and antiviral activity. Placenta 2016, 47, 86–95. [Google Scholar] [CrossRef]
- Farhadihosseinabadi, B.; Farahani, M.; Tayebi, T.; Jafari, A.; Biniazan, F.; Modaresifar, K.; Moravvej, H.; Bahrami, S.; Redl, H.; Tayebi, L.; et al. Amniotic membrane and its epithelial and mesenchymal stem cells as an appropriate source for skin tissue engineering and regenerative medicine. Artif. Cells Nanomed. Biotechnol. 2018, 46 (Suppl. 2), 431–440. [Google Scholar] [CrossRef]
- Malhotra, C.; Jain, A.K. Human amniotic membrane transplantation: Different modalities of its use in ophthalmology. World J. Transplant. 2014, 4, 111–121. [Google Scholar] [CrossRef]
- Go, Y.Y.; Chae, S.; Song, J. Comprehensive analysis of human chorionic membrane extracts regulating mesenchymal stem cells during osteogenesis. Cell Prolif. 2021, 55, e13160. [Google Scholar] [CrossRef]
- Go, Y.Y.; Kim, S.E.; Cho, G.J.; Chae, S.-W.; Song, J.-J. Differential effects of amnion and chorion membrane extracts on osteoblast-like cells due to the different growth factor composition of the extracts. PLoS ONE 2017, 12, e0182716. [Google Scholar] [CrossRef] [Green Version]
- Go, Y.Y.; Lee, C.M.; Ju, W.M.; Chae, S.W.; Song, J.J. Extracellular Vesicles (Secretomes) from Human Trophoblasts Promote the Re-generation of Skin Fibroblasts. Int. J. Mol. Sci. 2021, 22, 6959. [Google Scholar] [CrossRef]
- Rodríguez-Carballo, E.; Gámez, B.; Ventura, F. p38 MAPK Signaling in Osteoblast Differentiation. Front. Cell Dev. Biol. 2016, 4, 40. [Google Scholar] [CrossRef] [PubMed]
- Tsang, E.J.; Wu, B.; Zuk, P. MAPK signaling has stage-dependent osteogenic effects on human adipose-derived stem cells in vitro. Connect. Tissue Res. 2017, 59, 129–146. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Rotwein, P. Akt promotes BMP2-mediated osteoblast differentiation and bone development. J. Cell Sci. 2009, 122, 716–726. [Google Scholar] [CrossRef] [PubMed]
- Baker, N.; Sohn, J.; Tuan, R.S. Promotion of human mesenchymal stem cell osteogenesis by PI3-kinase/Akt signaling, and the influence of caveolin-1/cholesterol homeostasis. Stem Cell Res. Ther. 2015, 6, 238. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Shou, P.; Zheng, C.; Jiang, M.; Cao, G.; Yang, Q.; Cao, J.; Xie, N.; Velletri, T.; Zhang, X.; et al. Fate decision of mesenchymal stem cells: Adipocytes or osteoblasts? Cell Death Differ. 2016, 23, 1128–1139. [Google Scholar] [CrossRef]
- Teitelbaum, S.L. Bone resorption by osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef]
- Justesen, J.; Stenderup, K.; Ebbesen, E.; Mosekilde, L.; Steiniche, T.; Kassem, M. Adipocyte tissue volume in bone marrow is increased with aging and in patients with osteoporosis. Biogerontology 2001, 2, 165–171. [Google Scholar] [CrossRef]
- Feehan, K.T.; Gilroy, D.W. Is Resolution the End of Inflammation? Trends Mol. Med. 2019, 25, 198–214. [Google Scholar] [CrossRef]
- Zhang, N.; Cui, M.; Liu, X.; Liu, L.; Yu, L.; Zhao, X.; Cao, L.; Yuanyuan, J. IL-17F promotes osteoblastic osteogenesis via the MAPK/ERK1/2 signaling pathway. Exp. Ther. Med. 2021, 22, 1052. [Google Scholar] [CrossRef]
- Moerman, E.J.; Teng, K.; Lipschitz, D.A.; Lecka-Czernik, B. Aging activates adipogenic and suppresses osteogenic programs in mesenchymal marrow stroma/stem cells: The role of PPAR-gamma2 transcription factor and TGF-beta/BMP signaling path-ways. Aging Cell 2004, 3, 379–389. [Google Scholar] [CrossRef]
- Croes, M.; Oner, F.C.; Kruyt, M.C.; Blokhuis, T.J.; Bastian, O.; Dhert, W.; Alblas, J. Proinflammatory Mediators Enhance the Osteogenesis of Human Mesenchymal Stem Cells after Lineage Commitment. PLoS ONE 2015, 10, e0132781. [Google Scholar] [CrossRef]
- Brylka, L.J.; Schinke, T. Chemokines in Physiological and Pathological Bone Remodeling. Front. Immunol. 2019, 10, 2182. [Google Scholar] [CrossRef]
- Fu, R.; Liu, H.; Zhao, S.; Wang, Y.; Li, L.; Gao, S.; Ruan, E.; Wang, G.; Wang, H.; Song, J.; et al. Osteoblast inhibition by chemokine cytokine ligand3 in myeloma-induced bone disease. Cancer Cell Int. 2014, 14, 132. [Google Scholar] [CrossRef]
- Maruyama, M.; Rhee, C.; Utsunomiya, T.; Zhang, N.; Ueno, M.; Yao, Z.; Goodman, S.B. Modulation of the Inflammatory Response and Bone Healing. Front. Endocrinol. 2020, 11, 386. [Google Scholar] [CrossRef]
- Toosi, S.; Behravan, J. Osteogenesis and bone remodeling: A focus on growth factors and bioactive peptides. BioFactors 2019, 46, 326–340. [Google Scholar] [CrossRef]
- Croes, M.; Kruyt, M.C.; Groen, W.M.; Van Dorenmalen, K.M.A.; Dhert, W.; Öner, F.C.; Alblas, J. Interleukin 17 enhances bone morphogenetic protein-2-induced ectopic bone formation. Sci. Rep. 2018, 8, 7269. [Google Scholar] [CrossRef]
- Han, X.; Yang, Q.; Lin, L.; Xu, C.; Zheng, C.; Chen, X.; Han, Y.; Li, M.; Cao, W.; Cao, K.; et al. Interleukin-17 enhances immunosuppression by mesenchymal stem cells. Cell Death Differ. 2014, 21, 1758–1768. [Google Scholar] [CrossRef]
- Chen, J.; Cao, J.; Luo, Y.; Xie, L.; Song, J.; Xue, W.; Jia, R.; Song, J. Expression of ERK and p-ERK proteins of ERK signaling pathway in the kidneys of fluoride-exposed carp (Cyprinus carpio). Acta Histochem. 2014, 116, 1337–1341. [Google Scholar] [CrossRef]
- Huang, D.; Li, X.; Sun, L.; Huang, P.; Ying, H.; Wang, H.; Wu, J.; Song, H. Regulation of Hippo signalling by p38 signalling. J. Mol. Cell Biol. 2016, 8, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Paiva, K.B.; Granjeiro, J.M. Matrix Metalloproteinases in Bone Resorption, Remodeling, and Repair. Prog. Mol. Biol. Transl. Sci. 2017, 148, 203–303. [Google Scholar] [CrossRef] [PubMed]
- Almalki, S.G.; Agrawal, D.K. Effects of matrix metalloproteinases on the fate of mesenchymal stem cells. Stem Cell Res. Ther. 2016, 7, 129. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Tang, Y.; Zhang, X.; Chu, Z.; Liu, Y.; Tang, C. MMP-1 promotes osteogenic differentiation of human bone marrow mesen-chymal stem cells via the JNK and ERK pathway. Int. J. Biochem. Cell Biol. 2020, 129, 105880. [Google Scholar] [CrossRef]
- Ozeki, N.; Mogi, M.; Hase, N.; Hiyama, T.; Yamaguchi, H.; Kawai, R.; Nakata, K. Polyphosphate-induced matrix metalloproteinase-13 is required for osteoblast-like cell dif-ferentiation in human adipose tissue derived mesenchymal stem cells. Biosci. Trends 2016, 10, 365–371. [Google Scholar] [CrossRef]
- Ho, I.A.W.; Chan, K.Y.W.; Ng, W.-H.; Guo, C.M.; Hui, K.M.; Cheang, P.; Lam, P.Y.P. Matrix Metalloproteinase 1 Is Necessary for the Migration of Human Bone Marrow-Derived Mesenchymal Stem Cells Toward Human Glioma. Stem Cells 2009, 27, 1366–1375. [Google Scholar] [CrossRef]
- Lozito, T.P.; Tuan, R.S. Endothelial and cancer cells interact with mesenchymal stem cells via both microparticles and secreted factors. J. Cell. Mol. Med. 2014, 18, 2372–2384. [Google Scholar] [CrossRef]
- Nandi, P.; Lim, H.; Torres-Garcia, E.J.; Lala, P.K. Human trophoblast stem cell self-renewal and differentiation: Role of decorin. Sci. Rep. 2018, 8, 8977. [Google Scholar] [CrossRef]
- Simon, B.; Bucher, M.; Maloyan, A. A Primary Human Trophoblast Model to Study the Effect of Inflammation Associated with Maternal Obesity on Regulation of Autophagy in the Placenta. J. Vis. Exp. 2017, 127, 56484. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Go, Y.-Y.; Lee, C.-M.; Chae, S.-W.; Song, J.-J. Osteogenic Efficacy of Human Trophoblasts-Derived Conditioned Medium on Mesenchymal Stem Cells. Int. J. Mol. Sci. 2022, 23, 10196. https://doi.org/10.3390/ijms231710196
Go Y-Y, Lee C-M, Chae S-W, Song J-J. Osteogenic Efficacy of Human Trophoblasts-Derived Conditioned Medium on Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2022; 23(17):10196. https://doi.org/10.3390/ijms231710196
Chicago/Turabian StyleGo, Yoon-Young, Chan-Mi Lee, Sung-Won Chae, and Jae-Jun Song. 2022. "Osteogenic Efficacy of Human Trophoblasts-Derived Conditioned Medium on Mesenchymal Stem Cells" International Journal of Molecular Sciences 23, no. 17: 10196. https://doi.org/10.3390/ijms231710196