
Systemic Beta-Hydroxybutyrate Affects BDNF and Autophagy into the Retina of Diabetic Mice
 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
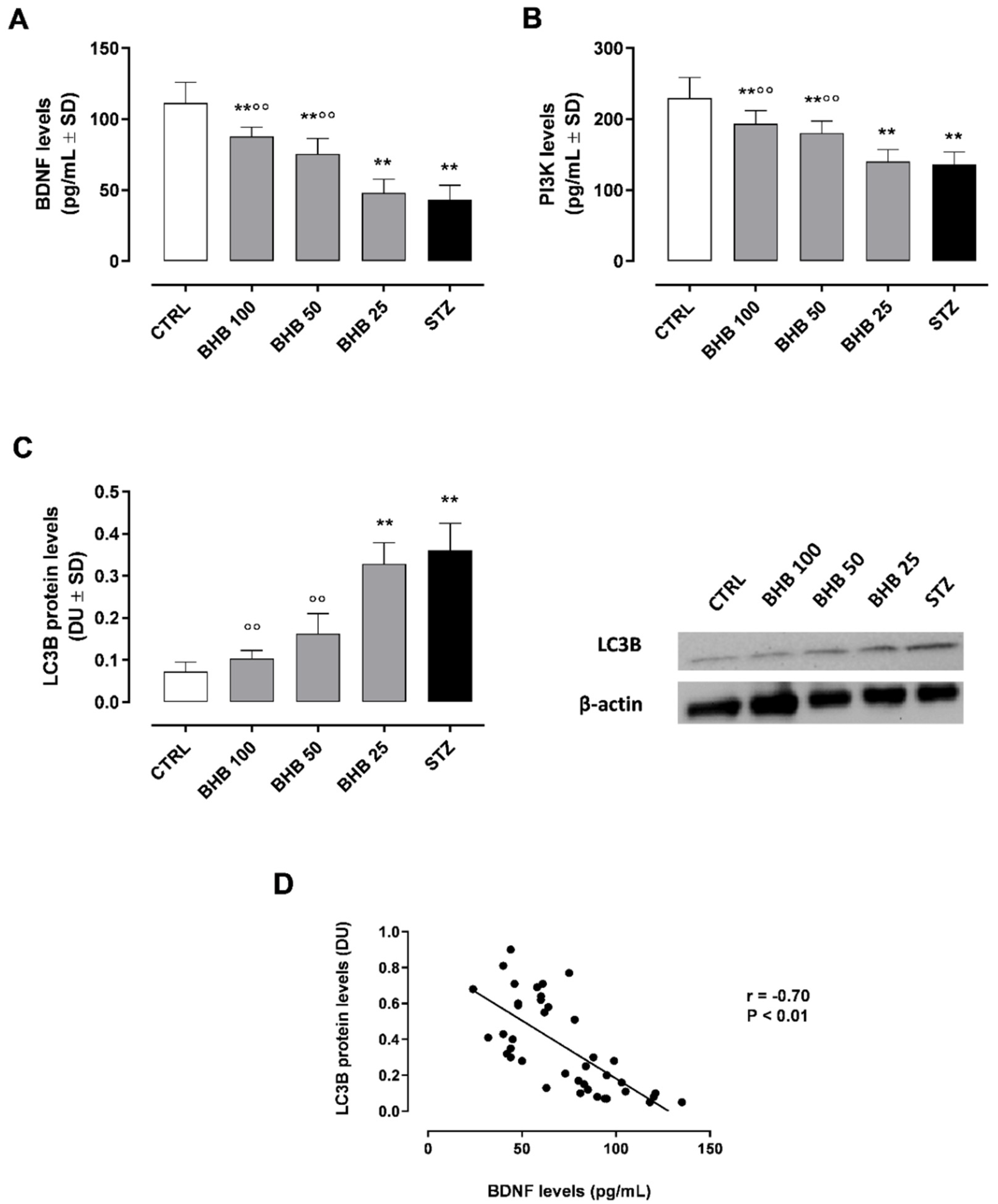
2.1. BHB Restores Retinal BDNF Levels and Activity
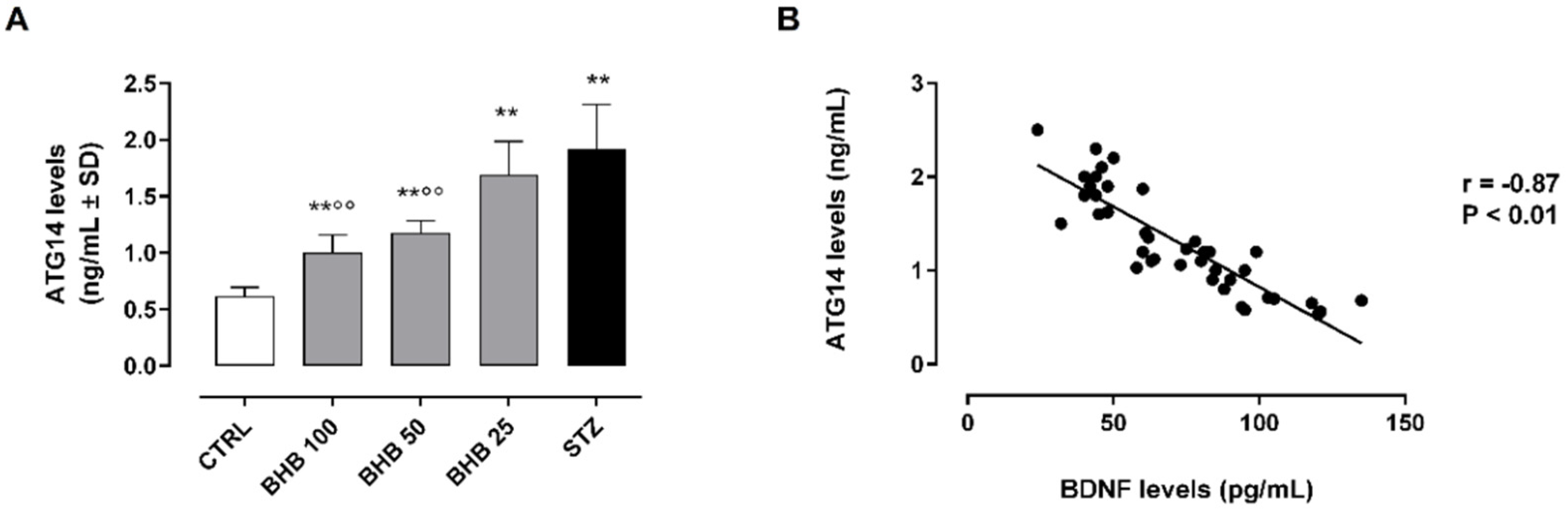
2.2. BHB Reduces the Diabetes-Induced Autophagy through BDNF
2.3. BDNF Upregulation by BHB Reduces the Autophagosome–Lysosome Formation in Diabetic Retina
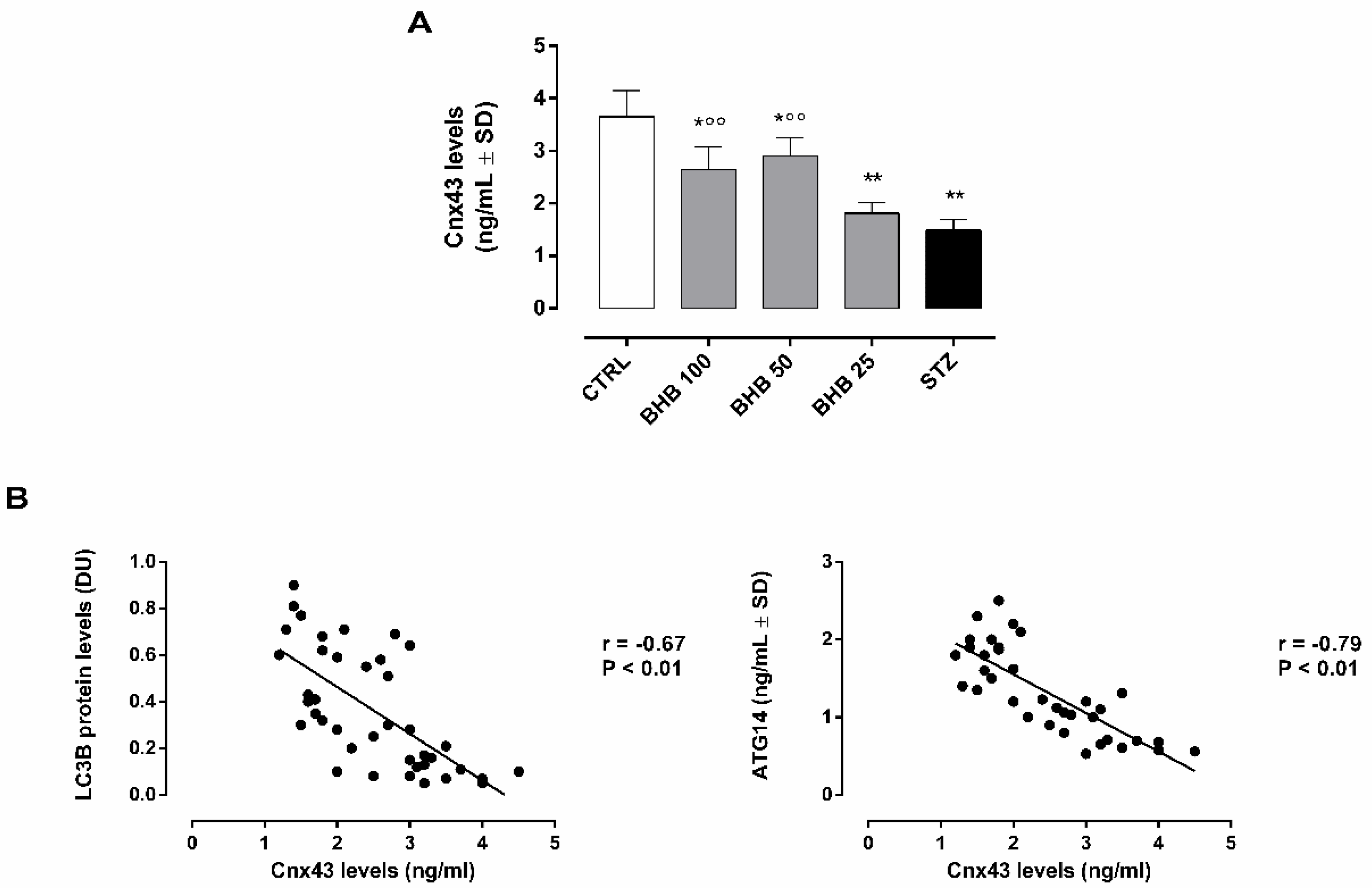
2.4. BHB Improves Retinal Gap Junctions in Diabetic Retina trough the Regulation of Autophagosome–Lysosome Formation
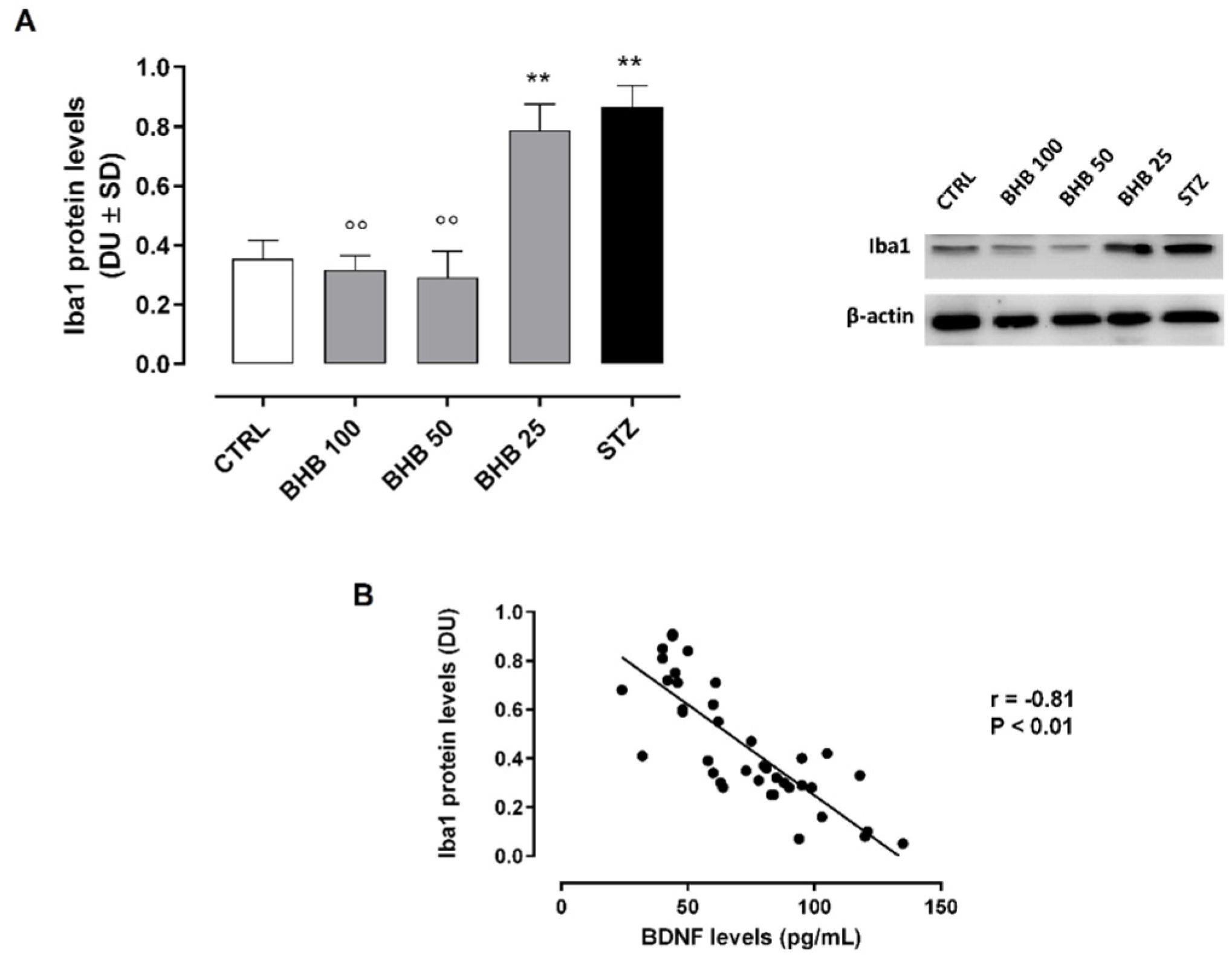
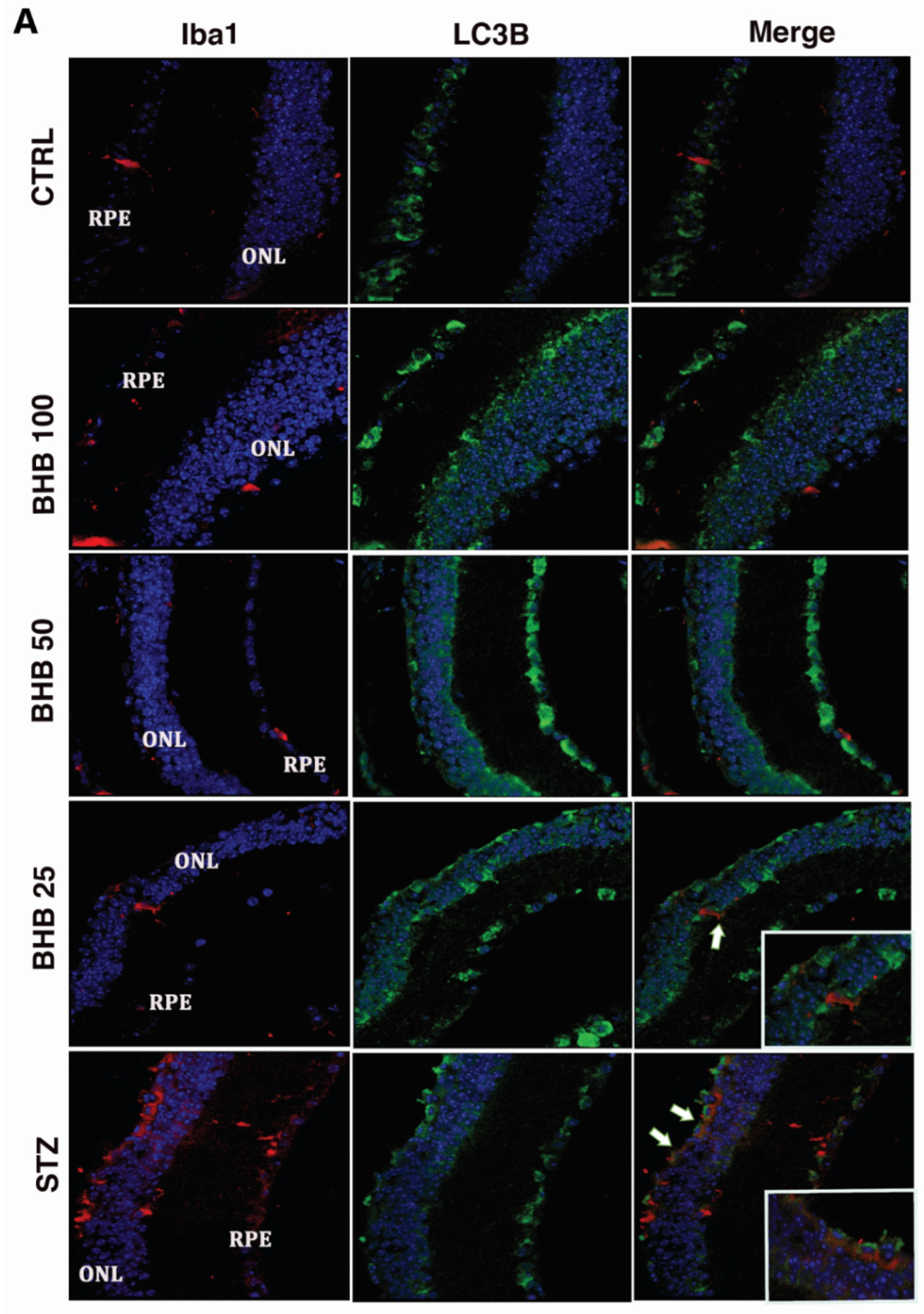
2.5. BHB Reduces M1 Microglial Activation in Diabetic Retina
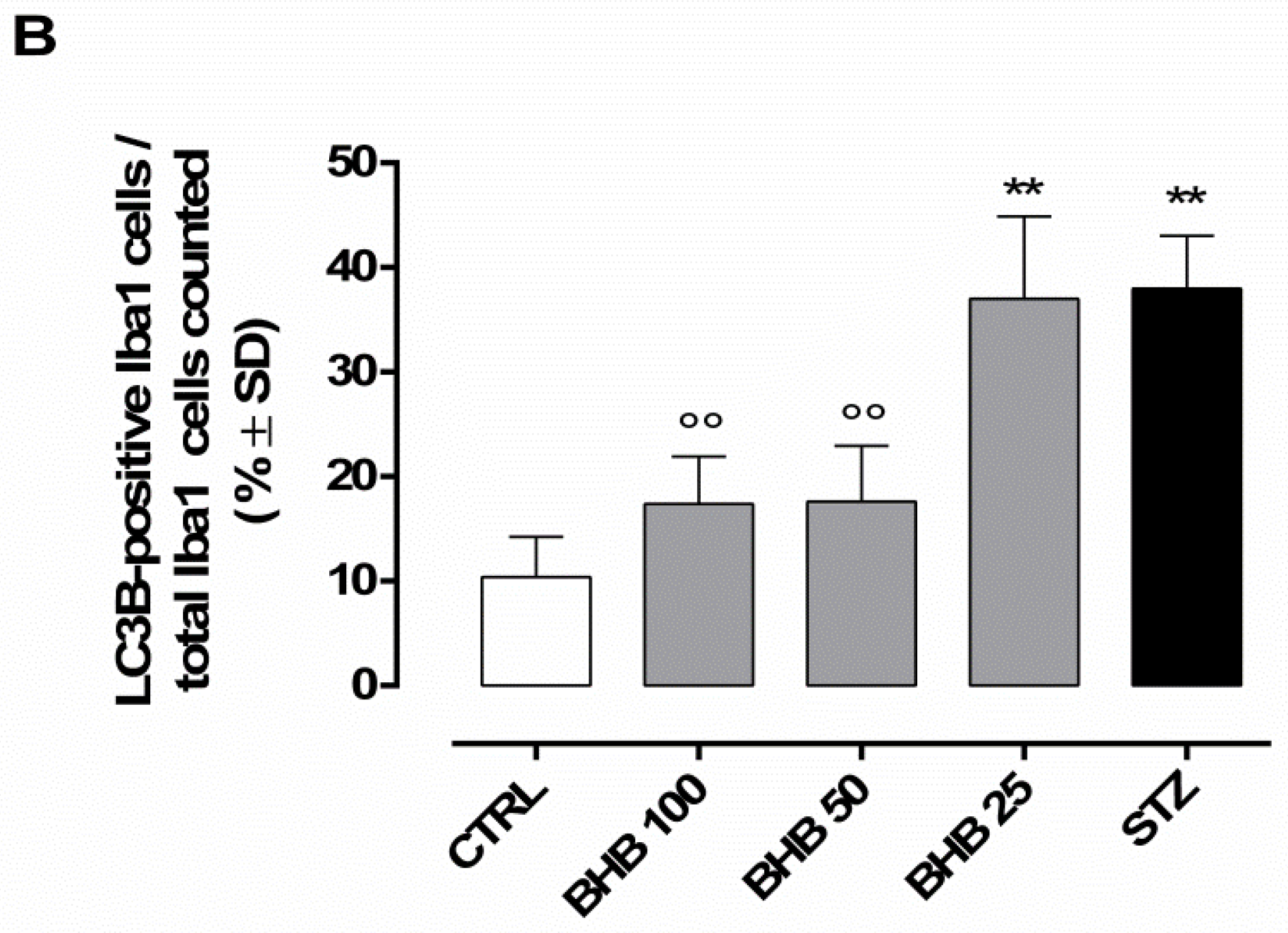
2.6. BHB Decreases Retinal Microglial Autophagy Dysregulated by Diabetes
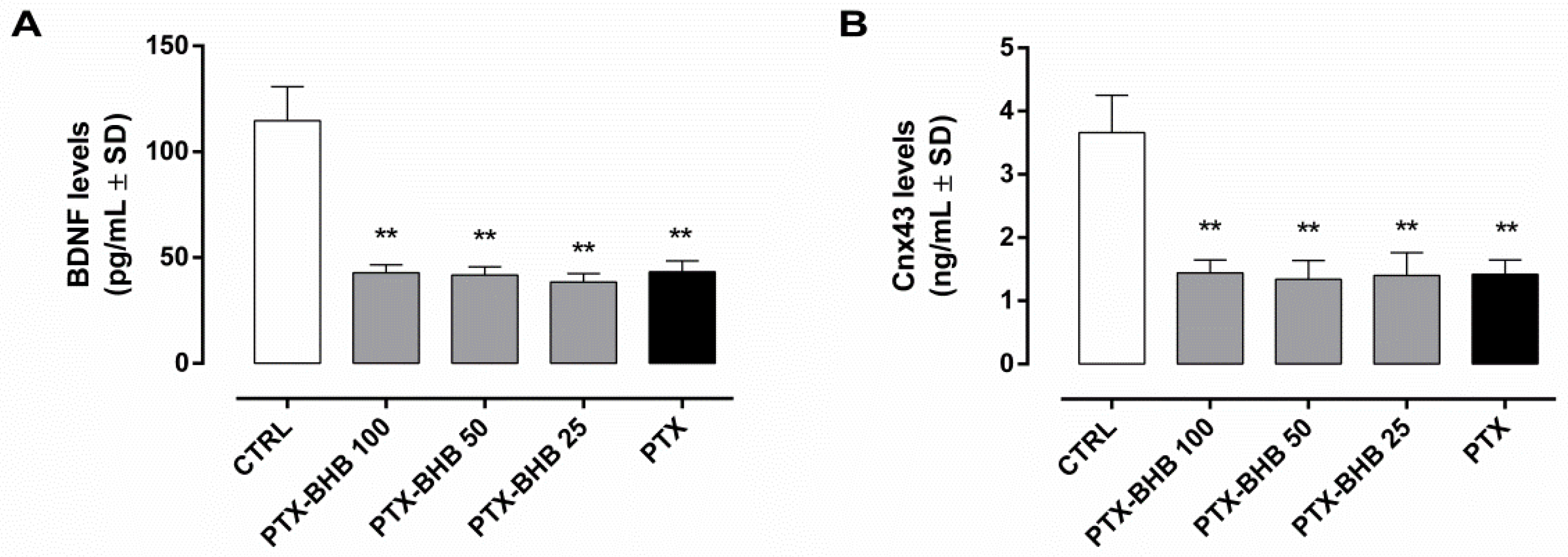
2.7. HCA2 Blocking by Pertussis Toxin (PTX) Abolishes BHB Effects on BDNF and Cnx43
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Procedures
4.2. Enzyme-Linked Immunosorbent Assay (ELISA)
4.3. Immunofluorescence
4.4. Protein Isolation and Quantization
4.5. Western Blotting
4.6. In Vivo Proof of Concept
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Altmann, C.; Schmidt, M.H.H. The Role of Microglia in Diabetic Retinopathy: Inflammation, Microvasculature Defects and Neurodegeneration. Int. J. Mol. Sci. 2018, 19, 110. [Google Scholar] [CrossRef] [PubMed]
- Afarid, M.; Namvar, E.; Sanie-Jahromi, F. Diabetic Retinopathy and BDNF: A Review on Its Molecular Basis and Clinical Applications. J. Ophthalmol. 2020, 2020, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Seki, M.; Tanaka, T.; Nawa, H.; Usui, T.; Fukuchi, T.; Ikeda, K.; Abe, H.; Takei, N. Involvement of Brain-Derived Neurotrophic Factor in Early Retinal Neuropathy of Streptozotocin-Induced Diabetes in Rats. Diabetes 2004, 53, 2412–2419. [Google Scholar] [CrossRef]
- Taşlipinar Uzel, A.G.; UĞurlu, N.; Toklu, Y.; Çİçek, M.; Boral, B.; Şener, B.; ÇaĞil, N. Relationship between stages of diabetic retinopathy and levels of brain-derived neurotrophic factor in aqueous humor and serum. Retina 2020, 40, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Adornetto, A.; Gesualdo, C.; Laganà, M.L.; Trotta, M.C.; Rossi, S.; Russo, R. Autophagy: A Novel Pharmacological Target in Diabetic Retinopathy. Front. Pharmacol. 2021, 12, 695267. [Google Scholar] [CrossRef]
- Yin, X.; He, T.; Chen, R.; Cui, H.; Li, G. Impact of Neurotrophic Factors Combination Therapy on Retinitis Pigmentosa. J. Int. Med. Res. 2020, 48, 030006052096783. [Google Scholar] [CrossRef]
- Shim, M.S.; Kim, K.-Y.; Noh, M.; Ko, J.Y.; Ahn, S.; An, M.A.; Iwata, T.; Perkins, G.A.; Weinreb, R.N.; Ju, W.-K. Optineurin E50K Triggers BDNF Deficiency-Mediated Mitochondrial Dysfunction in Retinal Photoreceptor Cell Line. Biochem. Biophys. Res. Commun. 2018, 503, 2690–2697. [Google Scholar] [CrossRef]
- Lichtenstein, A.; Minogue, P.J.; Beyer, E.C.; Berthoud, V.M. Autophagy: A Pathway That Contributes to Connexin Degradation. J. Cell Sci. 2011, 124, 910–920. [Google Scholar] [CrossRef]
- Trotta, M.C.; Maisto, R.; Guida, F.; Boccella, S.; Luongo, L.; Balta, C.; D’Amico, G.; Herman, H.; Hermenean, A.; Bucolo, C.; et al. The Activation of Retinal HCA2 Receptors by Systemic Beta-Hydroxybutyrate Inhibits Diabetic Retinal Damage through Reduction of Endoplasmic Reticulum Stress and the NLRP3 Inflammasome. PLoS ONE 2019, 14, e0211005. [Google Scholar] [CrossRef]
- Bobbie, M.W.; Roy, S.; Trudeau, K.; Munger, S.J.; Simon, A.M.; Roy, S. Reduced Connexin 43 Expression and Its Effect on the Development of Vascular Lesions in Retinas of Diabetic Mice. Investig. Ophthalmol. Vis. Sci. 2010, 51, 3758. [Google Scholar] [CrossRef]
- Tien, T.; Muto, T.; Zhang, J.; Sohn, E.H.; Mullins, R.F.; Roy, S. Association of Reduced Connexin 43 Expression with Retinal Vascular Lesions in Human Diabetic Retinopathy. Exp. Eye Res. 2016, 146, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Hu, E.; Du, H.; Zhu, X.; Wang, L.; Shang, S.; Wu, X.; Lu, H.; Lu, X. Beta-Hydroxybutyrate Promotes the Expression of BDNF in Hippocampal Neurons under Adequate Glucose Supply. Neuroscience 2018, 386, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Marosi, K.; Mattson, M. Ketone Beta-Hydroxybutyrate up-regulates BDNF expression through NF-ΚB as an adaptive response against ROS, which may improve neuronal bioenergetics and enhance neuroprotection (P3.090). Neurology 2017, 88 (Suppl. 16), P3.090. [Google Scholar]
- Hu, E.; Du, H.; Shang, S.; Zhang, Y.; Lu, X. Beta-Hydroxybutyrate Enhances BDNF Expression by Increasing H3K4me3 and Decreasing H2AK119ub in Hippocampal Neurons. Front. Neurosci. 2020, 14, 591177. [Google Scholar] [CrossRef] [PubMed]
- Telegina, D.V.; Kolosova, N.G.; Kozhevnikova, O.S. Immunohistochemical Localization of NGF, BDNF, and Their Receptors in a Normal and AMD-like Rat Retina. BMC Med. Genom. 2019, 12, 48. [Google Scholar] [CrossRef]
- Wu, S.-Y.; Pan, B.-S.; Tsai, S.-F.; Chiang, Y.-T.; Huang, B.-M.; Mo, F.-E.; Kuo, Y.M. BDNF Reverses Aging-Related Microglial Activation. J. Neuroinflamm. 2020, 17, 210. [Google Scholar] [CrossRef]
- Shippy, D.C.; Wilhelm, C.; Viharkumar, P.A.; Raife, T.J.; Ulland, T.K. β-Hydroxybutyrate Inhibits Inflammasome Activation to Attenuate Alzheimer’s Disease Pathology. J. Neuroinflamm. 2020, 17, 280. [Google Scholar] [CrossRef]
- Huang, C.; Wang, P.; Xu, X.; Zhang, Y.; Gong, Y.; Hu, W.; Gao, M.; Wu, Y.; Ling, Y.; Zhao, X.; et al. The Ketone Body Metabolite β-Hydroxybutyrate Induces an Antidepression-Associated Ramification of Microglia via HDACs Inhibition-Triggered Akt-Small RhoGTPase Activation. Glia 2018, 66, 256–278. [Google Scholar] [CrossRef]
- Plaza-Zabala, A.; Sierra-Torre, V.; Sierra, A. Autophagy and Microglia: Novel Partners in Neurodegeneration and Aging. IJMS 2017, 18, 598. [Google Scholar] [CrossRef]
- Platania, C.B.M.; Maisto, R.; Trotta, M.C.; D’Amico, M.; Rossi, S.; Gesualdo, C.; D’Amico, G.; Balta, C.; Herman, H.; Hermenean, A.; et al. Retinal and circulating miRNA expression patterns in diabetic retinopathy: An in silico and in vivo approach. Br. J. Pharm. 2019, 176, 2179–2194. [Google Scholar] [CrossRef]
- Rossi, S.; Maisto, R.; Gesualdo, C.; Trotta, M.C.; Ferraraccio, F.; Kaneva, M.K.; Getting, S.J.; Surace, E.; Testa, F.; Simonelli, F.; et al. Corrigendum to “Activation of Melanocortin Receptors MC1 and MC5 Attenuates Retinal Damage in Experimental Diabetic Retinopathy”. Mediat. Inflamm. 2021, 2021, 1–2. [Google Scholar] [CrossRef]
- Bathina, S.; Das, U.N. Brain-derived neurotrophic factor and its clinical implications. Arch. Med. Sci. 2015, 11, 1164–1178. [Google Scholar] [CrossRef] [PubMed]
- Diao, J.; Liu, R.; Rong, Y.; Zhao, M.; Zhang, J.; Lai, Y.; Zhou, Q.; Wilz, L.M.; Li, J.; Vivona, S.; et al. ATG14 Promotes Membrane Tethering and Fusion of Autophagosomes to Endolysosomes. Nature 2015, 520, 563–566. [Google Scholar] [CrossRef]
- Fang, Y.; Chen, B.; Gong, A.Y.; Malhotra, D.K.; Gupta, R.; Dworkin, L.D.; Gong, R. The Ketone Body β-Hydroxybutyrate Mitigates the Senescence Response of Glomerular Podocytes to Diabetic Insults. Kidney Int. 2021, 100, 1037–1053. [Google Scholar] [CrossRef]
- Offermanns, S.; Schwaninger, M. Nutritional or Pharmacological Activation of HCA2 Ameliorates Neuroinflammation. Trends Mol. Med. 2015, 21, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Sleiman, S.F.; Henry, J.; Al-Haddad, R.; El Hayek, L.; Abou Haidar, E.; Stringer, T.; Ulja, D.; Karuppagounder, S.S.; Holson, E.B.; Ratan, R.R.; et al. Exercise promotes the expression of brain derived neurotrophic factor (BDNF) through the action of the ketone body β-hydroxybutyrate. Elife 2016, 5, e15092. [Google Scholar] [CrossRef]
- Marosi, K.; Kim, S.W.; Moehl, K.; Scheibye-Knudsen, M.; Cheng, A.; Cutler, R.; Camandola, S.; Mattson, M.P. 3-Hydroxybutyrate regulates energy metabolism and induces BDNF expression in cerebral cortical neurons. J. Neurochem. 2016, 139, 769–781. [Google Scholar] [CrossRef]
- Norgren, J.; Daniilidou, M.; Kåreholt, I.; Sindi, S.; Akenine, U.; Nordin, K.; Rosenborg, S.; Ngandu, T.; Kivipelto, M.; Sandebring-Matton, A. Serum ProBDNF Is Associated with Changes in the Ketone Body β-Hydroxybutyrate and Shows Superior Repeatability Over Mature BDNF: Secondary Outcomes from a Cross-Over Trial in Healthy Older Adults. Front. Aging Neurosci. 2021, 13, 716594. [Google Scholar] [CrossRef] [PubMed]
- Jun, Y.H.; Kim, S.T. Brain-Derived Neurotrophic Factor in Non-Proliferative Diabetic Retinopathy with Diabetic Macular Edema. Eur. J. Ophthalmol. 2021, 31, 1915–1919. [Google Scholar] [CrossRef] [PubMed]
- Walsh, J.J.; Myette-Côté, É.; Little, J.P. The Effect of Exogenous Ketone Monoester Ingestion on Plasma BDNF During an Oral Glucose Tolerance Test. Front. Physiol. 2020, 11, 1094. [Google Scholar] [CrossRef] [PubMed]
- Carretta, M.D.; Barría, Y.; Borquez, K.; Urra, B.; Rivera, A.; Alarcón, P.; Hidalgo, M.A.; Burgos, R.A. β-hydroxybutyrate and hydroxycarboxylic acid receptor 2 agonists activate the AKT, ERK and AMPK pathways, which are involved in bovine neutrophil chemotaxis. Sci. Rep. 2020, 10, 12491. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Tanihara, H.; Honda, Y.; Tatsuno, T.; Noguchi, H.; Nakayama, C. BDNF Attenuates Retinal Cell Death Caused by Chemically Induced Hypoxia in Rats. Investig. Ophthalmol. Vis. Sci. 1999, 40, 2130–2140. [Google Scholar]
- Gupta, V.; You, Y.; Li, J.; Gupta, V.; Golzan, M.; Klistorner, A.; van den Buuse, M.; Graham, S. BDNF Impairment Is Associated with Age-Related Changes in the Inner Retina and Exacerbates Experimental Glaucoma. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2014, 1842, 1567–1578. [Google Scholar] [CrossRef]
- Chen, H.; Weber, A.J. BDNF Enhances Retinal Ganglion Cell Survival in Cats with Optic Nerve Damage. Investig. Ophthalmol. Vis. Sci. 2001, 42, 966–974. [Google Scholar]
- Domenici, L.; Origlia, N.; Falsini, B.; Cerri, E.; Barloscio, D.; Fabiani, C.; Sansò, M.; Giovannini, L. Rescue of Retinal Function by BDNF in a Mouse Model of Glaucoma. PLoS ONE 2014, 9, e115579. [Google Scholar] [CrossRef]
- Martin, K.R.G.; Quigley, H.A.; Zack, D.J.; Levkovitch-Verbin, H.; Kielczewski, J.; Valenta, D.; Baumrind, L.; Pease, M.E.; Klein, R.L.; Hauswirth, W.W. Gene Therapy with Brain-Derived Neurotrophic Factor as a Protection: Retinal Ganglion Cells in a Rat Glaucoma Model. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4357. [Google Scholar] [CrossRef]
- Ren, R.; Li, Y.; Liu, Z.; Liu, K.; He, S. Long-Term Rescue of Rat Retinal Ganglion Cells and Visual Function by AAV-Mediated BDNF Expression after Acute Elevation of Intraocular Pressure. Investig. Ophthalmol. Vis. Sci. 2012, 53, 1003. [Google Scholar] [CrossRef]
- Igarashi, T.; Miyake, K.; Kobayashi, M.; Kameya, S.; Fujimoto, C.; Nakamoto, K.; Takahashi, H.; Igarashi, T.; Miyake, N.; Iijima, O.; et al. Tyrosine Triple Mutated AAV2-BDNF Gene Therapy in a Rat Model of Transient IOP Elevation. Mol. Vis. 2016, 22, 816–826. [Google Scholar]
- Osborne, A.; Khatib, T.Z.; Songra, L.; Barber, A.C.; Hall, K.; Kong, G.Y.X.; Widdowson, P.S.; Martin, K.R. Neuroprotection of Retinal Ganglion Cells by a Novel Gene Therapy Construct That Achieves Sustained Enhancement of Brain-Derived Neurotrophic Factor/Tropomyosin-Related Kinase Receptor-B Signaling. Cell Death Dis. 2018, 9, 1007. [Google Scholar] [CrossRef]
- Frake, R.A.; Ricketts, T.; Menzies, F.M.; Rubinsztein, D.C. Autophagy and Neurodegeneration. J. Clin. Investig. 2015, 125, 65–74. [Google Scholar] [CrossRef]
- Lopes de Faria, J.M.; Duarte, D.A.; Montemurro, C.; Papadimitriou, A.; Consonni, S.R.; Lopes de Faria, J.B. Defective Autophagy in Diabetic Retinopathy. Investig. Ophthalmol. Vis. Sci. 2016, 57, 4356. [Google Scholar] [CrossRef] [PubMed]
- Dehdashtian, E.; Mehrzadi, S.; Yousefi, B.; Hosseinzadeh, A.; Reiter, R.J.; Safa, M.; Ghaznavi, H.; Naseripour, M. Diabetic Retinopathy Pathogenesis and the Ameliorating Effects of Melatonin; Involvement of Autophagy, Inflammation and Oxidative Stress. Life Sci. 2018, 193, 20–33. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, S.; Anderzhanova, E.A.; Bajaj, T.; Wiechmann, S.; Dethloff, F.; Weckmann, K.; Heinz, D.E.; Ebert, T.; Hartmann, J.; Geiger, T.M.; et al. Stress-Primed Secretory Autophagy Promotes Extracellular BDNF Maturation by Enhancing MMP9 Secretion. Nat. Commun. 2021, 12, 4643. [Google Scholar] [CrossRef]
- Piano, I.; Novelli, E.; Della Santina, L.; Strettoi, E.; Cervetto, L.; Gargini, C. Involvement of Autophagic Pathway in the Progression of Retinal Degeneration in a Mouse Model of Diabetes. Front. Cell. Neurosci. 2016, 10, 42. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Wang, Q.; Wan, D.; Sun, Y.; Wang, L.; Chen, H.; Liu, C.; Petersen, R.B.; Li, J.; Xue, W.; et al. Histone HIST1H1C/H1.2 Regulates Autophagy in the Development of Diabetic Retinopathy. Autophagy 2017, 13, 941–954. [Google Scholar] [CrossRef]
- Cai, X.; Li, J.; Wang, M.; She, M.; Tang, Y.; Li, J.; Li, H.; Hui, H. GLP-1 Treatment Improves Diabetic Retinopathy by Alleviating Autophagy through GLP-1R-ERK1/2-HDAC6 Signaling Pathway. Int. J. Med. Sci. 2017, 14, 1203–1212. [Google Scholar] [CrossRef]
- Park, H.-Y.L.; Kim, J.H.; Park, C.K. Different Contributions of Autophagy to Retinal Ganglion Cell Death in the Diabetic and Glaucomatous Retinas. Sci. Rep. 2018, 8, 13321. [Google Scholar] [CrossRef]
- Du, M.; Wu, M.; Fu, D.; Yang, S.; Chen, J.; Wilson, K.; Lyons, T.J. Effects of Modified LDL and HDL on Retinal Pigment Epithelial Cells: A Role in Diabetic Retinopathy? Diabetologia 2013, 56, 2318–2328. [Google Scholar] [CrossRef]
- Yao, J.; Tao, Z.-F.; Li, C.-P.; Li, X.-M.; Cao, G.-F.; Jiang, Q.; Yan, B. Regulation of Autophagy by High Glucose in Human Retinal Pigment Epithelium. Cell. Physiol. Biochem. 2014, 33, 107–116. [Google Scholar] [CrossRef]
- Shi, H.; Zhang, Z.; Wang, X.; Li, R.; Hou, W.; Bi, W.; Zhang, X. Inhibition of Autophagy Induces IL-1β Release from ARPE-19 Cells via ROS Mediated NLRP3 Inflammasome Activation under High Glucose Stress. Biochem. Biophys. Res. Commun. 2015, 463, 1071–1076. [Google Scholar] [CrossRef]
- Fu, D.; Yu, J.Y.; Connell, A.R.; Yang, S.; Hookham, M.B.; McLeese, R.; Lyons, T.J. Beneficial Effects of Berberine on Oxidized LDL-Induced Cytotoxicity to Human Retinal Müller Cells. Investig. Ophthalmol. Vis. Sci. 2016, 57, 3369. [Google Scholar] [CrossRef] [PubMed]
- Fu, D.; Yu, J.Y.; Yang, S.; Wu, M.; Hammad, S.M.; Connell, A.R.; Du, M.; Chen, J.; Lyons, T.J. Survival or Death: A Dual Role for Autophagy in Stress-Induced Pericyte Loss in Diabetic Retinopathy. Diabetologia 2016, 59, 2251–2261. [Google Scholar] [CrossRef] [PubMed]
- Madrakhimov, S.B.; Yang, J.Y.; Kim, J.H.; Han, J.W.; Park, T.K. MTOR-Dependent Dysregulation of Autophagy Contributes to the Retinal Ganglion Cell Loss in Streptozotocin-Induced Diabetic Retinopathy. Cell Commun. Signal. 2021, 19, 29. [Google Scholar] [CrossRef] [PubMed]
- Camberos-Luna, L.; Gerónimo-Olvera, C.; Montiel, T.; Rincon-Heredia, R.; Massieu, L. The Ketone Body, β-Hydroxybutyrate Stimulates the Autophagic Flux and Prevents Neuronal Death Induced by Glucose Deprivation in Cortical Cultured Neurons. Neurochem. Res. 2016, 41, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Nikoletopoulou, V.; Sidiropoulou, K.; Kallergi, E.; Dalezios, Y.; Tavernarakis, N. Modulation of Autophagy by BDNF Underlies Synaptic Plasticity. Cell Metab. 2017, 26, 230–242.e5. [Google Scholar] [CrossRef]
- Torres-Esquivel, C.; Montiel, T.; Flores-Méndez, M.; Massieu, L. Effect of β-Hydroxybutyrate on Autophagy Dynamics During Severe Hypoglycemia and the Hypoglycemic Coma. Front. Cell. Neurosci. 2020, 14, 547215. [Google Scholar] [CrossRef]
- Chen, X.; Gu, X.; Zhang, H. Sidt2 regulates hepatocellular lipid metabolism through autophagy. J. Lipid Res. 2018, 59, 404–415. [Google Scholar] [CrossRef]
- Ho, C.-F.; Chan, K.-W.; Yeh, H.-I.; Kuo, J.; Liu, H.-J.; Wang, C.-Y. Ketone Bodies Upregulate Endothelial Connexin 43 (Cx43) Gap Junctions. Vet. J. 2013, 198, 696–701. [Google Scholar] [CrossRef]
- Choi, I.; Zhang, Y.; Seegobin, S.P.; Pruvost, M.; Wang, Q.; Purtell, K.; Zhang, B.; Yue, Z. Microglia Clear Neuron-Released α-Synuclein via Selective Autophagy and Prevent Neurodegeneration. Nat. Commun. 2020, 11, 1386. [Google Scholar] [CrossRef]
- Pomilio, C.; Gorojod, R.M.; Riudavets, M.; Vinuesa, A.; Presa, J.; Gregosa, A.; Bentivegna, M.; Alaimo, A.; Alcon, S.P.; Sevlever, G.; et al. Microglial Autophagy Is Impaired by Prolonged Exposure to β-Amyloid Peptides: Evidence from Experimental Models and Alzheimer’s Disease Patients. GeroScience 2020, 42, 613–632. [Google Scholar] [CrossRef]
- Folick, A.; Koliwad, S.K.; Valdearcos, M. Microglial Lipid Biology in the Hypothalamic Regulation of Metabolic Homeostasis. Front. Endocrinol. 2021, 12, 668396. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Hamanaka, G.; Lo, E.H.; Arai, K. Heterogeneity of Microglia and Their Differential Roles in White Matter Pathology. CNS Neurosci Ther. 2019, 25, 1290–1298. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Wong, W.T. Aging Changes in Retinal Microglia and Their Relevance to Age-Related Retinal Disease. In Retinal Degenerative Diseases; Bowes Rickman, C., LaVail, M.M., Anderson, R.E., Grimm, C., Hollyfield, J., Ash, J., Eds.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2016; Volume 854, pp. 73–78. ISBN 978-3-319-17120-3. [Google Scholar]
- Wu, H.; Wang, M.; Li, X.; Shao, Y. The Metaflammatory and Immunometabolic Role of Macrophages and Microglia in Diabetic Retinopathy. Hum. Cell 2021, 34, 1617–1628. [Google Scholar] [CrossRef]
- Cochran, J.; Taufalele, P.V.; Lin, K.D.; Zhang, Y.; Abel, E.D. Sex Differences in the Response of C57BL/6 Mice to Ketogenic Diets. Diabetes 2018, 67, 1884-P. [Google Scholar] [CrossRef]
- Sheikh-Ali, M.; Karon, B.S.; Basu, A.; Kudva, Y.C.; Muller, L.A.; Xu, J.; Schwenk, W.F.; Miles, J.M. Can serum beta-hydroxybutyrate be used to diagnose diabetic ketoacidosis? Diabetes Care 2008, 31, 643–647. [Google Scholar] [CrossRef] [PubMed]
- Adams, J. The proteasome: Structure, function, and role in the cell. Cancer Treat. Rev. 2003, 29 (Suppl. 1), 3–9. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, W.; Du, B.; Miao, X.; Bai, Y.; Xin, Y.; Tan, Y.; Cui, W.; Liu, B.; Cui, T.; et al. Therapeutic effect of MG-132 on diabetic cardiomyopathy is associated with its suppression of proteasomal activities: Roles of Nrf2 and NF-κB. Am. J. Physiol. Heart Circ. Physiol. 2013, 304, H567–H578. [Google Scholar] [CrossRef]
- Yan, Y.; Jiang, K.; Liu, P.; Zhang, X.; Dong, X.; Gao, J.; Liu, Q.; Barr, M.P.; Zhang, Q.; Hou, X.; et al. Bafilomycin A1 induces caspase-independent cell death in hepatocellular carcinoma cells via targeting of autophagy and MAPK pathways. Sci. Rep. 2016, 6, 37052. [Google Scholar] [CrossRef]
- Yuan, N.; Song, L.; Zhang, S.; Lin, W.; Cao, Y.; Xu, F.; Fang, Y.; Wang, Z.; Zhang, H.; Li, X.; et al. Bafilomycin A1 targets both autophagy and apoptosis pathways in pediatric B-cell acute lymphoblastic leukemia. Haematologica 2015, 100, 345–356. [Google Scholar] [CrossRef]
- Moulis, M.; Vindis, C. Methods for Measuring Autophagy in Mice. Cells 2017, 6, 14. [Google Scholar] [CrossRef]
- Iwai-Kanai, E.; Yuan, H.; Huang, C.; Sayen, M.R.; Perry-Garza, C.N.; Kim, L.; Gottlieb, R.A. A method to measure cardiac autophagic flux in vivo. Autophagy 2008, 4, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Trotta, M.C.; Gharbia, S.; Herman, H.; Mladin, B.; Hermenean, A.; Balta, C.; Cotoraci, C.; Peteu, V.E.; Gesualdo, C.; Petrillo, F.; et al. Sex and Age-Related Differences in Neuroinflammation and Apoptosis in Balb/c Mice Retina Involve Resolvin D1. Int. J. Mol. Sci. 2021, 22, 6280. [Google Scholar] [CrossRef] [PubMed]
- Su, S.B.; Silver, P.B.; Zhang, M.; Chan, C.-C.; Caspi, R.R. Pertussis Toxin Inhibits Induction of Tissue-Specific Autoimmune Disease by Disrupting G Protein-Coupled Signals. J. Immunol. 2001, 167, 250–256. [Google Scholar] [CrossRef]
- Yadav, R.; Bhowmick, S.; Gorecki, P.; O’Rourke, J.; Cone, R.E. Paradoxical Effect of Pertussis Toxin on the Delayed Hypersensitivity Response to Autoantigens in Mice. PLoS ONE 2010, 5, e11983. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trotta, M.C.; Gesualdo, C.; Herman, H.; Gharbia, S.; Balta, C.; Lepre, C.C.; Russo, M.; Itro, A.; D’Amico, G.; Peluso, L.; et al. Systemic Beta-Hydroxybutyrate Affects BDNF and Autophagy into the Retina of Diabetic Mice. Int. J. Mol. Sci. 2022, 23, 10184. https://doi.org/10.3390/ijms231710184
Trotta MC, Gesualdo C, Herman H, Gharbia S, Balta C, Lepre CC, Russo M, Itro A, D’Amico G, Peluso L, et al. Systemic Beta-Hydroxybutyrate Affects BDNF and Autophagy into the Retina of Diabetic Mice. International Journal of Molecular Sciences. 2022; 23(17):10184. https://doi.org/10.3390/ijms231710184
Chicago/Turabian StyleTrotta, Maria Consiglia, Carlo Gesualdo, Hildegard Herman, Sami Gharbia, Cornel Balta, Caterina Claudia Lepre, Marina Russo, Annalisa Itro, Giovanbattista D’Amico, Luisa Peluso, and et al. 2022. "Systemic Beta-Hydroxybutyrate Affects BDNF and Autophagy into the Retina of Diabetic Mice" International Journal of Molecular Sciences 23, no. 17: 10184. https://doi.org/10.3390/ijms231710184