
Imidacloprid Impairs Glutamatergic Synaptic Plasticity and Desensitizes Mechanosensitive, Nociceptive, and Photogenic Response of Drosophila melanogaster by Mediating Oxidative Stress, Which Could Be Rescued by Osthole
 and
and 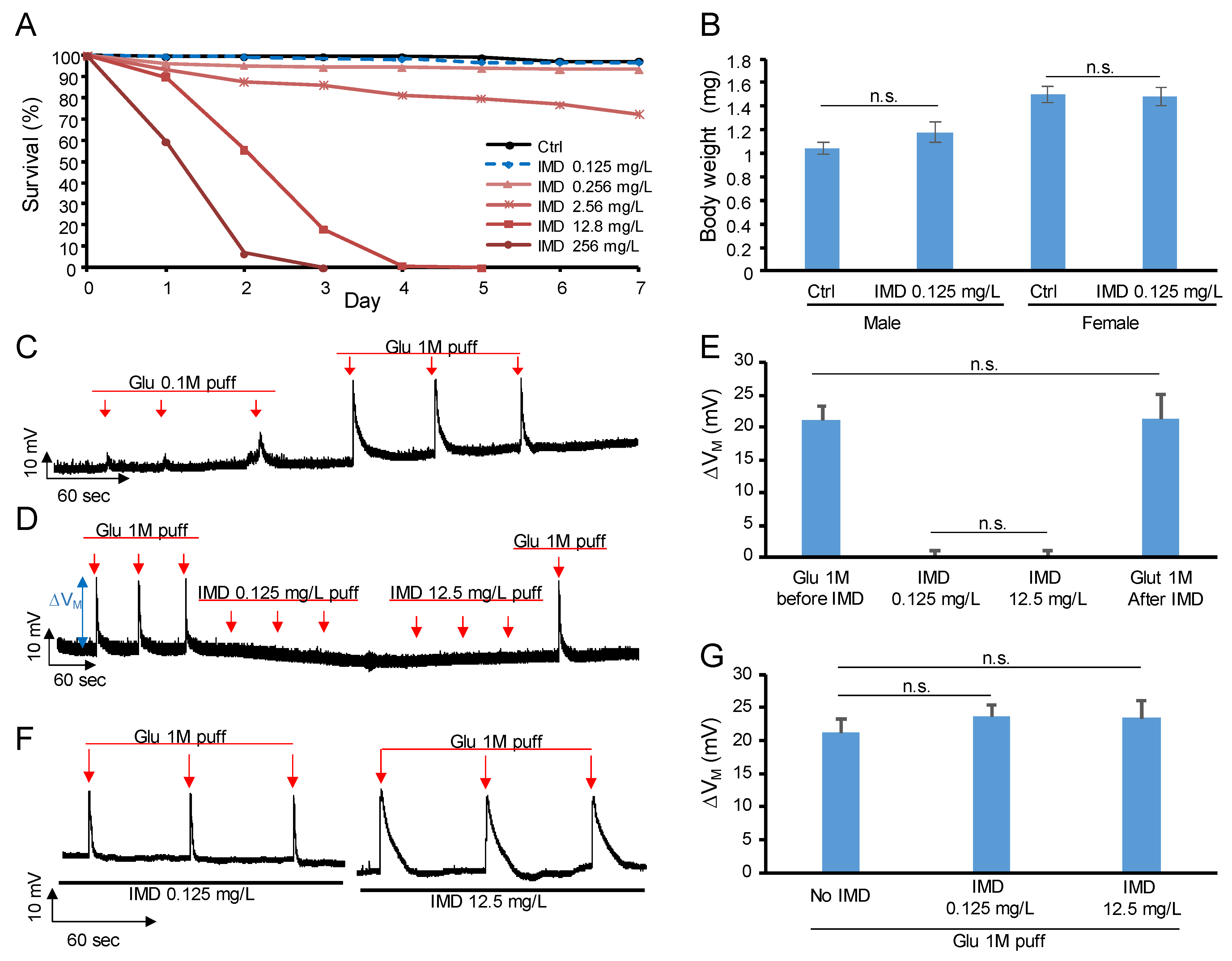{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Determine the IMD Dose
2.2. IMD Is Neither Agonist Nor Antagonist of Glutamate Receptor
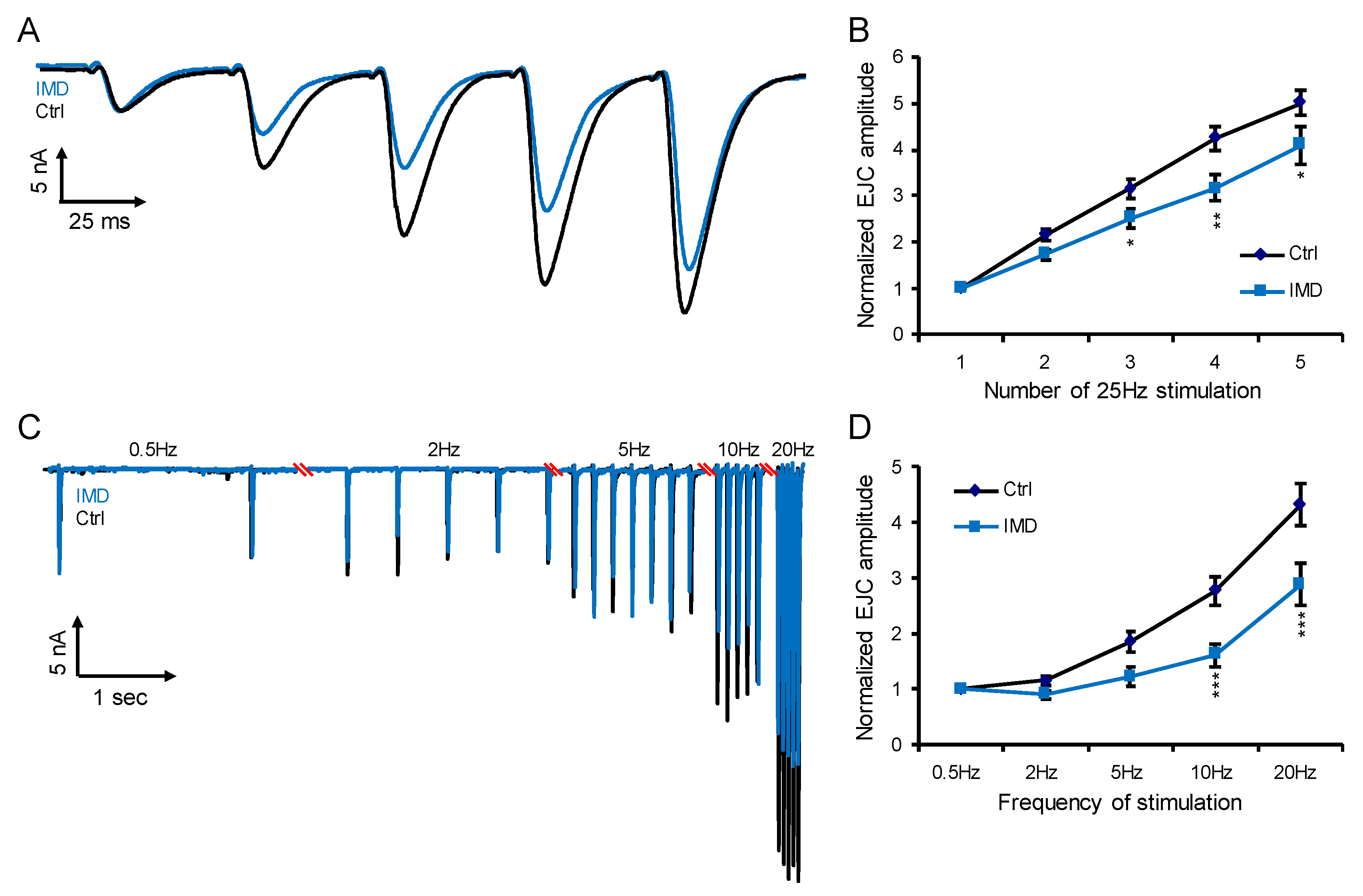
2.3. Chronic IMD Treatment Damps the Glutamatergic Synaptic Plasticity
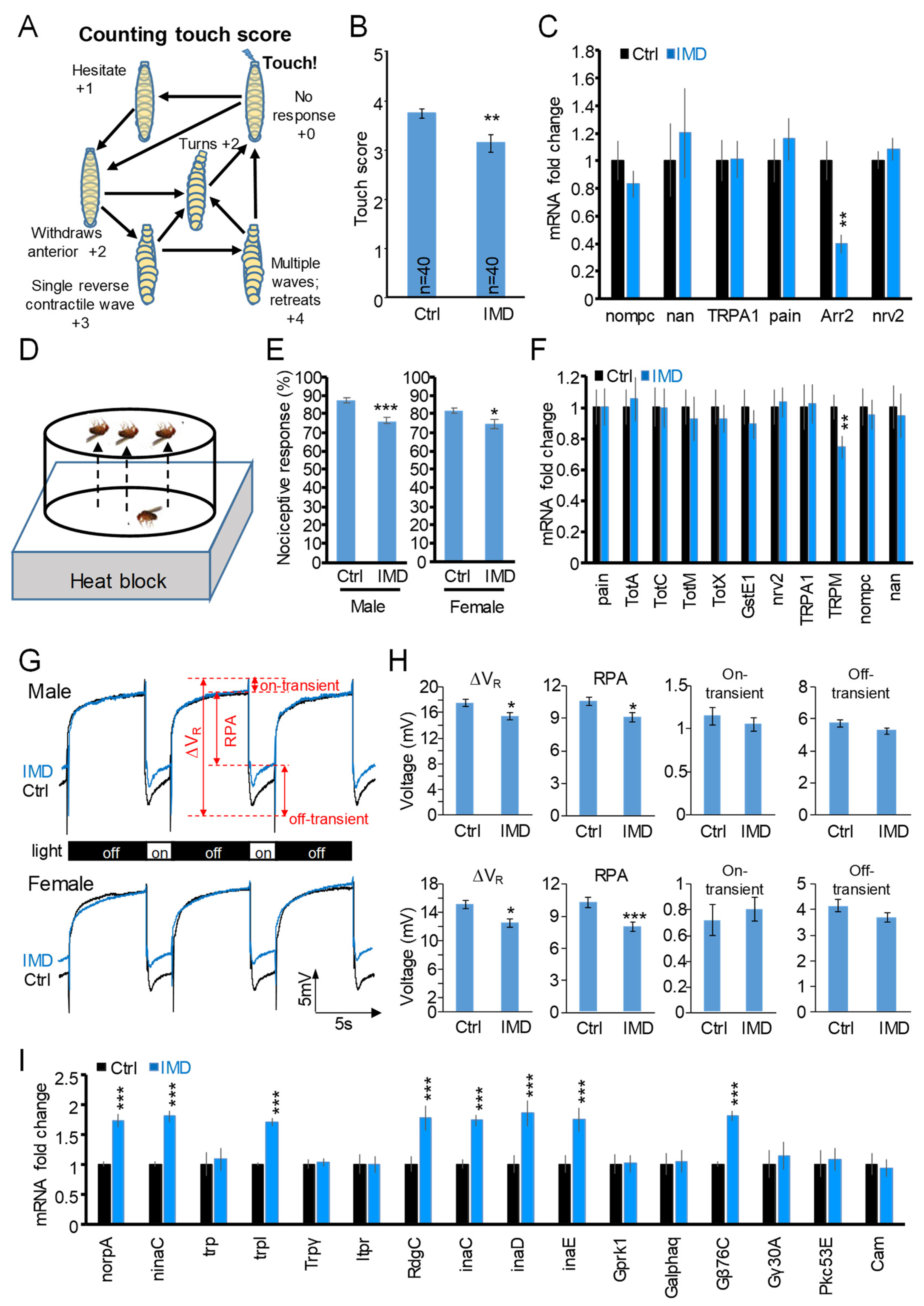
2.4. Chronic IMD Treatment Evokes Nerve-Conduction-Associated Behavioral Phenotypes
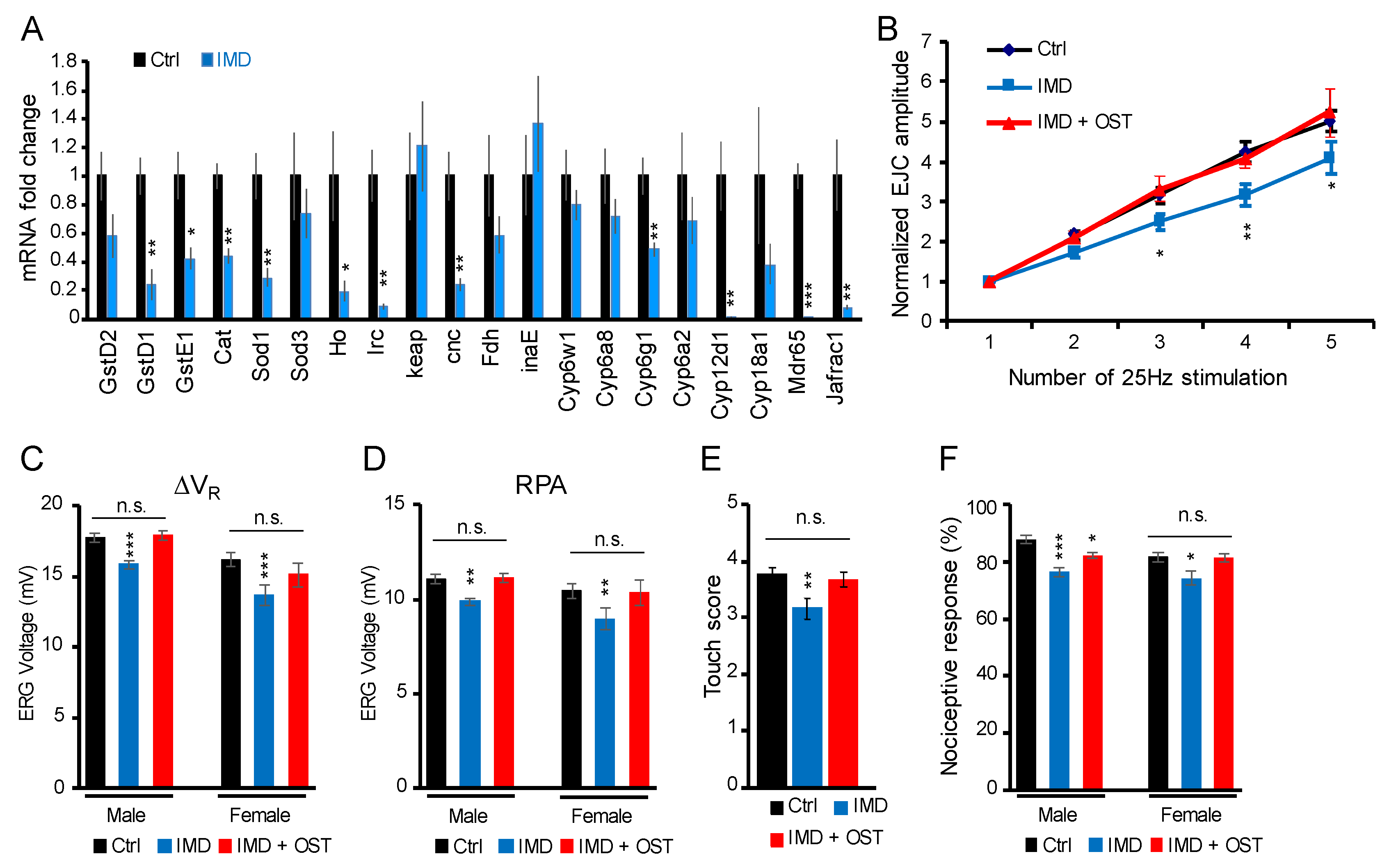
2.5. IMD-Induced Phenotypes Were Rescued by Antioxidant Osthole
3. Discussion
4. Materials and Methods
4.1. Chemicals and Fly Strain
4.2. NMJ Electrophysiology
4.2.1. Larvae Sample Preparation
4.2.2. Bridge Mode Recording of Membrane Potential
4.2.3. Two-Electrode Voltage-Clamp Recording of STF
4.3. Behavioral Assays
4.3.1. Survival Assay
4.3.2. Body Weight
4.3.3. Electroretinogram (ERG)
4.3.4. Mechanosensory Response
4.3.5. Nociception
4.4. RNA Extraction and RT-qPCR
4.5. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jeschke, P.; Nauen, R. Neonicotinoids—From zero to hero in insecticide chemistry. Pest Manag. Sci. Former. Pestic. Sci. 2008, 64, 1084–1098. [Google Scholar] [CrossRef] [PubMed]
- Bass, C.; Denholm, I.; Williamson, M.S.; Nauen, R. The global status of insect resistance to neonicotinoid insecticides. Pestic. Biochem. Physiol. 2015, 121, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Crossthwaite, A.J.; Rendine, S.; Stenta, M.; Slater, R. Target-site resistance to neonicotinoids. J. Chem. Biol. 2014, 7, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Déglise, P.; Grünewald, B.; Gauthier, M. The insecticide imidacloprid is a partial agonist of the nicotinic receptor of honeybee Kenyon cells. Neurosci. Lett. 2002, 321, 13–16. [Google Scholar] [CrossRef]
- Uhl, P.; Bucher, R.; Schäfer, R.B.; Entling, M.H. Sublethal effects of imidacloprid on interactions in a tritrophic system of non-target species. Chemosphere 2015, 132, 152–158. [Google Scholar]
- Sur, R.; Stork, A. Uptake, translocation and metabolism of imidacloprid in plants. Bull. Insectology 2003, 56, 35–40. [Google Scholar]
- Goulson, D. Pesticides linked to bird declines. Nature 2014, 511, 295–296. [Google Scholar] [CrossRef]
- Elbert, A.; Haas, M.; Springer, B.; Thielert, W.; Nauen, R. Applied aspects of neonicotinoid uses in crop protection. Pest Manag. Sci. Former. Pestic. Sci. 2008, 64, 1099–1105. [Google Scholar]
- Chen, M.; Tao, L.; McLean, J.; Lu, C. Quantitative analysis of neonicotinoid insecticide residues in foods: Implication for dietary exposures. J. Agric. Food Chem. 2014, 62, 6082–6090. [Google Scholar] [CrossRef]
- Mitchell, E.A.; Mulhauser, B.; Mulot, M.; Mutabazi, A.; Glauser, G.; Aebi, A. A worldwide survey of neonicotinoids in honey. Science 2017, 358, 109–111. [Google Scholar]
- Thunnissen, N.; Lautz, L.; van Schaik, T.; Hendriks, A. Ecological risks of imidacloprid to aquatic species in the Netherlands: Measured and estimated concentrations compared to species sensitivity distributions. Chemosphere 2020, 254, 126604. [Google Scholar]
- Morrissey, C.A.; Mineau, P.; Devries, J.H.; Sanchez-Bayo, F.; Liess, M.; Cavallaro, M.C.; Liber, K. Neonicotinoid contamination of global surface waters and associated risk to aquatic invertebrates: A review. Environ. Int. 2015, 74, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Lu, Z.; Lin, S.; Dai, W.; Zhang, Q. Neonicotinoid insecticides in the drinking water system–Fate, transportation, and their contributions to the overall dietary risks. Environ. Pollut. 2020, 258, 113722. [Google Scholar] [CrossRef] [PubMed]
- Morrison, L.M.; Renaud, J.B.; Sabourin, L.; Sumarah, M.W.; Yeung, K.K.; Lapen, D.R. High-throughput quantitation of neonicotinoids in lyophilized surface water by LC-APCI-MS/MS. J. AOAC Int. 2018, 101, 1940–1947. [Google Scholar] [CrossRef]
- Martelli, F.; Zhongyuan, Z.; Wang, J.; Wong, C.-O.; Karagas, N.E.; Roessner, U.; Rupasinghe, T.; Venkatachalam, K.; Perry, T.; Bellen, H.J. Low doses of the neonicotinoid insecticide imidacloprid induce ROS triggering neurological and metabolic impairments in Drosophila. Proc. Natl. Acad. Sci. USA 2020, 117, 25840–25850. [Google Scholar] [CrossRef]
- Mikolić, A.; Karačonji, I.B. Imidacloprid as reproductive toxicant and endocrine disruptor: Investigations in laboratory animals. Arh. Hig. Rada Toksikol. 2018, 69, 103–108. [Google Scholar] [CrossRef]
- Young, H.K.; Denecke, S.M.; Robin, C.; Fournier-Level, A. Sublethal larval exposure to imidacloprid impacts adult behaviour in Drosophila melanogaster. J. Evol. Biol. 2020, 33, 151–164. [Google Scholar] [CrossRef]
- Yuan, X.; Shen, J.; Zhang, X.; Tu, W.; Fu, Z.; Jin, Y. Imidacloprid disrupts the endocrine system by interacting with androgen receptor in male mice. Sci. Total Environ. 2020, 708, 135163. [Google Scholar] [CrossRef]
- Vohra, P.; Khera, K.S. Effect of imidacloprid on reproduction of female albino rats in three generation study. J. Vet. Sci. Technol. 2016, 7, 1000340. [Google Scholar] [CrossRef]
- Li, P.; Ann, J.; Akk, G. Activation and modulation of human α4β2 nicotinic acetylcholine receptors by the neonicotinoids clothianidin and imidacloprid. J. Neurosci. Res. 2011, 89, 1295–1301. [Google Scholar] [CrossRef]
- Collingridge, G.L.; Olsen, R.W.; Peters, J.; Spedding, M. A nomenclature for ligand-gated ion channels. Neuropharmacology 2009, 56, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.K. Genomics, cys-loop ligand-gated ion channels and new targets for the control of insect pests and vectors. Curr. Opin. Insect Sci. 2018, 30, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Low, C.-M.; Moody, O.A.; Jenkins, A.; Traynelis, S.F. Ionotropic GABA and glutamate receptor mutations and human neurologic diseases. Mol. Pharmacol. 2015, 88, 203–217. [Google Scholar] [CrossRef] [PubMed]
- Keil, A.P.; Daniels, J.L.; Hertz-Picciotto, I. Autism spectrum disorder, flea and tick medication, and adjustments for exposure misclassification: The CHARGE (CHildhood Autism Risks from Genetics and Environment) case–control study. Environ. Health 2014, 13, 3. [Google Scholar] [CrossRef]
- Decourtye, A.; Armengaud, C.; Renou, M.; Devillers, J.; Cluzeau, S.; Gauthier, M.; Pham-Delègue, M.-H. Imidacloprid impairs memory and brain metabolism in the honeybee (Apis mellifera L.). Pestic. Biochem. Physiol. 2004, 78, 83–92. [Google Scholar] [CrossRef]
- Hsiao, C.-J.; Lin, C.-L.; Lin, T.-Y.; Wang, S.-E.; Wu, C.-H. Imidacloprid toxicity impairs spatial memory of echolocation bats through neural apoptosis in hippocampal CA1 and medial entorhinal cortex areas. Neuroreport 2016, 27, 462–468. [Google Scholar] [CrossRef]
- Kara, M.; Yumrutas, O.; Demir, C.F.; Ozdemir, H.H.; Bozgeyik, I.; Coskun, S.; Eraslan, E.; Bal, R. Insecticide imidacloprid influences cognitive functions and alters learning performance and related gene expression in a rat model. Int. J. Exp. Pathol. 2015, 96, 332–337. [Google Scholar] [CrossRef]
- Riedel, G.; Platt, B.; Micheau, J. Glutamate receptor function in learning and memory. Behav. Brain Res. 2003, 140, 1–47. [Google Scholar] [CrossRef]
- Jan, L.; Jan, Y. L-glutamate as an excitatory transmitter at the Drosophila larval neuromuscular junction. J. Physiol. 1976, 262, 215–236. [Google Scholar] [CrossRef]
- Zhang, B.; Stewart, B. Electrophysiological recording from Drosophila larval body-wall muscles. Cold Spring Harb. Protoc. 2010, 2010, pdb.prot5487. [Google Scholar] [CrossRef]
- Jan, Y.N.; Jan, L.Y. Genetic dissection of short-term and long-term facilitation at the Drosophila neuromuscular junction. Proc. Natl. Acad. Sci. USA 1978, 75, 515–519. [Google Scholar] [CrossRef] [PubMed]
- Haugland, F.N.; Wu, C. A voltage-clamp analysis of gene-dosage effects of the Shaker locus on larval muscle potassium currents in Drosophila. J. Neurosci. 1990, 10, 1357–1371. [Google Scholar] [CrossRef] [PubMed]
- Knecht, Z.A.; Silbering, A.F.; Ni, L.; Klein, M.; Budelli, G.; Bell, R.; Abuin, L.; Ferrer, A.J.; Samuel, A.D.; Benton, R. Distinct combinations of variant ionotropic glutamate receptors mediate thermosensation and hygrosensation in Drosophila. eLife 2016, 5, e17879. [Google Scholar] [PubMed]
- Montell, C. Drosophila TRP channels. Pflüg. Arch. 2005, 451, 19–28. [Google Scholar] [CrossRef]
- Kernan, M.; Cowan, D.; Zuker, C. Genetic dissection of mechanosensory transduction: Mechanoreception-defective mutations of Drosophila. Neuron 1994, 12, 1195–1206. [Google Scholar] [CrossRef]
- Pierce, K.L.; Lefkowitz, R.J. Classical and new roles of β-arrestins in the regulation of G-protein-coupled receptors. Nat. Rev. Neurosci. 2001, 2, 727–733. [Google Scholar] [CrossRef]
- Balieira, K.V.B.; Mazzo, M.; Bizerra, P.F.V.; de Jesus Santos Guimarães, A.R.; Nicodemo, D.; Mingatto, F.E. Imidacloprid-induced oxidative stress in honey bees and the antioxidant action of caffeine. Apidologie 2018, 49, 562–572. [Google Scholar] [CrossRef]
- El-Gendy, K.S.; Aly, N.M.; Mahmoud, F.H.; Kenawy, A.; El-Sebae, A.K.H. The role of vitamin C as antioxidant in protection of oxidative stress induced by imidacloprid. Food Chem. Toxicol. 2010, 48, 215–221. [Google Scholar] [CrossRef]
- Yang, S.-M.; Chan, Y.-L.; Hua, K.-F.; Chang, J.-M.; Chen, H.-L.; Tsai, Y.-J.; Hsu, Y.-J.; Chao, L.K.; Feng-Ling, Y.; Tsai, Y.-L. Osthole improves an accelerated focal segmental glomerulosclerosis model in the early stage by activating the Nrf2 antioxidant pathway and subsequently inhibiting NF-κB-mediated COX-2 expression and apoptosis. Free. Radic. Biol. Med. 2014, 73, 260–269. [Google Scholar] [CrossRef]
- Ryzhova, I.; Zachepilo, T.; Chesnokova, E.; Lopatina, N. Metabotropic glutamate receptors in mechanisms of plasticity of the central nervous system in the honeybee Apis mellifera. J. Evol. Biochem. Physiol. 2010, 46, 251–258. [Google Scholar] [CrossRef]
- Ramirez-Amaya, V. Neural Plasticity and Memory: From Genes to Brain Imaging; CRC Press: Boca Raton, FL, USA, 2011; pp. 47–59. [Google Scholar]
- Burnett, E.J.; Chandler, L.J.; Trantham-Davidson, H. Glutamatergic plasticity and alcohol dependence-induced alterations in reward, affect and cognition. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2016, 65, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Quintero, G.C. Role of nucleus accumbens glutamatergic plasticity in drug addiction. Neuropsychiatr. Dis. Treat. 2013, 9, 1499. [Google Scholar] [CrossRef]
- Bazzari, A.H.; Parri, H.R. Neuromodulators and long-term synaptic plasticity in learning and memory: A steered-glutamatergic perspective. Brain Sci. 2019, 9, 300. [Google Scholar] [CrossRef] [PubMed]
- Fowler, M.A.; Montell, C. Drosophila TRP channels and animal behavior. Life Sci. 2013, 92, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Turner, H.N.; Armengol, K.; Patel, A.A.; Himmel, N.J.; Sullivan, L.; Iyer, S.C.; Bhattacharya, S.; Iyer, E.P.R.; Landry, C.; Galko, M.J. The TRP channels Pkd2, NompC, and Trpm act in cold-sensing neurons to mediate unique aversive behaviors to noxious cold in Drosophila. Curr. Biol. 2016, 26, 3116–3128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, R.I.; Corey, D.P. The force be with you: A mechanoreceptor channel in proprioception and touch. Neuron 2010, 67, 349–351. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Li, Z.; Jin, M.; Yin, Y.-L.; de Waal, P.W.; Pal, K.; Yin, Y.; Gao, X.; He, Y.; Gao, J. A complex structure of arrestin-2 bound to a G protein-coupled receptor. Cell Res. 2019, 29, 971–983. [Google Scholar] [CrossRef]
- Zhu, L.; Almaça, J.; Dadi, P.K.; Hong, H.; Sakamoto, W.; Rossi, M.; Lee, R.J.; Vierra, N.C.; Lu, H.; Cui, Y. β-arrestin-2 is an essential regulator of pancreatic β-cell function under physiological and pathophysiological conditions. Nat. Commun. 2017, 8, 14295. [Google Scholar] [CrossRef]
- Luan, B.; Zhao, J.; Wu, H.; Duan, B.; Shu, G.; Wang, X.; Li, D.; Jia, W.; Kang, J.; Pei, G. Deficiency of a β-arrestin-2 signal complex contributes to insulin resistance. Nature 2009, 457, 1146–1149. [Google Scholar] [CrossRef]
- Du, J.-J.; Sun, J.-C.; Li, N.; Li, X.-Q.; Sun, W.-Y.; Wei, W. β-Arrestin2 deficiency attenuates oxidative stress in mouse hepatic fibrosis through modulation of NOX4. Acta Pharmacol. Sin. 2021, 42, 1090–1100. [Google Scholar] [CrossRef]
- Khalil, S.R.; Awad, A.; Mohammed, H.H.; Nassan, M.A. Imidacloprid insecticide exposure induces stress and disrupts glucose homeostasis in male rats. Environ. Toxicol. Pharmacol. 2017, 55, 165–174. [Google Scholar] [CrossRef]
- Chakroun, S.; Grissa, I.; Ezzi, L.; Ammar, O.; Neffati, F.; Kerkeni, E.; Najjar, M.F.; Haouas, Z.; Cheikh, H.B. Imidacloprid enhances liver damage in Wistar rats: Biochemical, oxidative damage and histological assessment. J. Coast. Life Med. 2017, 5, 540–546. [Google Scholar] [CrossRef]
- Butcherine, P.; Kelaher, B.P.; Benkendorff, K. Assessment of acetylcholinesterase, catalase, and glutathione S-transferase as biomarkers for imidacloprid exposure in penaeid shrimp. Aquat. Toxicol. 2022, 242, 106050. [Google Scholar] [CrossRef]
- Duzguner, V.; Erdogan, S. Chronic exposure to imidacloprid induces inflammation and oxidative stress in the liver & central nervous system of rats. Pestic. Biochem. Physiol. 2012, 104, 58–64. [Google Scholar]
- Chen, M.-Y.; Liu, H.-P.; Cheng, J.; Chiang, S.-Y.; Liao, W.-P.; Lin, W.-Y. Transgenerational impact of DEHP on body weight of Drosophila. Chemosphere 2019, 221, 493–499. [Google Scholar] [CrossRef]
- Liu, H.-P.; Cheng, J.; Chen, M.-Y.; Chuang, T.-N.; Dong, J.-C.; Liu, C.-H.; Lin, W.-Y. Neuromuscular, retinal, and reproductive impact of low-dose polystyrene microplastics on Drosophila. Environ. Pollut. 2022, 292, 118455. [Google Scholar] [CrossRef]
- Chen, M.-Y.; Liu, H.-P.; Liu, C.-H.; Cheng, J.; Chang, M.-S.; Chiang, S.-Y.; Liao, W.-P.; Lin, W.-Y. DEHP toxicity on vision, neuromuscular junction, and courtship behaviors of Drosophila. Environ. Pollut. 2018, 243, 1558–1567. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.-H.; Chen, M.-Y.; Cheng, J.; Chuang, T.-N.; Liu, H.-P.; Lin, W.-Y. Imidacloprid Impairs Glutamatergic Synaptic Plasticity and Desensitizes Mechanosensitive, Nociceptive, and Photogenic Response of Drosophila melanogaster by Mediating Oxidative Stress, Which Could Be Rescued by Osthole. Int. J. Mol. Sci. 2022, 23, 10181. https://doi.org/10.3390/ijms231710181
Liu C-H, Chen M-Y, Cheng J, Chuang T-N, Liu H-P, Lin W-Y. Imidacloprid Impairs Glutamatergic Synaptic Plasticity and Desensitizes Mechanosensitive, Nociceptive, and Photogenic Response of Drosophila melanogaster by Mediating Oxidative Stress, Which Could Be Rescued by Osthole. International Journal of Molecular Sciences. 2022; 23(17):10181. https://doi.org/10.3390/ijms231710181
Chicago/Turabian StyleLiu, Chuan-Hsiu, Mei-Ying Chen, Jack Cheng, Tsai-Ni Chuang, Hsin-Ping Liu, and Wei-Yong Lin. 2022. "Imidacloprid Impairs Glutamatergic Synaptic Plasticity and Desensitizes Mechanosensitive, Nociceptive, and Photogenic Response of Drosophila melanogaster by Mediating Oxidative Stress, Which Could Be Rescued by Osthole" International Journal of Molecular Sciences 23, no. 17: 10181. https://doi.org/10.3390/ijms231710181