
Gut Metabolites and Breast Cancer: The Continuum of Dysbiosis, Breast Cancer Risk, and Potential Breast Cancer Therapy



Abstract
:1. Introduction
1.1. The Gut Microbiota and Its Metabolites
1.2. The Crosstalk between Gut Metabolites and the Host Immune System
2. Methods
2.1. Search Strategy
2.2. Data Extraction
3. Results

{kind=link}
{kind=link}
{kind=link}
{kind=link}
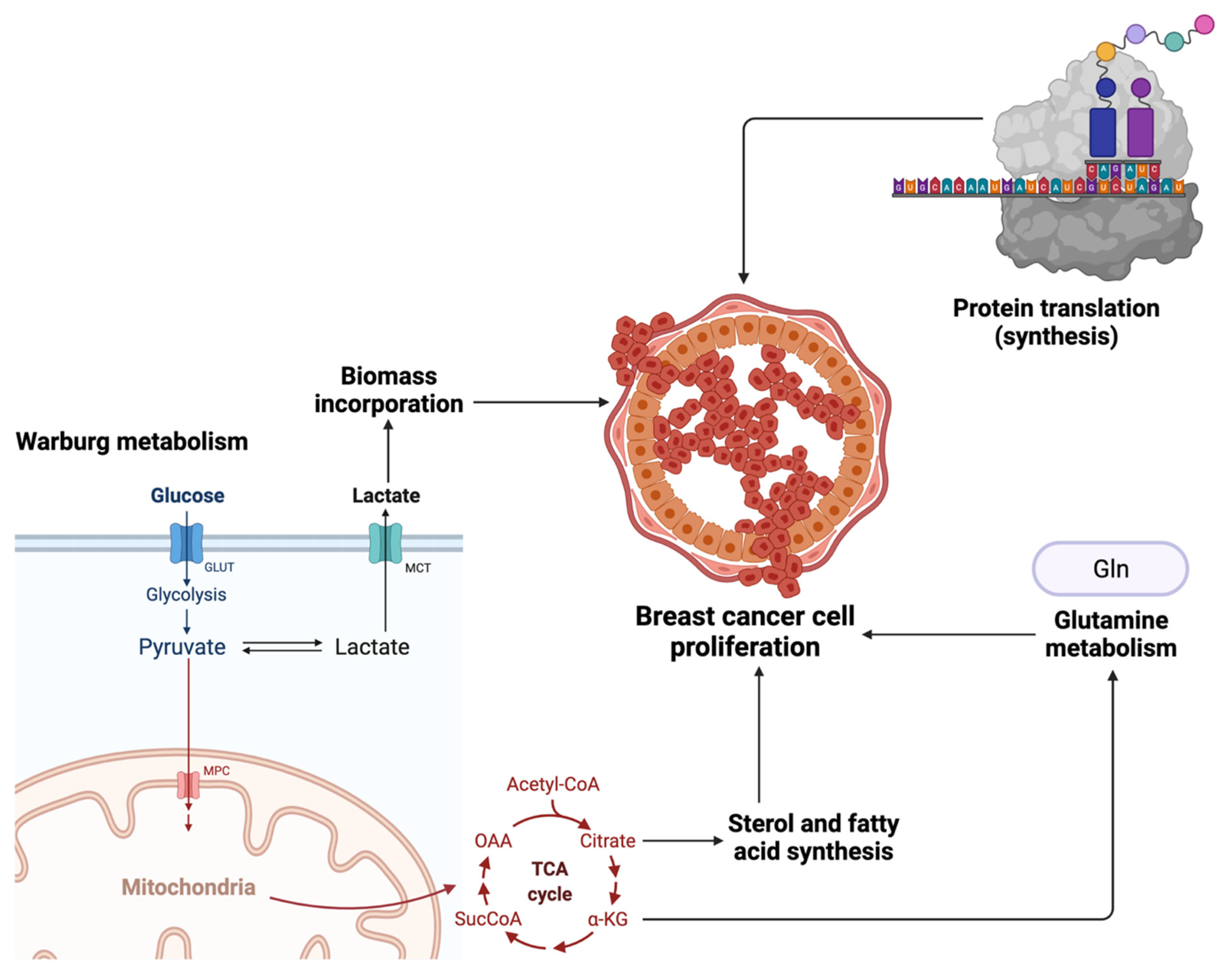{kind=link}
{kind=link}
{kind=link}
| Metabolite Group | Metabolite | Cancer Type | Type of Study | Cancer Cell Line/Animal Type | Type of Assay | Inhibitory Effect | Reference |
|---|---|---|---|---|---|---|---|
| Bacteriocin | Nisin | Breast | In vitro | MCF7 | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) | High cytotoxicity with the IC50 value of 5 μM, and selectivity against the MCF7 cells. | [16] |
| Nisin | Breast | In vitro | MCF7 | MTT | Decreased cell viability in a concentration-dependent manner with the IC50 value of 105.46 μM. | [17] | |
| Short-chain fatty acids | Sodium Butyrate | Breast | In vitro | MCF7 | MTT | Inhibited cell proliferation in a dose-dependent manner with the IC50 value of 1.26 mM. Induced morphological changes to the MCF7 cells, and cell cycle arrest in the G1 phase. | [18] |
| Sodium Butyrate | Breast | In vitro | MCF7 | Cell counting kit-8 (CCK-8) and Western blot | Inhibited MCF7 cell viability in a dose- and time-dependent manner, decreased B-cell lymphoma 2 (Bcl-2) protein expression, and induced morphological changes. | [19] | |
| Sodium Butyrate | Breast | In vitro | MCF7 and MB-MDA-468 | MTT and Annexin-V-FITC | Induced cytotoxicity and apoptosis in both breast cancer cell lines, and increased expression of 15-lipoxygenase type 1 (15-Lox-1) and production of 13-Hydroxyoctadecadienoic acid (13(S)HODE). | [20] | |
| Sodium Butyrate | Breast | In vitro | MCF7, T47-D, and MDA-MB-231 | MTT and sulforhodamine B (SRB) | Initiated epigenetic changes to acetylation of proteins; pyruvate kinase activity was increased in MDA-MB-231 cells and lactate dehydrogenase activity was increased in T47-D cells. Increased oxygen consumption in the MDA-MB-231 and T47-D cell lines. | [21] | |
| Sodium Butyrate | Breast | In vitro | MCF7 | CCK-8 | Inhibited cell proliferation in a dose- and time-dependent manner. Induced cell cycle arrest in the G1/G2 phase and a decrease in the S phase and caused chromatin relaxation. | [22] | |
| Butyrate | Breast | In vitro | MCF7 | Western blot and polymerase chain reaction (PCR) | Cell inhibition of 34% against MCF7 cells, increased histone H3K9 acetylation, and increased expression of p21waf1 and Retinoic acid receptor beta (RARβ). | [23] | |
| Sodium Butyrate | Breast | In vitro | SKBR3 | MTT | Combined treatment of NaB and trastuzumab demonstrated synergistic growth inhibition and elevated mRNA and protein levels of p27Kip1. | [24] | |
| Sodium Butyrate | Breast | In vitro | MRK-nu-1 | Western blot and caspase assay | Induction of caspase-3, -10, and -8, and formation of DNA fragmentation, in a dose- and time-dependent manner. Triggered apoptosis via the induction of caspase-10 activity. | [25] | |
| Sodium Butyrate | Breast | In vitro | MCF7 | MTT | Inhibited cell growth of MCF7 cells dose-dependently, induced cell cycle arrest in the G2/M phase, reduced p53 expression, decreased Bcl-2 mRNA and protein levels, increased apoptosis, and reduced glutathione levels. | [26] | |
| Sodium Butyrate | Breast | In vitro | MCF7 | Western blot and flow cytometry | Induced cell cycle arrest and apoptosis via interaction with p21waf1/cip1 with cyclin-dependent kinase (CDK) and decreased proliferating cell nuclear antigen (PCNA) levels. | [27] | |
| Sodium Butyrate | Breast | In vitro | MCF7, T47-D, and BT-20 | Western blot | Increased the expression of tumour necrosis factor receptor 1 (TNF-R1) and receptor 2 (-R2), TRAIL receptor 1 (TRAIL-R1) and receptor 2 (-R2), and Fas in MCF7 cells and acted synergistically with these receptors to inhibit cell proliferation and induced apoptosis via p21waf1 and its interaction with PCNA. | [28] | |
| Sodium Butyrate | Breast | In vitro | MCF7, MCF-7ras, T47-D, BT-20, and MDA-MB-231 | Western blot and PCR | Inhibited cell proliferation in all cell lines. Induced cell cycle arrest in the G2/M phase in MDA-MB-231 cells, and in the G1 phase for the other four cell lines. Inhibited cell growth in a p53-independent manner and induced apoptosis via the Fas/Fas L system. | [29] | |
| Sodium Butyrate | Breast | In vitro | MCF7 | MTT | Increased bioavailability when coupled with the hyaluronic acid drug delivery system due to the ability to bind to CD44 receptors, which are prominent on tumour surfaces. | [30] | |
| Sodium Butyrate | Breast | In vitro | MDA-MB-231 | Flow cytometry, Western blot, and protein array analysis | Induced cell cycle arrest in the G2 phase via the inhibition of histone H1 kinase activities, and increased levels of p21waf1. | [31] | |
| Sodium Butyrate | Breast | In vitro | MCF7, MDA-MB-231, T47-D, and BT-20 | Flow cytometry and Burton method to assess variation of DNA content | Inhibitory effect of 85-90% with a dose- and time-dependent inhibition of cell proliferation, induced cell cycle arrest in the G2/M phase, resulting in the induction of apoptosis in the estrogen receptor-positive cell lines MCF7 and T47-D. | [32] | |
| Sodium Butyrate | Breast | In vitro | MCF7 | Estrogen receptor assays | Initiated significant hyperacetylation of histones in MCF7 cells and lowered estrogen receptor levels. | [33] | |
| Sodium Butyrate | Breast | In vitro | MCF7 | CEA-Roche and Biorad protein assay | Induced morphological changes in MCF7 cells and reduced cell proliferation. | [34] | |
| Natural purine nucleoside | Inosine | Breast | In vitro | MCF7 and MDA-MB-231 | CyQuant XTT | Demonstrated primary cytoprotective activities during breast cancer hypoxia, rather than adenosine, which was previously thought to be the primary compound responsible for this bioactivity. | [35] |
| Metabolite Group | Metabolite | Cancer Type | Clinical Study Details | Clinical Observations | Reference |
|---|---|---|---|---|---|
| Short-chain fatty acids | Butyric acids, propionate, and acetate | Colorectal | A case-control study with 14 colorectal cancer (CRC) patients and 14 non-CRC subjects. | A decreasing abundance of SCFA-producing bacterium, e.g., Bifidobacterium, in CRC patients in comparison to non-CRC participants. The levels of all three SCFAs assessed were reduced in CRC patients, and the values for butyric acid and propionate were statistically significant. | [36] |
| Acetic, propionic, butyric, valeric, and plasma isovaleric acid | Solid cancer tumours | Prospective cohort biomarker study of 52 patients with solid cancer tumours that completed programmed cell death-1 inhibitors (PD-1i) therapy. | High concentrations of all SCFAs correlated with extended progression-free survival, and it was indicated that SCFA concentrations in stool samples may be associated with PD-1i efficacy. | [37] | |
| Butyrate and propionate | Breast | Conducted 16S rRNA gene sequencing, cell culture methods, and targeted metabolomics on faecal samples from premenopausal breast cancer patients and premenopausal healthy participants. | The abundance of SCFA-producing bacteria and enzymes was significantly reduced in premenopausal breast cancer patients in comparison to premenopausal healthy participants, and the overall composition of the gut microbiota differed substantially between the two groups. | [38] | |
| Bacteriocin | Azurin-p28 peptide | P53(+) metastatic solid tumours | NSC745104: Phase I human clinical trial of azurin-p28 in 15 patients (aged 47–80 years old) with p53(+) metastatic solid tumours | Participants did not exhibit an immune response to p28, significant adverse events, or dose-limiting toxicities. Indicative of a highly favourable therapeutic index for anticancer activity. | [39] |
| Azurin-p28 peptide | Central nervous system (CNS) tumours | NSC745104: Phase I human clinical trial on 18 children aged 3–21 years old with progressive or recurrent CNS tumours | The p28 peptide was well-tolerated in children with CNS tumours at the recommended adult phase II dose (4.16 mg/kg/dose), which correlated closely with the previous study on adult participants. The primary adverse event was grade 1 infusion-related reactions; however, these often did not require treatment and were short-lived. | [40] |
4. The Correlation between Gut Metabolites and Breast Cancer Development
4.1. The Microbiota of Healthy Breast Versus Breast Tumour Microenvironment
4.2. Microbial Dysbiosis and Breast Cancer Growth
5. Anticancer Action of Nisin against Breast Cancer
6. The Duality of Sodium Butyrate in Breast Cancer
The Anticancer Action of Butyrate against BC
7. Potential Implementation of Inosine in Breast Cancer Therapy
8. Gut Microbial Metabolites and Clinical Research in Breast Cancer
8.1. Clinical Studies Exploring the Association between Gut Metabolites and Cancer Development
8.2. Gut Metabolites and Standard Chemotherapies
9. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Jaye, K.; Li, C.G.; Bhuyan, D.J. The complex interplay of gut microbiota with the five most common cancer types: From carcinogenesis to therapeutics to prognoses. Crit. Rev. Oncol./Hematol. 2021, 165, 103429. [Google Scholar] [CrossRef] [PubMed]
- Mikó, E.; Kovács, T.; Sebő, É.; Tóth, J.; Csonka, T.; Ujlaki, G.; Sipos, A.; Szabó, J.; Méhes, G.; Bai, P. Microbiome-Microbial Metabolome-Cancer Cell Interactions in Breast Cancer-Familiar, but Unexplored. Cells 2019, 8, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xuan, C.Y.; Shamonki, J.M.; Chung, A.; DiNome, M.L.; Chung, M.; Sieling, P.A.; Lee, D.J. Microbial Dysbiosis Is Associated with Human Breast Cancer. PLoS ONE 2014, 9, e83744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ximenez, C.; Torres, J. Development of Microbiota in Infants and its Role in Maturation of Gut Mucosa and Immune System. Arch. Med. Res. 2017, 48, 666–680. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.L.; Xia, Y.L.; Sun, J. Breast and gut microbiome in health and cancer. Genes Dis. 2021, 8, 581–589. [Google Scholar] [CrossRef]
- Eslami-S, Z.; Majidzadeh-A, K.; Halvaei, S.; Babapirali, F.; Esmaeili, R. Microbiome and Breast Cancer: New Role for an Ancient Population. Front. Oncol. 2020, 10, 120. [Google Scholar] [CrossRef] [Green Version]
- Varsha, K.K.; Maheshwari, A.P.; Nampoothiri, K.M. Accomplishment of probiotics in human health pertaining to immunoregulation and disease control. Clin. Nutr. ESPEN 2021, 44, 26–37. [Google Scholar] [CrossRef]
- Ge, Y.; Wang, X.; Guo, Y.; Yan, J.; Abuduwaili, A.; Aximujiang, K.; Yan, J.; Wu, M. Gut microbiota influence tumor development and Alter interactions with the human immune system. J. Exp. Clin. Cancer Res. 2021, 40, 42. [Google Scholar] [CrossRef]
- Vergara, D.; Simeone, P.; Damato, M.; Maffia, M.; Lanuti, P.; Trerotola, M. The Cancer Microbiota: EMT and Inflammation as Shared Molecular Mechanisms Associated with Plasticity and Progression. J. Oncol. 2019, 2019, 1253727. [Google Scholar] [CrossRef]
- Parida, S.; Sharma, D. The power of small changes: Comprehensive analyses of microbial dysbiosis in breast cancer. Biochim. Biophys. Acta-Rev. Cancer 2019, 1871, 392–405. [Google Scholar] [CrossRef]
- Hill, D.A.; Artis, D. Intestinal bacteria and the regulation of immune cell homeostasis. Annu. Rev. Immunol. 2009, 28, 623–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaye, K.; Li, C.G.; Chang, D.; Bhuyan, D.J. The role of key gut microbial metabolites in the development and treatment of cancer. Gut Microbes 2022, 14, 2038865. [Google Scholar] [CrossRef] [PubMed]
- Tsvetikova, S.A.; Koshel, E.I. Microbiota and cancer: Host cellular mechanisms activated by gut microbial metabolites. Int. J. Med. Microbiol. 2020, 310, 151425. [Google Scholar] [CrossRef]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yang, H.R.; Wang, D.J.; Wang, X.X. Association between the gut microbiota and patient responses to cancer immune checkpoint inhibitors. Oncol. Lett. 2020, 20, 342. [Google Scholar] [CrossRef]
- Akbari, V.; Avand, A. In Vitro Cytotoxic Activity of a Lactococcus Lactis Antimicrobial Peptide against Breast Cancer Cells. Iran. J. Pharm. Sci. 2017, 13, 82–83. [Google Scholar]
- Paiva, A.D.; de Oliveira, M.D.; de Paula, S.O.; Baracat-Pereira, M.C.; Breukink, E.; Mantovani, H.C. Toxicity of bovicin HC5 against mammalian cell lines and the role of cholesterol in bacteriocin activity. Microbiology 2012, 158, 2851–2858. [Google Scholar] [CrossRef]
- Semaan, J.; El-Hakim, S.; Ibrahim, J.N.; Safi, R.; Einar, A.A.; El Boustany, C. Comparative effect of sodium butyrate and sodium propionate on proliferation, cell cycle and apoptosis in human breast cancer cells MCF-7. Breast Cancer 2020, 27, 696–705. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, P.-C.; Ma, Y.-B.; Fan, R.; Gao, F.-F.; Wei, L.; Zhang, J.-W. Sodium butyrate-induced apoptosis and ultrastructural changes in MCF-7 breast cancer cells. Ultrastruct. Pathol. 2016, 40, 200–204. [Google Scholar] [CrossRef]
- Salimi, V.; Shabani, M.; Tavakoli-Yaraki, M.; Nourbakhsh, M. Involvement of 15-lipoxygenase-1 in the regulation of breast cancer cell death induced by sodium butyrate. Cytotechnology 2016, 68, 2519–2528. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.F.; Carvalho, E.; Rumjanek, F.D.; Amoedo, N.D.; Pezzuto, P. Reciprocal modulation of histone deacetylase inhibitors sodium butyrate and trichostatin A on the energy metabolism of breast cancer cells. J. Cell. Biochem. 2015, 116, 797–808. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ma, L.; Li, L.; Sun, Y.; Wu, X.; Chen, L.; Wu, P. Histone deacetylase inhibitor sodium butyrate suppresses DNA double strand break repair induced by etoposide more effectively in MCF-7 cells than in HEK293 cells. BMC Biochem. 2015, 16, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrade, F.O.; Nagamine, M.K.; De Conti, A.; Chaible, L.M.; Fontelles, C.C.; Jordão, A.A., Jr.; Vannucchi, H.; Dagli, M.L.Z.; Bassoli, B.K.; Moreno, F.S.; et al. Efficacy of the dietary histone deacetylase inhibitor butyrate alone or in combination with vitamin A against proliferation of MCF-7 human breast cancer cells. Braz. J. Med. Biol. Res. 2012, 45, 841–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.W.; Wei, F.; Xu, J.; Wang, Y.C.; Chen, L.B.; Wang, J.H.; Guan, X.X. Trastuzumab enhances the anti-tumor effects of the histone deacetylase inhibitor sodium butyrate on a HER2-overexpressing breast cancer cell line. Int. J. Mol. Med. 2011, 28, 985–991. [Google Scholar] [CrossRef]
- Yokoyama, Y.; Kano, K.; Nohara, K. The important role of caspase-10 in sodium butyrate-induced apoptosis. Kobe J. Med. Sci. 2007, 53, 265–273. [Google Scholar]
- Louis, M.; Rosato, R.R.; Brault, L.; Osbild, S.; Battaglia, E.; Yang, X.-H.; Grant, S.; Bagrel, D. The histone deacetylase inhibitor sodium butyrate induces breast cancer cell apoptosis through diverse cytotoxic actions including glutathione depletion and oxidative stress. Int. J. Oncol. 2004, 25, 1701–1711. [Google Scholar] [CrossRef]
- Chopin, V.; Toillon, R.-A.; Le Bourhis, X.; Jouy, N. P21WAF1/CIP1 is dispensable for G1 arrest, but indispensable for apoptosis induced by sodium butyrate in MCF-7 breast cancer cells. Oncogene 2004, 23, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Chopin, V.; Hondermarck, H.; Le Bourhis, X.; Slomianny, C. Synergistic induction of apoptosis in breast cancer cells by cotreatment with butyrate and TNF-alpha, TRAIL, or anti-Fas agonist antibody involves enhancement of death receptors’ signaling and requires P21waf1. Exp. Cell Res. 2004, 298, 560–573. [Google Scholar] [CrossRef]
- Chopin, V.; Toillon, R.A.; Jouy, N.; Bourhis, X.L. Sodium butyrate induces P53-independent, Fas-mediated apoptosis in MCF-7 human breast cancer cells. Br. J. Pharmacol. 2002, 135, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Pellizzaro, C.; Daidone, M.G.; Miglierini, G.; Perbellini, A.; Coradini, D. Hyaluronic acid as drug delivery for sodium butyrate: Improvement of the anti-proliferative activity on a breast-cancer cell line. Int. J. Cancer 1999, 81, 411–416. [Google Scholar]
- Lallemand, F.; Courilleau, D.; Buquet-Fagot, C.; Atfi, A.; Montagne, M.-N.; Mester, J. Sodium butyrate induces G2 arrest in the human breast cancer cells MDA-MB-231 and renders them competent for DNA rereplication. Exp. Cell Res. 1999, 247, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Coradini, D.; Biffi, A.; Costa, A.; Pellizzaro, C.; Pirronello, E.; DiFronzo, G. Effect of sodium butyrate on human breast cancer cell lines. Cell Prolif. 1997, 30, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M.S.; Aliabadi, Z.; Moore, M.R. Associated Effects of Sodium-Butyrate on Histone Acetylation and Estrogen-Receptor in the Human-Breast Cancer Cell-Line MCF-7. Biochem. Biophys. Res. Commun. 1984, 119, 132–138. [Google Scholar] [CrossRef]
- Abe, M.; Kufe, D.W. Effect Of Sodium-Butyrate on Human-Breast Carcinoma (MCF-7) cellular proliferation, morphology, and cea production. Breast Cancer Res. Treat. 1984, 4, 269–274. [Google Scholar] [CrossRef]
- Smith, D.M. Inosine Production and Cytoprotective Activity in a Gradient Model of Breast Cancer Hypoxia Nutrient Exclusion (CHyNE). 2018. Available online: https://ttu-ir.tdl.org/handle/2346/86834 (accessed on 1 June 2022).
- Yusuf, F.; Adewiah, S.; Syam, A.F.; Fatchiyah, F. Altered profile of gut microbiota and the level short chain fatty acids in colorectal cancer patients. J. Phys. Conf. Ser. 2019, 1146, 012037. [Google Scholar] [CrossRef]
- Nomura, M.; Nagatomo, R.; Doi, K.; Shimizu, J.; Baba, K.; Saito, T.; Matsumoto, S.; Inoue, K.; Muto, M. Association of short-chain fatty acids in the gut microbiome with clinical response to treatment with nivolumab or pembrolizumab in patients with solid cancer tumors. JAMA Netw. Open 2020, 3, e202895. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Liu, Y.; Ye, S.; Yin, S.; Gu, J. Changes of intestinal microflora of breast cancer in premenopausal women. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 503–513. [Google Scholar] [CrossRef]
- Warso, M.; Richards, J.; Mehta, D.; Christov, K.; Schaeffer, C.; Rae Bressler, L.; Yamada, T.; Majumdar, D.; Kennedy, S.; Beattie, C. A first-in-class, first-in-human, phase I trial of p28, a non-HDM2-mediated peptide inhibitor of p53 ubiquitination in patients with advanced solid tumours. Br. J. Cancer 2013, 108, 1061–1070. [Google Scholar] [CrossRef]
- Lulla, R.R.; Goldman, S.; Yamada, T.; Beattie, C.W.; Bressler, L.; Pacini, M.; Pollack, I.F.; Fisher, P.G.; Packer, R.J.; Dunkel, I.J. Phase I trial of p28 (NSC745104), a non-HDM2-mediated peptide inhibitor of p53 ubiquitination in pediatric patients with recurrent or progressive central nervous system tumors: A Pediatric Brain Tumor Consortium Study. Neuro-Oncology 2016, 18, 1319–1325. [Google Scholar] [CrossRef] [Green Version]
- Alizadehmohajer, N.; Shojaeifar, S.; Nedaeinia, R.; Esparvarinha, M.; Mohammadi, F.; Ferns, G.A.; Ghayour-Mobarhan, M.; Manian, M.; Balouchi, A. Association between the microbiota and women’s cancers—Cause or consequences? Biomed. Pharmacother. 2020, 127, 110203. [Google Scholar] [CrossRef]
- Dworkin, A.M.; Huang, T.H.-M.; Toland, A.E. Epigenetic alterations in the breast: Implications for breast cancer detection, prognosis and treatment. Semin. Cancer Biol. 2009, 19, 165–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cauchi, J.P.; Camilleri, L.; Scerri, C. Environmental and lifestyle risk factors of breast cancer in Malta—A retrospective case-control study. EPMA J. 2016, 7, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarman, R.; Ribeiro-Milograna, S.; Kalle, W. Potential of the Microbiome as a Biomarker for Early Diagnosis and Prognosis of Breast Cancer. J. Breast Cancer 2020, 23, 579–587. [Google Scholar] [CrossRef]
- Yazdi, H.R.; Movafagh, A.; Fallah, F.; Alizadeh Shargh, S.; Mansouri, N.; Heidary Pour, A.; Hashemi, M. Evaluation of Methylobacterium radiotolerance and Sphyngomonas yanoikoaie in sentinel lymph nodes of breast cancer cases. Asian Pac. J. Cancer Prev. 2016, 17, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Sheflin, A.M.; Whitney, A.K.; Weir, T.L. Cancer-Promoting Effects of Microbial Dysbiosis. Curr. Oncol. Rep. 2014, 16, 406. [Google Scholar] [CrossRef] [Green Version]
- Fuhrman, B.J.; Feigelson, H.S.; Flores, R.; Gail, M.H.; Xu, X.; Ravel, J.; Goedert, J.J. Associations of the fecal microbiome with urinary estrogens and estrogen metabolites in postmenopausal women. J. Clin. Endocrinol. Metab. 2014, 99, 4632–4640. [Google Scholar] [CrossRef] [PubMed]
- Flores, R.; Shi, J.; Fuhrman, B.; Xu, X.; Veenstra, T.D.; Gail, M.H.; Gajer, P.; Ravel, J.; Goedert, J.J. Fecal microbial determinants of fecal and systemic estrogens and estrogen metabolites: A cross-sectional study. J. Transl. Med. 2012, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Goedert, J.J.; Jones, G.; Hua, X.; Xu, X.; Yu, G.; Flores, R.; Falk, R.T.; Gail, M.H.; Shi, J.; Ravel, J. Investigation of the association between the fecal microbiota and breast cancer in postmenopausal women: A population-based case-control pilot study. JNCI J. Natl. Cancer Inst. 2015, 107, djv147. [Google Scholar] [CrossRef]
- Goedert, J.J.; Hua, X.; Bielecka, A.; Okayasu, I.; Milne, G.L.; Jones, G.S.; Fujiwara, M.; Sinha, R.; Wan, Y.; Xu, X. Postmenopausal breast cancer and oestrogen associations with the IgA-coated and IgA-noncoated faecal microbiota. Br. J. Cancer 2018, 118, 471–479. [Google Scholar] [CrossRef] [Green Version]
- Fernández, M.F.; Reina-Pérez, I.; Astorga, J.M.; Rodríguez-Carrillo, A.; Plaza-Díaz, J.; Fontana, L. Breast cancer and its relationship with the microbiota. Int. J. Environ. Res. Public Health 2018, 15, 1747. [Google Scholar] [CrossRef] [Green Version]
- Baindara, P.; Mandal, S.M. Bacteria and bacterial anticancer agents as a promising alternative for cancer therapeutics. Biochimie 2020, 177, 164–189. [Google Scholar] [CrossRef] [PubMed]
- Yaghoubi, A.; Soleimanpour, S.; Khazaei, M.; Hasanian, S.M.; Avan, A.; Cho, W.C. Bacteriotherapy in breast cancer. Int. J. Mol. Sci. 2019, 20, 5880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, N.E.; Ritchie, K.; Kamarajan, P.; Miao, D.; Kapila, Y.L. Nisin, an apoptogenic bacteriocin and food preservative, attenuates HNSCC tumorigenesis via CHAC 1. Cancer Med. 2012, 1, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Begde, D.; Bundale, S.; Mashitha, P.; Rudra, J.; Nashikkar, N.; Upadhyay, A. Immunomodulatory efficacy of nisin—A bacterial lantibiotic peptide. J. Pept. Sci. 2011, 17, 438–444. [Google Scholar] [CrossRef]
- Rana, K.; Sharma, R.; Preet, S. Augmented therapeutic efficacy of 5-fluorouracil in conjunction with lantibiotic nisin against skin cancer. Biochem. Biophys. Res. Commun. 2019, 520, 551–559. [Google Scholar] [CrossRef]
- Rana, K.; Pandey, S.K.; Chauhan, S.; Preet, S. Anticancer therapeutic potential of 5-fluorouracil and nisin co-loaded chitosan coated silver nanoparticles against murine skin cancer. Int. J. Pharm. 2022, 620, 121744. [Google Scholar] [CrossRef]
- Mirzaei, R.; Afaghi, A.; Babakhani, S.; Sohrabi, M.R.; Hosseini-Fard, S.R.; Babolhavaeji, K.; Khani Ali Akbari, S.; Yousefimashouf, R.; Karampoor, S. Role of microbiota-derived short-chain fatty acids in cancer development and prevention. Biomed. Pharmacother. 2021, 139, 111619. [Google Scholar] [CrossRef]
- Jiang, W.J.; Guo, Q.Y.; Wu, J.; Guo, B.; Wang, Y.M.; Zhao, S.N.; Lou, H.L.; Yu, X.; Mei, X.Y.; Wu, C.Q.; et al. Dual effects of sodium butyrate on hepatocellular carcinoma cells. Mol. Biol. Rep. 2012, 39, 6235–6242. [Google Scholar] [CrossRef]
- Dominguez-Recio, M.E.; Jimenez-Rodriguez, B.; Lavado, R.; Comino-Mendez, I.; Alba, E.; Queipo-Ortuno, M.I.; Laborda-Illanes, A.; Sanchez-Alcoholado, L. Breast and gut microbiota action mechanisms in breast cancer pathogenesis and treatment. Cancers 2020, 12, 2465. [Google Scholar]
- Chen, J.Z.; Zhao, K.N.; Vitetta, L. Effects of Intestinal Microbial-Elaborated Butyrate on Oncogenic Signaling Pathways. Nutrients 2019, 11, 1026. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.-M.; Lee, M.; Lee, J.; Moon, H.-G.; Noh, D.-Y.; Han, W.; Kim, S.W. Enhanced anti-tumor activity and cytotoxic effect on cancer stem cell population of metformin-butyrate compared with metformin HCl in breast cancer. Oncotarget 2016, 7, 38500–38512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandal, M.; Kumar, R. BcI-2 expression regulates sodium butyrate-induced apoptosis in human MCF-7 breast cancer cells1. Cell Growth Differ. 1996, 7, 311–318. [Google Scholar] [PubMed]
- Yonezawa, T.; Kobayashi, Y.; Obara, Y. Short-chain fatty acids induce acute phosphorylation of the p38 mitogen-activated protein kinase/heat shock protein 27 pathway via GPR43 in the MCF-7 human breast cancer cell line. Cell. Signal. 2007, 19, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Tirandaz, H.; Mohammadi, E. Efficient tumor targeting by anaerobic butyrate-producing bacteria. Med. Hypotheses 2013, 80, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.J.; Jiang, W.; Xiao, C.C.; Yang, W.P.; Qin, Q.H.; Mao, A.Y.; Tan, Q.X.; Lian, B.; Wei, C.Y. Sodium Butyrate Combined with Docetaxel for the Treatment of Lung Adenocarcinoma A549 Cells by Targeting Gli1. Oncotargets Ther. 2020, 13, 8861–8875. [Google Scholar] [CrossRef]
- Huang, G.; Huang, H. Application of hyaluronic acid as carriers in drug delivery. Drug Deliv. 2018, 25, 766–772. [Google Scholar] [CrossRef]
- Lajkó, E.; Spring, S.; Hegedüs, R.; Biri-Kovács, B.; Ingebrandt, S.; Mező, G.; Kőhidai, L. Comparative cell biological study of in vitro antitumor and antimetastatic activity on melanoma cells of GnRH-III-containing conjugates modified with short-chain fatty acids. Beilstein J. Org. Chem. 2018, 14, 2495–2509. [Google Scholar] [CrossRef] [Green Version]
- Geng, H.-W.; Yin, F.-Y.; Zhang, Z.-F.; Gong, X.; Yang, Y. Butyrate suppresses glucose metabolism of colorectal cancer cells via GPR109a-AKT signaling pathway and enhances chemotherapy. Front. Mol. Biosci. 2021, 8, 112. [Google Scholar] [CrossRef]
- Chang, P.V.; Hao, L.; Offermanns, S.; Medzhitov, R. The microbial metabolite butyrate regulates intestinal macrophage function via histone deacetylase inhibition. Proc. Natl. Acad. Sci. USA 2014, 111, 2247–2252. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.R.; Kim, S.H.; Choi, H.G.; Jeon, C.K.; Lim, S.-J. Butyrate-mediated acquisition of chemoresistance by human colon cancer cells. Oncol. Rep. 2016, 36, 1119–1126. [Google Scholar] [CrossRef] [Green Version]
- Soares, A.S.; Costa, V.M.; Diniz, C.; Fresco, P. Inosine Strongly Enhances Proliferation of Human C32 Melanoma Cells through PLC-PKC-MEK1/2-ERK1/2 and PI3K Pathways. Basic Clin. Pharmacol. Toxicol. 2015, 116, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.H.; Wang, C.Y.; Huang, X.; Yi, H.K.; Li, Z.L.; Ma, K.L.; Ye, Y.S.; Zhang, J.W. Inosine induces acute hyperuricaemia in rhesus monkey (Macaca mulatta) as a potential disease animal model. Pharm. Biol. 2021, 59, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Mager, L.F.; Burkhard, R.; Pett, N.; Cooke, N.C.; Brown, K.; Ramay, H.; Paik, S.; Stagg, J.; Groves, R.A.; Gallo, M. Microbiome-derived inosine modulates response to checkpoint inhibitor immunotherapy. Science 2020, 369, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Aarnoutse, R.; Ziemons, J.; Penders, J.; Rensen, S.S.; De Vos-Geelen, J.; Smidt, M.L. The clinical link between human intestinal microbiota and systemic cancer therapy. Int. J. Mol. Sci. 2019, 20, 4145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plaza-Díaz, J.; Álvarez-Mercado, A.I.; Ruiz-Marín, C.M.; Reina-Pérez, I.; Pérez-Alonso, A.J.; Sánchez-Andujar, M.B.; Torné, P.; Gallart-Aragón, T.; Sánchez-Barrón, M.T.; Lartategui, S.R. Association of breast and gut microbiota dysbiosis and the risk of breast cancer: A case-control clinical study. BMC Cancer 2019, 19, 495. [Google Scholar] [CrossRef] [Green Version]
- Goh, K.S.; Ng, Z.J.; Halim, M.; Oslan, S.N.; Oslan, S.N.H.; Tan, J.S. A Comprehensive Review on the Anticancer Potential of Bacteriocin: Preclinical and Clinical Studies. Int. J. Pept. Res. Ther. 2022, 28, 1–12. [Google Scholar] [CrossRef]
- Abuei, H.; Behbahani, A.B.; Dehbidi, G.R.; Pirouzfar, M.; Zare, F.; Farhadi, A. Construction, Expression, and Purification of p28 as a Cell-Penetrating Peptide with Anticancer Effects on Burkitt’s Lymphoma Cell Line. Shiraz E-Med. J. 2019, 20, e85190. [Google Scholar] [CrossRef] [Green Version]
- Soleimani, M.; Sadeghi, H.M.; Jahanian-Najafabadi, A. A Bi-functional targeted P28-NRC chimeric protein with enhanced cytotoxic effects on breast cancer cell lines. Iran. J. Pharm. Res. IJPR 2019, 18, 735. [Google Scholar]
- Soltani, S.; Hammami, R.; Cotter, P.D.; Rebuffat, S.; Said, L.B.; Gaudreau, H.; Bédard, F.; Biron, E.; Drider, D.; Fliss, I. Bacteriocins as a new generation of antimicrobials: Toxicity aspects and regulations. FEMS Microbiol. Rev. 2021, 45, fuaa039. [Google Scholar] [CrossRef]
- Singh, R.P.; Bashir, H.; Kumar, R. Emerging role of microbiota in immunomodulation and cancer immunotherapy. Semin. Cancer Biol. 2021, 70, 37–52. [Google Scholar] [CrossRef]
- Vareki, S.M.; Chanyi, R.M.; Abdur-Rashid, K.; Brennan, L.; Burton, J.P. Moving on from Metchnikoff: Thinking about microbiome therapeutics in cancer. Ecancermedicalscience 2018, 12, 867. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Feng, Z.-P.; Liu, C.-G.; Xin, H.-Y.; Zhang, Z.-W.; You, H.; Xin, H.-W. Gut microbiota homeostasis restoration may become a novel therapy for breast cancer. Investig. New Drugs 2021, 39, 871–878. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaye, K.; Chang, D.; Li, C.G.; Bhuyan, D.J. Gut Metabolites and Breast Cancer: The Continuum of Dysbiosis, Breast Cancer Risk, and Potential Breast Cancer Therapy. Int. J. Mol. Sci. 2022, 23, 9490. https://doi.org/10.3390/ijms23169490
Jaye K, Chang D, Li CG, Bhuyan DJ. Gut Metabolites and Breast Cancer: The Continuum of Dysbiosis, Breast Cancer Risk, and Potential Breast Cancer Therapy. International Journal of Molecular Sciences. 2022; 23(16):9490. https://doi.org/10.3390/ijms23169490
Chicago/Turabian StyleJaye, Kayla, Dennis Chang, Chun Guang Li, and Deep Jyoti Bhuyan. 2022. "Gut Metabolites and Breast Cancer: The Continuum of Dysbiosis, Breast Cancer Risk, and Potential Breast Cancer Therapy" International Journal of Molecular Sciences 23, no. 16: 9490. https://doi.org/10.3390/ijms23169490