
The MKK2a Gene Involved in the MAPK Signaling Cascades Enhances Populus Salt Tolerance

Abstract
:1. Introduction
2. Results
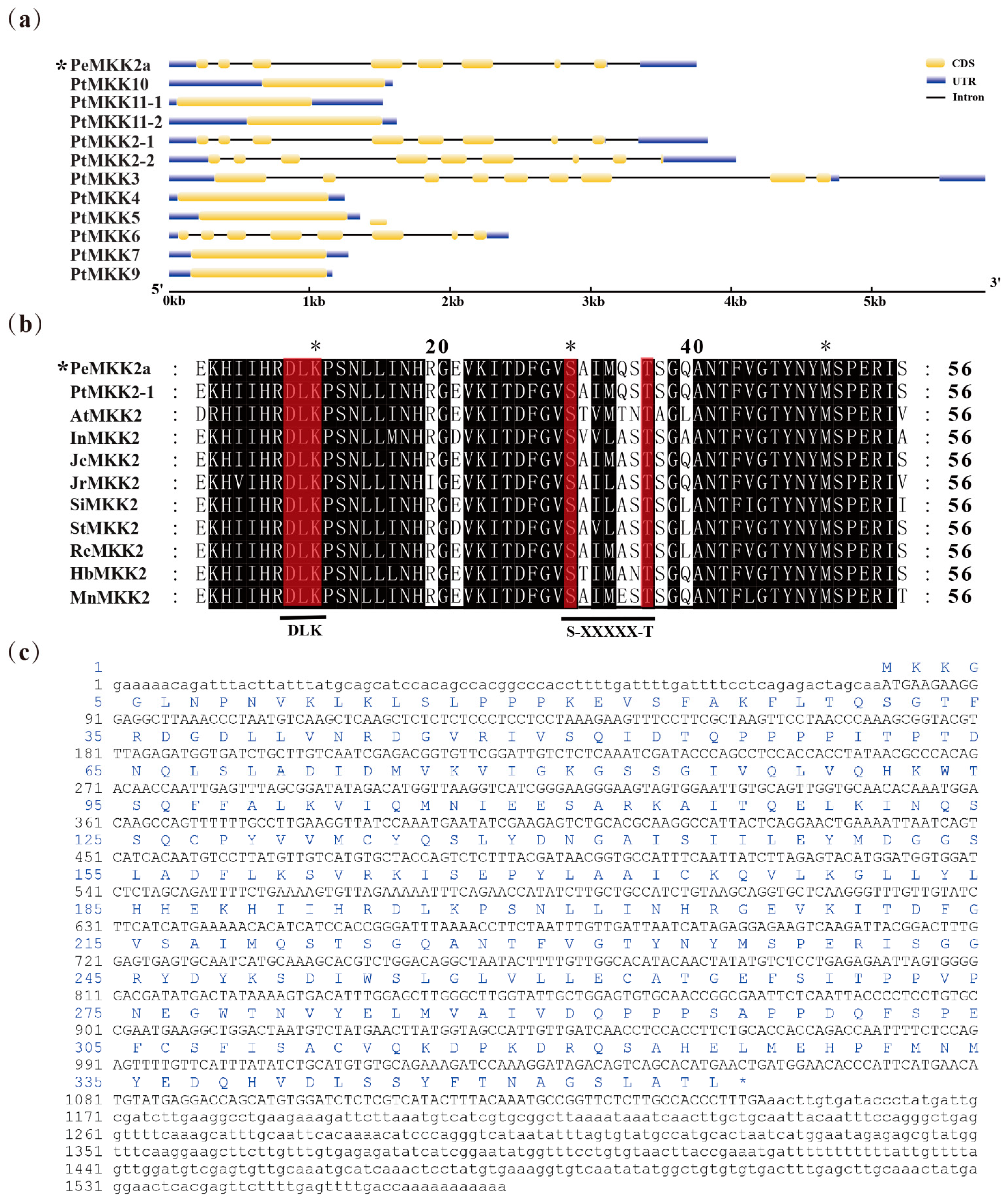
2.1. Cloning and Sequence Analysis of the PeMKK2a
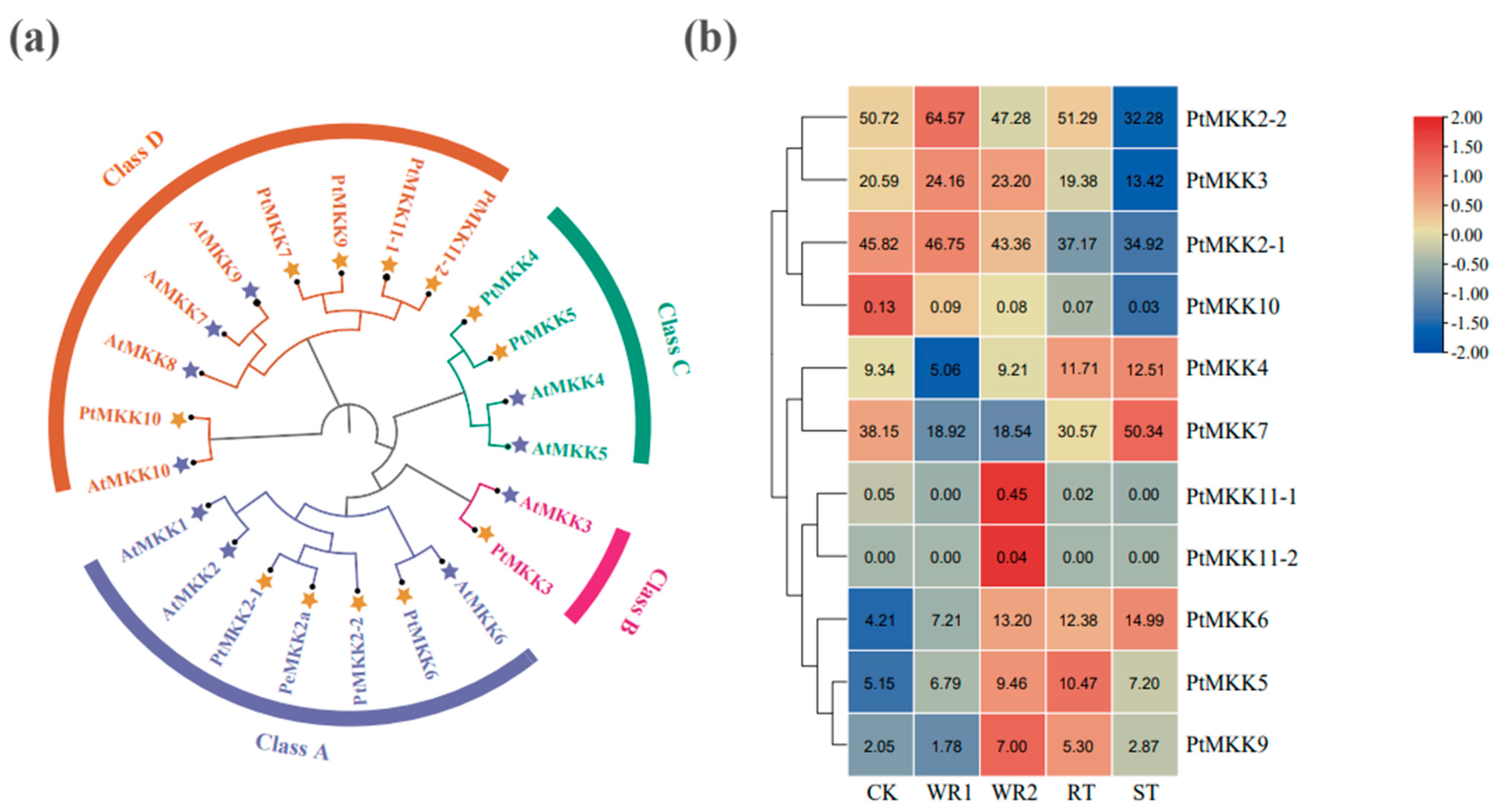
2.2. Expression Patterns of PtMKK Genes in Different Tissues
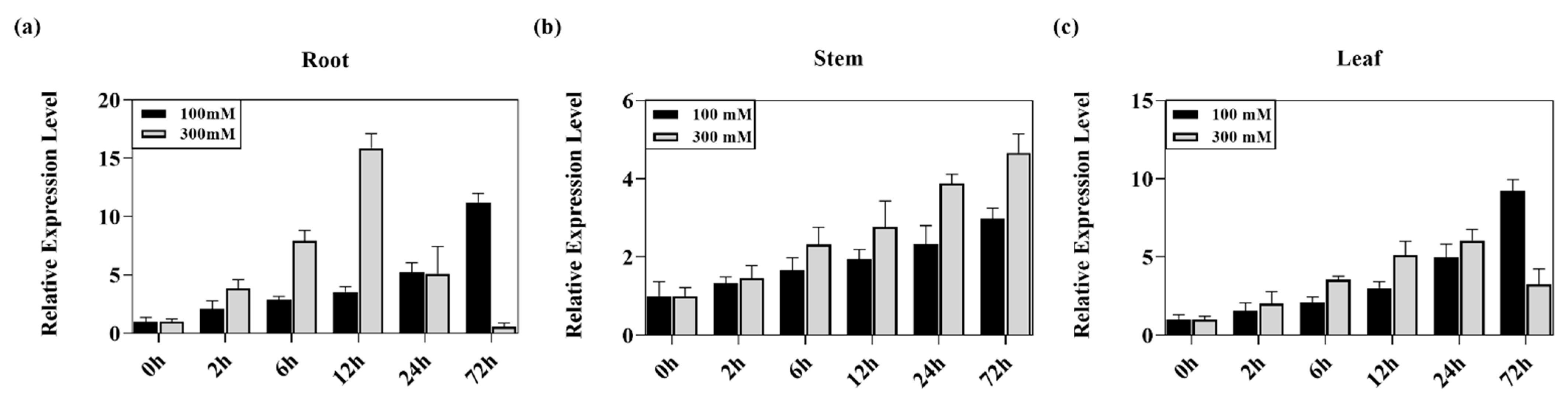
2.3. Expression Analysis of the PeMKK2a Gene in Response to Salt Stress
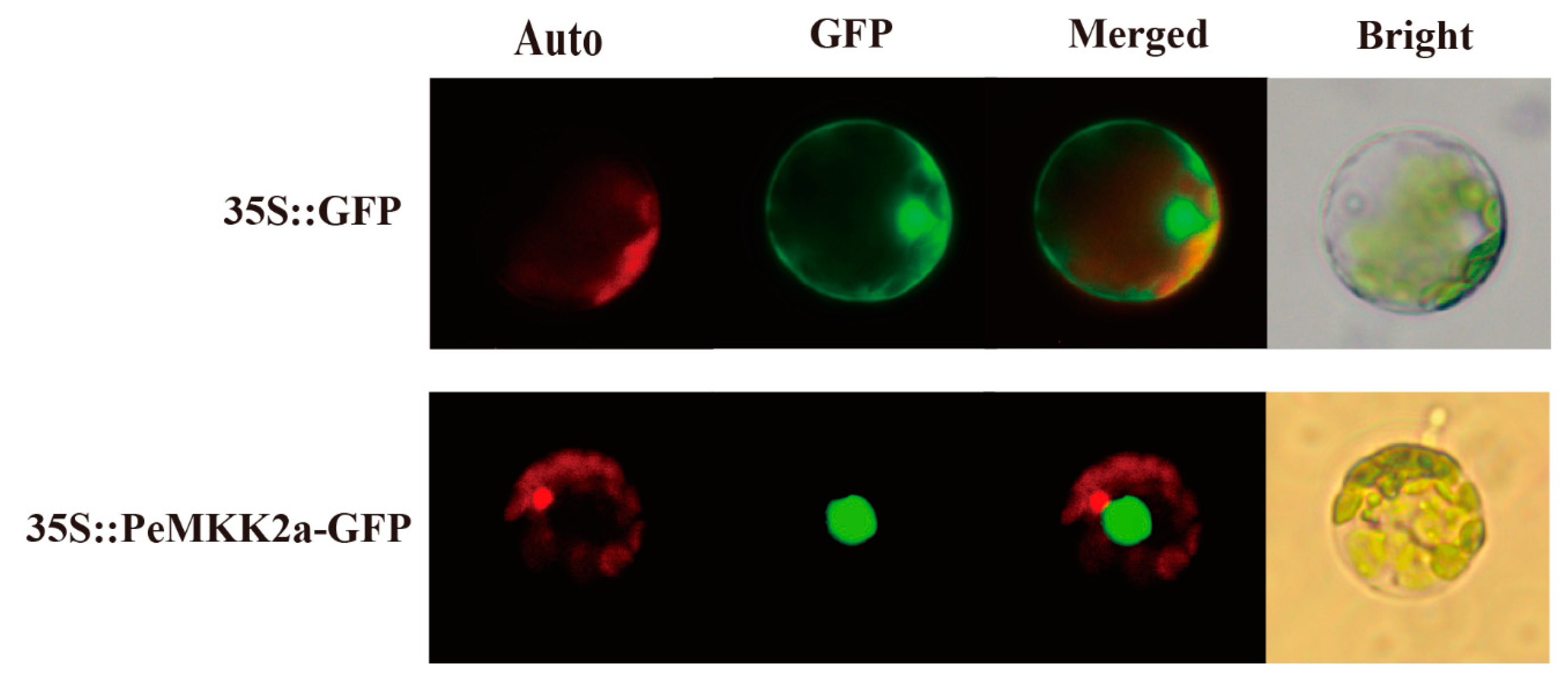
2.4. Subcellular Localization of the PeMKK2a Protein
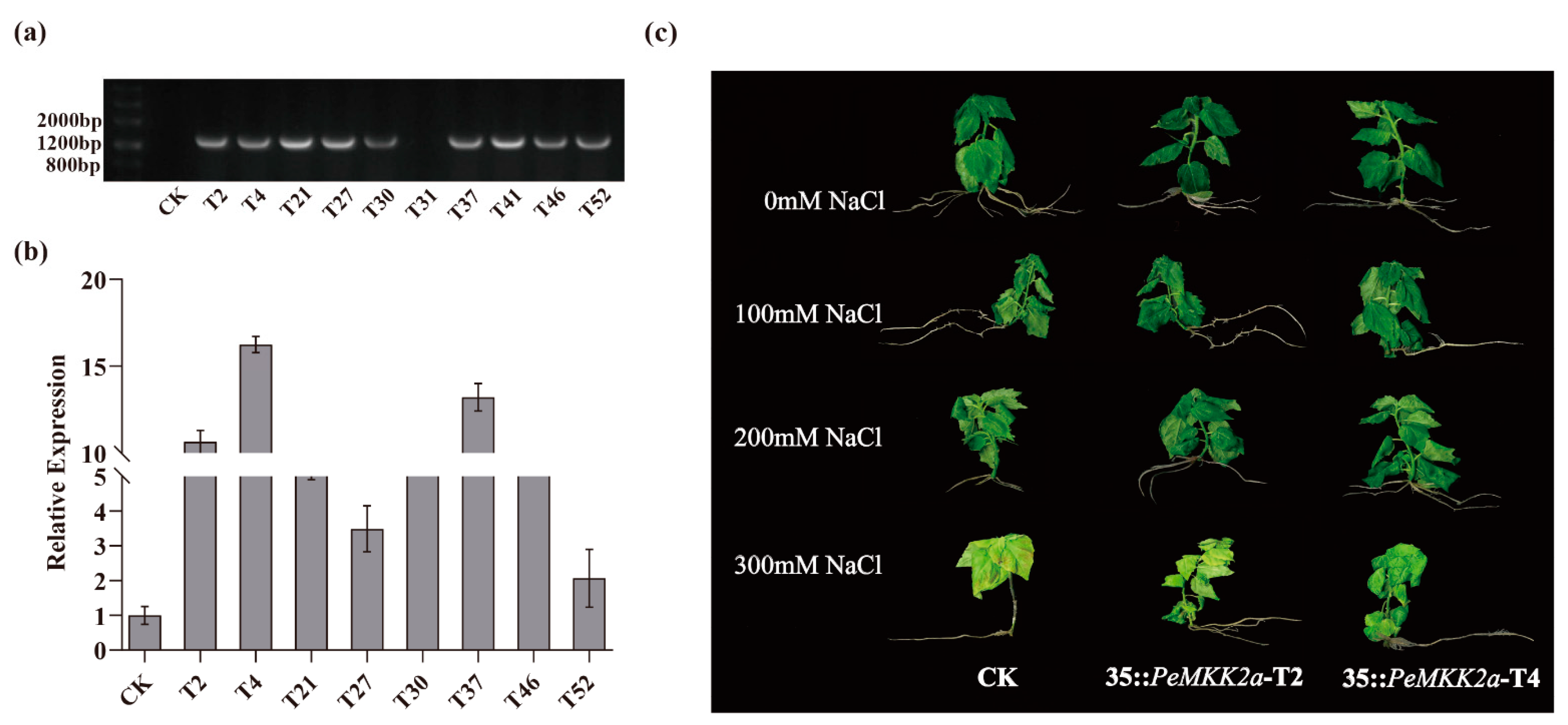
2.5. Generation of PeMKK2a-Overexpressing Transgenic Lines
2.6. Overexpression of PeMKK2a Enhances Salt Tolerance of Poplar
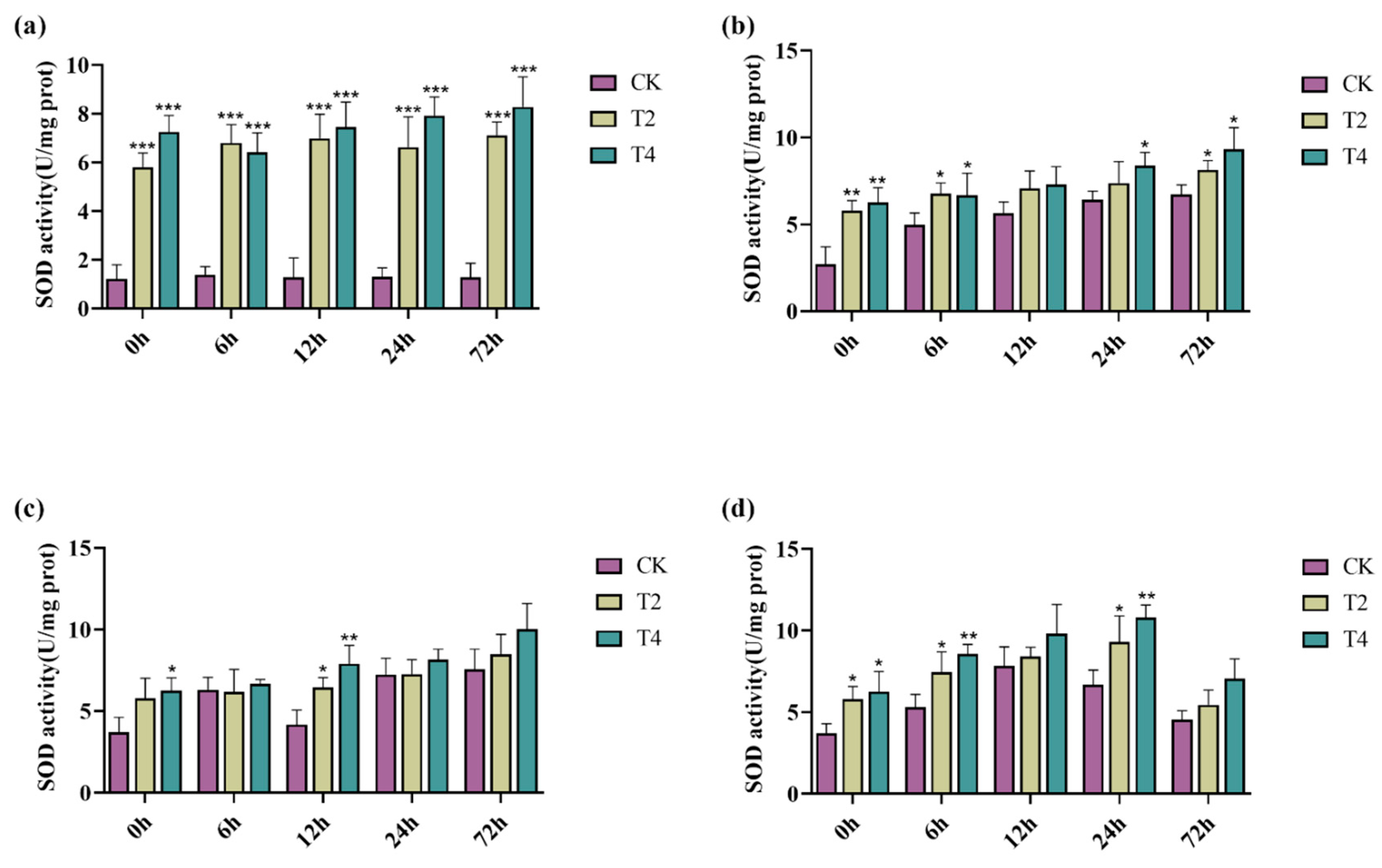
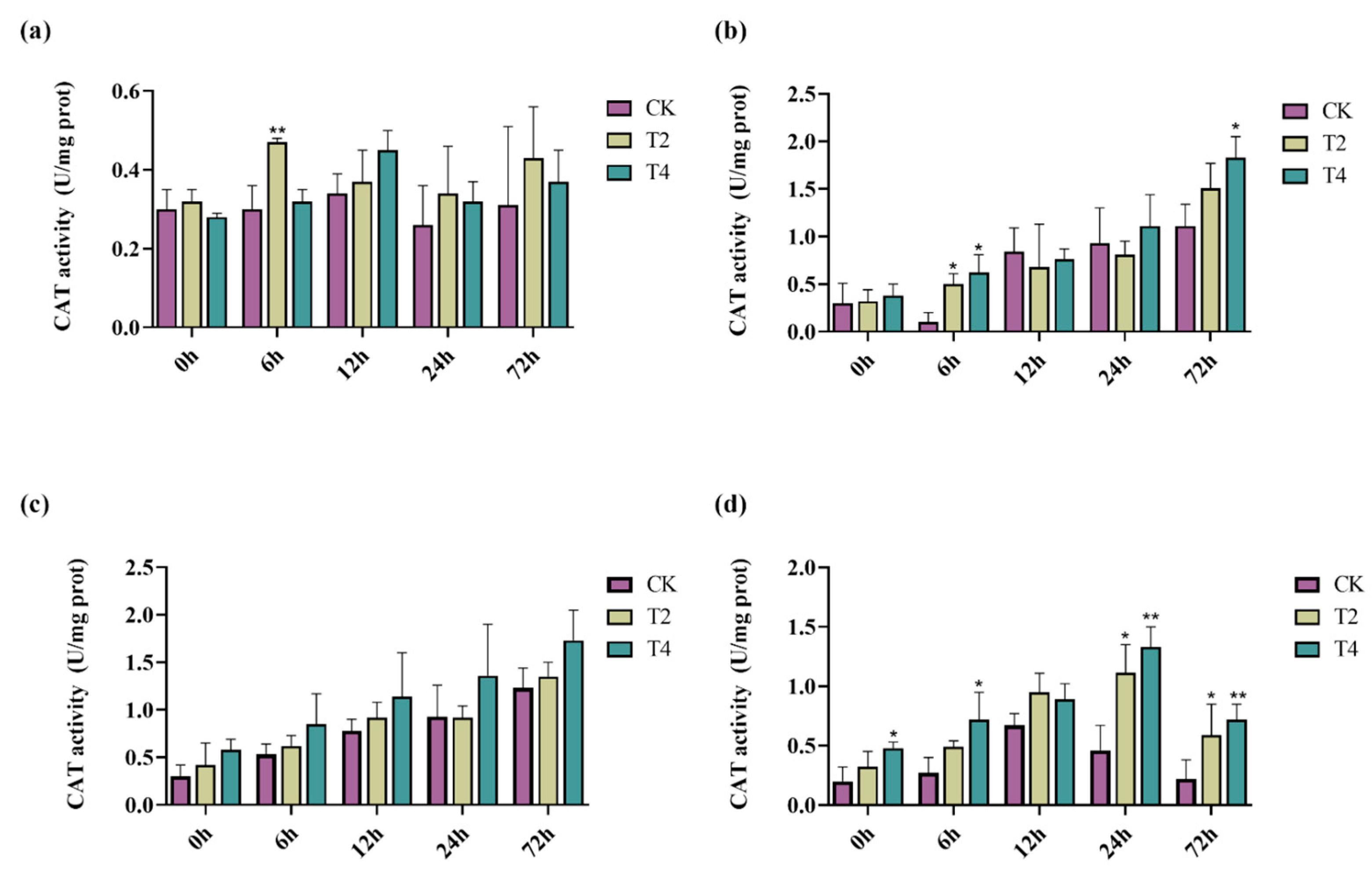
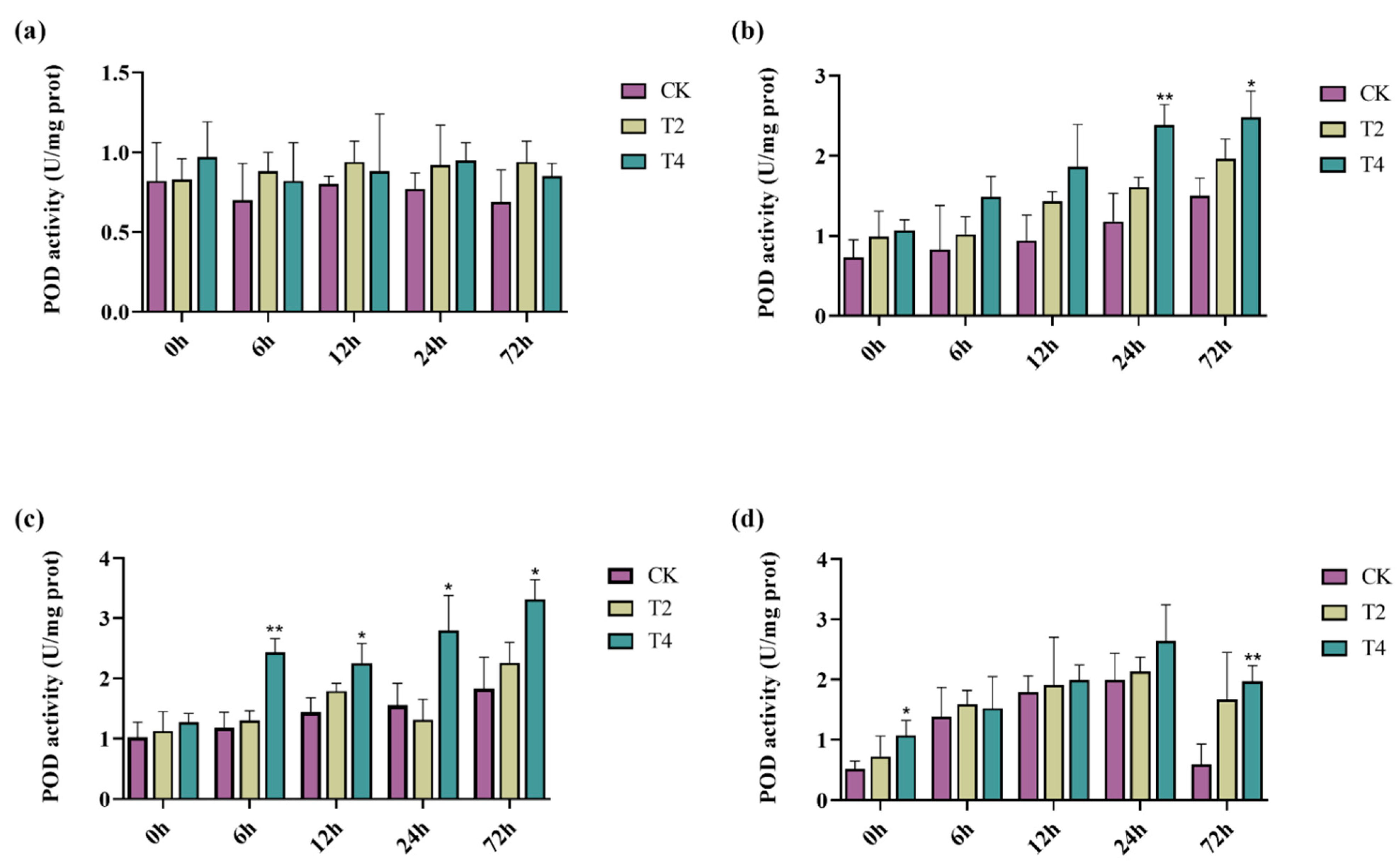
2.7. Overexpression of PeMKK2a Enhanced Antioxidant Capacity of Poplar
3. Discussion
4. Materials and Methods
4.1. Plant Materials, Growth Conditions, and Stress Treatments
4.2. Extraction of DNA and RNA, and cDNA Synthesis
4.3. Identification and Cloning of PeMKK2a Genes
4.4. Sequence Analysis
4.5. Expression Analysis of the PeMKK2a Gene of Poplar under Salt Stress
4.6. Transient Expression Vector Construction and Transformation
4.7. Overexpression Vector Construction and Transformation
4.8. Transgenic Poplar Confirmation and Salt Stress Treatment
4.9. Physiological Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Park, H.J.; Kim, W.-Y.; Yun, D.-J. A New Insight of Salt Stress Signaling in Plant. Mol. Cells 2016, 39, 447–459. [Google Scholar] [PubMed]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of Plant Responses to Salt Stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Karahara, I.; Katsuhara, M. Salinity tolerance mechanisms in glycophytes: An overview with the central focus on rice plants. Rice 2012, 5, 11. [Google Scholar] [PubMed]
- Colcombet, J.; Hirt, H. Arabidopsis MAPKs: A complex signalling network involved in multiple biological processes. Biochem. J. 2008, 413, 217–226. [Google Scholar] [PubMed]
- Sinha, A.K.; Jaggi, M.; Raghuram, B.; Tuteja, N. Mitogen-activated protein kinase signaling in plants under abiotic stress. Plant Signal Behav. 2011, 6, 196–203. [Google Scholar] [CrossRef]
- Tena, G.; Boudsocq, M.; Sheen, J. Protein kinase signaling networks in plant innate immunity. Curr. Opin. Plant Biol. 2011, 14, 519–529. [Google Scholar] [CrossRef]
- Liu, Y.F.; Zhang, F.; Liang, W.H. Function and mechanism of action of the MAPK cascade in rice. Chin. J. Biochem. Mol. Biol. 2021, 37, 1569–1576. [Google Scholar]
- Yin, B.B.; Pan, L.Y.; Fu, C. Regulation of MAPK cascade pathway gene expression in plants under adversity. Mol. Plant Breed. 2022, 20, 3257–3265. [Google Scholar]
- Seger, R.; Wexler, S. The MAPK Signaling Cascades. Encycl. Cell Biol. 2016, 3, 122–127. [Google Scholar]
- De Zelicourt, A.; Colcombet, J.; Hirt, H. The Role of MAPK Modules and ABA during Abiotic Stress Signaling. Trends Plant Sci. 2016, 21, 677–685. [Google Scholar]
- Cristina Rodriguez, M.; Petersen, M.; Mundy, J. Mitogen-Activated Protein Kinase Signaling in Plants. Annu. Rev. Plant Biol. 2010, 61, 621–649. [Google Scholar] [CrossRef] [PubMed]
- Keyse, S.M. Protein phosphatases and the regulation of mitogen-activated protein kinase signalling. Curr. Opin. Cell Biol. 2000, 12, 186–192. [Google Scholar] [CrossRef]
- Jonak, C.; Ökrész, L.; Bögre, L.; Hirt, H. Complexity, Cross Talk and Integration of Plant MAP Kinase Signalling. Curr. Opin. Plant Biol. 2002, 5, 415–424. [Google Scholar] [CrossRef]
- Takahashi, Y.; Soyano, T.; Kosetsu, K.; Sasabe, M.; Machida, Y. HINKEL kinesin, ANP MAPKKKs and MKK6/ANQ MAPKK, which phosphorylates and activates MPK4 MAPK, constitute a pathway that is required for cytokinesis in Arabidopsis thaliana. Plant Cell Physiol. 2010, 51, 1766–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, F.Y.; Hu, F.; Zhang, S.Y.; Wang, K.; Zhang, C.R.; Liu, T. MAPKs regulate root growth by influencing auxin signaling and cell cycle-related gene expression in cadmium-stressed rice. Environ. Sci. Pollut. Res. Int. 2013, 20, 5449–5460. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Su, J.; Zhang, Y.; Xu, J.; Zhang, S. Conveying endogenous and exogenous signals: MAPK cascades in plant growth and defense. Curr. Opin. Plant Biol. 2018, 45, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhang, S. Mitogen-activated protein kinase cascades in signaling plant growth and development. Trends Plant Sci. 2015, 20, 56–64. [Google Scholar] [CrossRef]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar]
- Asai, T.; Tena, G.; Plotnikova, J.; Willmann, M.R.; Chiu, W.-L.; Gomez-Gomez, L.; Boller, T.; Ausubel, F.M.; Sheen, J. MAP kinase signalling cascade in Arabidopsis innate immunity. Nature 2002, 415, 977–983. [Google Scholar] [CrossRef]
- Zhang, J.; Shao, F.; Li, Y.; Cui, H.; Chen, L.; Li, H.; Zou, Y.; Long, C.; Lan, L.; Chai, J.; et al. A Pseudomonas syringae Effector Inactivates MAPKs to Suppress PAMP-Induced Immunity in Plants. Cell Host Microbe 2007, 1, 175–185. [Google Scholar]
- Genot, B.; Lang, J.; Berriri, S.; Garmier, M.; Gilard, F.; Pateyron, S.; Haustraete, K.; Van Der Straeten, D.; Hirt, H.; Colcombet, J. Constitutively Active Arabidopsis MAP Kinase 3 Triggers Defense Responses Involving Salicylic Acid and SUMM2 Resistance Protein. Plant Physiol. 2017, 174, 1238–1249. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Zhang, S. MAPK cascades in plant disease resistance signaling. Annu. Rev. Phytopathol. 2013, 51, 245–266. [Google Scholar] [PubMed]
- Zhu, J.-K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Group, M.; Ichimura, K.; Shinozaki, K.; Tena, G.; Sheen, J.; Henry, Y.; Champion, A.; Kreis, M.; Zhang, S.; Hirt, H.; et al. Mitogen-activated protein kinase cascades in plants: A new nomenclature. Trends Plant Sci. 2002, 7, 301–308. [Google Scholar]
- Nicole, M.-C.; Hamel, L.-P.; Morency, M.-J.; Beaudoin, N.; Ellis, B.E.; Séguin, A. MAP-ping genomic organization and organ-specific expression profiles of poplar MAP kinases and MAP kinase kinases. BMC Genom. 2006, 7, 223. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, D.; Wang, L.; Li, D. Genome-Wide Analysis of Mitogen-Activated Protein Kinase Gene Family in Maize. Plant Mol. Biol. Rep. 2013, 31, 1446–1460. [Google Scholar] [CrossRef]
- Singh, R.; Lee, J.-E.; Dangol, S.; Choi, J.; Yoo, R.H.; Moon, J.S.; Shim, J.-K.; Rakwal, R.; Agrawal, G.K.; Jwa, N.-S. Protein interactome analysis of 12 mitogen-activated protein kinase kinase kinase in rice using a yeast two-hybrid system. Proteomics 2014, 14, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Alzwiy, I.A.; Morris, P.C. A mutation in the Arabidopsis MAP kinase kinase 9 gene results in enhanced seedling stress tolerance. Plant Sci. 2007, 173, 302–308. [Google Scholar] [CrossRef]
- Taj, G.; Agarwal, P.; Grant, M.; Kumar, A. MAPK machinery in plants: Recognition and response to different stresses through multiple signal transduction pathways. Plant Signal Behav. 2010, 5, 1370–1378. [Google Scholar] [CrossRef]
- Blanc, G.; Wolfe, K.H. Functional divergence of duplicated genes formed by polyploidy during Arabidopsis evolution. Plant Cell 2004, 16, 1679–1691. [Google Scholar]
- Mészáros, T.; Helfer, A.; Hatzimasoura, E.; Magyar, Z.; Serazetdinova, L.; Rios, G.; Bardóczy, V.; Teige, M.; Koncz, C.; Peck, S.; et al. The Arabidopsis MAP kinase kinase MKK1 participates in defence responses to the bacterial elicitor flagellin. Plant J. 2006, 48, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Jing, W.; Zhang, W. The mitogen-activated protein kinase cascade MKK1-MPK4 mediates salt signaling in rice. Plant Sci. 2014, 227, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Teige, M.; Scheikl, E.; Eulgem, T.; Dóczi, R.; Ichimura, K.; Shinozaki, K.; Dangl, J.L.; Hirt, H. The MKK2 Pathway Mediates Cold and Salt Stress Signaling in Arabidopsis. Mol. Cell 2004, 15, 141–152. [Google Scholar] [CrossRef]
- Pedley, K.F.; Martin, G.B. Identification of MAPKs and Their Possible MAPK Kinase Activators Involved in the Pto-mediated Defense Response of Tomato*. J. Biol. Chem. 2004, 279, 49229–49235. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.-L.; Zhou, L.; Yun, B.-W.; Nielsen, H.B.; Fiil, B.K.; Petersen, K.; MacKinlay, J.; Loake, G.J.; Mundy, J.; Morris, P.C. Arabidopsis Mitogen-Activated Protein Kinase Kinases MKK1 and MKK2 Have Overlapping Functions in Defense Signaling Mediated by MEKK1, MPK4, and MKS1. Plant Physiol. 2008, 148, 212–222. [Google Scholar] [CrossRef]
- Huang, Y.; Li, H.; Gupta, R.; Morris, P.C.; Luan, S.; Kieber, J.J. ATMPK4, an Arabidopsis homolog of mitogen-activated protein kinase, is activated in vitro by AtMEK1 through threonine phosphorylation. Plant Physiol. 2000, 122, 1301–1310. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Liu, J.; Bi, D.; Zhang, Z.; Cheng, F.; Chen, S.; Zhang, Y. MEKK1, MKK1/MKK2 and MPK4 function together in a mitogen-activated protein kinase cascade to regulate innate immunity in plants. Cell Res. 2008, 18, 1190–1198. [Google Scholar] [CrossRef]
- Kosetsu, K.; Matsunaga, S.; Nakagami, H.; Colcombet, J.; Sasabe, M.; Soyano, T.; Takahashi, Y.; Hirt, H.; Machida, Y. The MAP kinase MPK4 is required for cytokinesis in Arabidopsis thaliana. Plant Cell 2010, 22, 3778–3790. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, F.; Yoshida, R.; Ichimura, K.; Mizoguchi, T.; Seo, S.; Yonezawa, M.; Maruyama, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. The Mitogen-Activated Protein Kinase Cascade MKK3–MPK6 Is an Important Part of the Jasmonate Signal Transduction Pathway in Arabidopsis. Plant Cell 2007, 19, 805–818. [Google Scholar]
- Sethi, V.; Raghuram, B.; Sinha, A.K.; Chattopadhyay, S. A mitogen-activated protein kinase cascade module, MKK3-MPK6 and MYC2, is involved in blue light-mediated seedling development in Arabidopsis. Plant Cell 2014, 26, 3343–3357. [Google Scholar] [CrossRef]
- Jalmi, S.K.; Sinha, A.K. Functional Involvement of a Mitogen Activated Protein Kinase Module, OsMKK3-OsMPK7-OsWRK30 in Mediating Resistance against Xanthomonas oryzae in Rice. Sci. Rep. 2016, 6, 37974. [Google Scholar] [PubMed]
- Mao, X.; Zhang, J.; Liu, W.; Yan, S.; Liu, Q.; Fu, H.; Zhao, J.; Huang, W.; Dong, J.; Zhang, S.; et al. The MKKK62-MKK3-MAPK7/14 module negatively regulates seed dormancy in rice. Rice 2019, 12, 2. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Henry, J.; Auger, T.; Vogt, J.B.; Almazouzi, A.; Glasbrenner, H.; Groeschel, F. Assessment of the lifetime of the beam window of MEGAPIE target liquid metal container. J. Nucl. Mater. 2006, 356, 308–320. [Google Scholar] [CrossRef]
- Lei, L.; Li, Y.; Wang, Q.; Xu, J.; Chen, Y.; Yang, H.; Ren, D. Activation of MKK9-MPK3/MPK6 enhances phosphate acquisition in Arabidopsis thaliana. New Phytol. 2014, 203, 1146–1160. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.; Chen, W.; Wang, B.; Zhu, F.; Li, Y.; Yang, H.; Li, J.; Ren, D. Arabidopsis MKK10-MPK6 mediates red-light-regulated opening of seedling cotyledons through phosphorylation of PIF3. J. Exp. Bot. 2018, 69, 423–439. [Google Scholar] [CrossRef]
- Chang, Y.; Yang, H.; Ren, D.; Li, Y. Activation of ZmMKK10, a maize mitogen-activated protein kinase kinase, induces ethylene-dependent cell death. Plant Sci. 2017, 264, 129–137. [Google Scholar] [CrossRef]
- Ma, H.; Chen, J.; Zhang, Z.; Ma, L.; Yang, Z.; Zhang, Q.; Li, X.; Xiao, J.; Wang, S. MAPK kinase 10.2 promotes disease resistance and drought tolerance by activating different MAPKs in rice. Plant J. 2017, 92, 557–570. [Google Scholar] [CrossRef]
- Wu, J.; Wang, J.; Pan, C.; Guan, X.; Wang, Y.; Liu, S.; He, Y.; Chen, J.; Chen, L.; Lu, G. Genome-wide identification of MAPKK and MAPKKK gene families in tomato and transcriptional profiling analysis during development and stress response. PLoS ONE 2014, 9, e103032. [Google Scholar] [CrossRef]
- Kong, X.; Pan, J.; Zhang, D.; Jiang, S.; Cai, G.; Wang, L.; Li, D. Identification of mitogen-activated protein kinase kinase gene family and MKK–MAPK interaction network in maize. Biochem. Biophys. Res. Commun. 2013, 441, 964–969. [Google Scholar]
- Wang, C. The Bioological Function and Regulatory Mechanism of GhMKK6 Mediated MAPK Cascades Signaling Pathway in Cotton. Ph.D. Thesis, Shandong Agricultural University, Tai’an, China, 2018. [Google Scholar]
- Cai, H.; Yang, C.; Liu, S.; Qi, H.; Wu, L.; Xu, L.A.; Xu, M. MiRNA-target pairs regulate adventitious rooting in Populus: A functional role for miR167a and its target Auxin response factor 8. Tree Physiol. 2019, 39, 1922–1936. [Google Scholar] [CrossRef]
- Liu, S.; Wu, L.; Qi, H.; Xu, M. LncRNA/circRNA–miRNA–mRNA networks regulate the development of root and shoot meristems of Populus. Ind. Crops Prod. 2019, 133, 333–347. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, S.; He, W.-D.; Shao, X.-H.; Li, C.-Y.; Wei, Y.-R.; Deng, G.-M.; Kuang, R.-B.; Hu, C.-H.; Yi, G.-J.; et al. Comparative Phosphoproteomics Reveals an Important Role of MKK2 in Banana (Musa spp.) Cold Signal Network. Sci. Rep. 2017, 7, 40852. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Ma, S.M.; Nie, Y.B. Cloning and functional analysis of the TaMKK2 gene in wheat. Mol. Plant Breed. 2022, 2021, 1–13. [Google Scholar]
- Xu, M.; Chen, C.; Cai, H.; Wu, L. Overexpression of PeHKT1;1 Improves Salt Tolerance in Populus. Genes 2018, 9, 475. [Google Scholar] [CrossRef] [PubMed]
- Hamel, L.-P.; Nicole, M.-C.; Sritubtim, S.; Morency, M.-J.; Ellis, M.; Ehlting, J.; Beaudoin, N.; Barbazuk, B.; Klessig, D.; Lee, J.; et al. Ancient signals: Comparative genomics of plant MAPK and MAPKK gene families. Trends Plant Sci. 2006, 11, 192–198. [Google Scholar] [CrossRef]
- Nakagami, H.; Soukupová, H.; Schikora, A.; Zárský, V.; Hirt, H. A Mitogen-activated Protein Kinase Kinase Kinase Mediates Reactive Oxygen Species Homeostasis in Arabidopsis*. J. Biol. Chem. 2006, 281, 38697–38704. [Google Scholar] [CrossRef]
- Xu, M.; Zhang, B.; Su, X.; Zhang, S.; Huang, M. Reference gene selection for quantitative real-time polymerase chain reaction in Populus. Anal. Biochem. 2011, 408, 337–339. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Tan, B.; Xu, M.; Chen, Y.; Huang, M. Transient expression for functional gene analysis using Populus protoplasts. Plant Cell Tissue Organ Cult. 2013, 114, 11–18. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer_ID | Forward PCR Primer (5′-3′) | Reverse PCR Primer (5′-3′) |
|---|---|---|
| PeMKK2a_ORF | ATGAAGAAGGGAGGCTTAAACCCTAATG | TCAAAGGGTGGCAAGAGAACC |
| PeMKK2a_3OUTER | ATCGATACCCAGCCTCCACCACC | CGCGGATCCACAGCCTACTGATGATCAGTCGATG |
| PeMKK2a_3INNER | TGGTTAAGGTCATCGGGAAGGGAAG | TGGTTAAGGTCATCGGGAAGGGAAG |
| PeMKK2a_5OUTER PeMKK2a_5INNER | GCACTCACTCCAAAGTCCGTAA ACAGATGGCAGCAAGATATGGTT | CGCGGATCCACAGCCTACTGATGATCAGTCGATG TGGTTAAGGTCATCGGGAAGGGAAG |
| PeMKK2a_qRT-PCR | CCCTCCTGTGCCGAATGAAGG | TGGGTGTTCCATCAGTTCATGTGC |
| PeMPK4_qRT-PCR | ACTCACGGCGGCCAATTCAT | CGCCACCATCTCGTTCGTCT |
| PeMPK6_qRT-PCR | AGGAGGAGGAGGTGGAGGGA | CAACGCCGAACAGACGATGC |
| PeMKK2a_p35Sf3 | AGGAAGGTGGCTCCTACAAATGCCATC | TCAAAGGGTGGCAAGAGAACC |
| 18S [55] | TCAACTTTCGATGGTAGGATAGTG | CCGTGTCAGGATTGGGTAATTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Sun, Z.; Chen, C.; Xu, M. The MKK2a Gene Involved in the MAPK Signaling Cascades Enhances Populus Salt Tolerance. Int. J. Mol. Sci. 2022, 23, 10185. https://doi.org/10.3390/ijms231710185
Wang J, Sun Z, Chen C, Xu M. The MKK2a Gene Involved in the MAPK Signaling Cascades Enhances Populus Salt Tolerance. International Journal of Molecular Sciences. 2022; 23(17):10185. https://doi.org/10.3390/ijms231710185
Chicago/Turabian StyleWang, Jiali, Zimou Sun, Caihui Chen, and Meng Xu. 2022. "The MKK2a Gene Involved in the MAPK Signaling Cascades Enhances Populus Salt Tolerance" International Journal of Molecular Sciences 23, no. 17: 10185. https://doi.org/10.3390/ijms231710185