
Activation of ADRB2/PKA Signaling Pathway Facilitates Lipid Synthesis in Meibocytes, and Beta-Blocker Glaucoma Drug Impedes PKA-Induced Lipid Synthesis by Inhibiting ADRB2



Abstract
:1. Introduction
2. Results
2.1. PKA Activation Induced Lipogenesis in Human Meibomian Gland Epithelial Cells (HMGECs)
2.2. Differentiated HMGECs Express Beta 2-Adrenergic Receptor
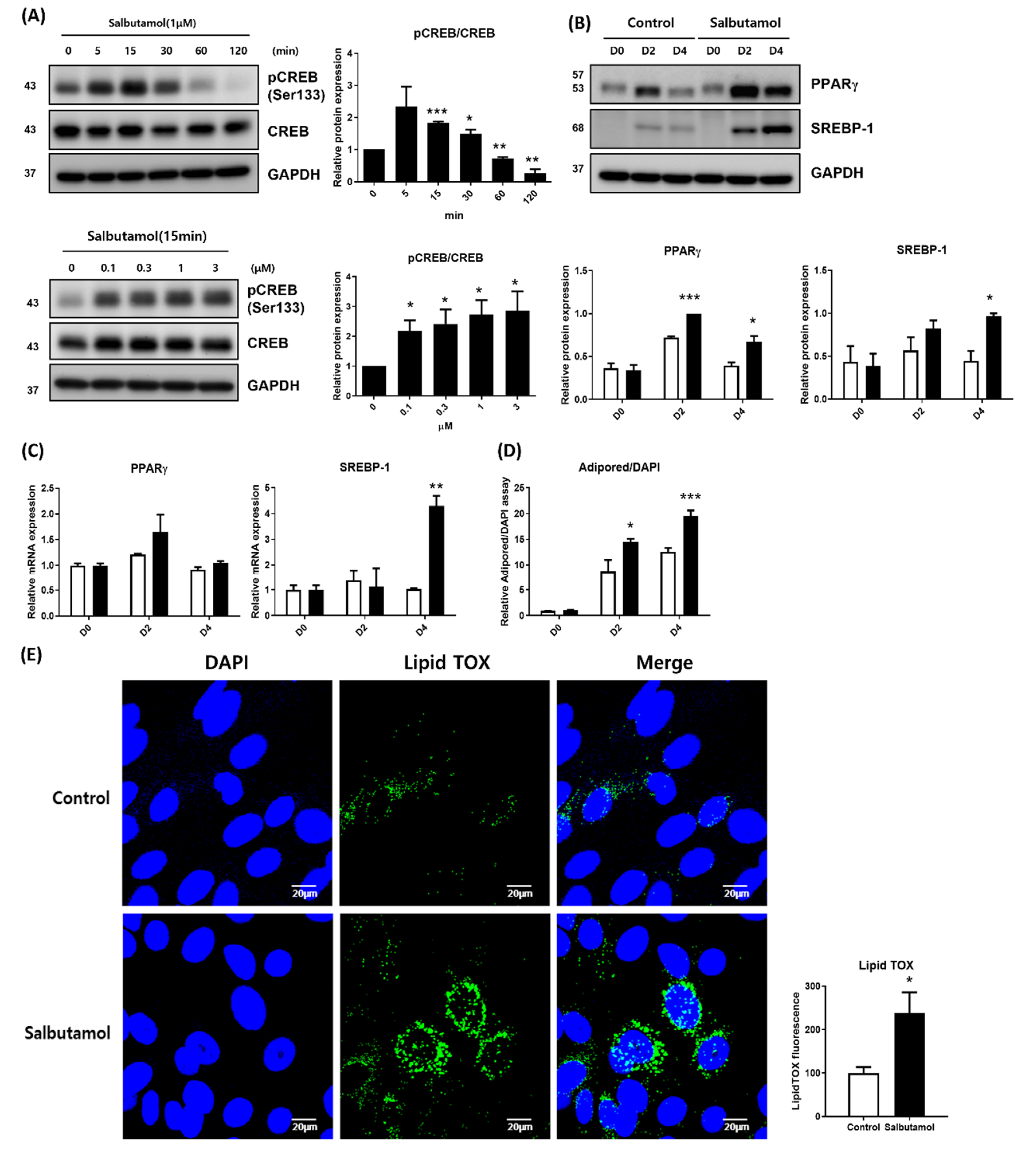
2.3. Beta 2-Adrenergic Agonist Facilitates PPARγ Expression and Lipid Droplet Synthesis
2.4. Beta 2-Adrenergic Antagonist Inhibits Lipogenesis in HMGECs
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Differentiation
4.2. Quantitative PCR Analysis
4.3. Immunoblot Analysis
4.4. Oil Red O Staining and AdipoRed Assay
4.5. Confocal Immunocytochemistry
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ADRB2 | Beta 2-adrenergic receptors |
| AKAP | PKA anchored proteins |
| cAMP | Cyclic adenosine monophosphate |
| CREB | cAMP-response element-binding protein |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| HMGEC | Human meibomian gland epithelial cell |
| MGD | Meibomian gland dysfunction |
| PPAR | Peroxisome proliferator activator receptor |
| PKA | Protein kinase A |
| SREBP | Sterol regulatory element-binding protein |
References
- Nichols, K.K.; Foulks, G.N.; Bron, A.J.; Glasgow, B.J.; Dogru, M.; Tsubota, K.; Lemp, M.A.; Sullivan, D.A. The international workshop on meibomian gland dysfunction: Executive summary. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1922–1929. [Google Scholar] [CrossRef] [Green Version]
- Chhadva, P.; Goldhardt, R.; Galor, A. Meibomian gland disease: The role of gland dysfunction in dry eye disease. Ophthalmology 2017, 124, S20–S26. [Google Scholar] [CrossRef]
- Ngo, W.; Gann, D.; Nichols, J.J. Impact of the 2011 international workshop on meibomian gland dysfunction on clinical trial attributes for meibomian gland dysfunction. Ocul. Surf. 2020, 18, 27–30. [Google Scholar] [CrossRef]
- Gidfar, S.; Afsharkhamseh, N.; Sanjari, S.; Djalilian, A.R. Notch signaling in meibomian gland epithelial cell differentiation. Investig. Ophthalmol. Vis. Sci. 2016, 57, 859–865. [Google Scholar] [CrossRef] [Green Version]
- Mauris, J.; Dieckow, J.; Schob, S.; Pulli, B.; Hatton, M.P.; Jeong, S.; Bauskar, A.; Gabison, E.; Nowak, R.; Argueso, P. Loss of cd147 results in impaired epithelial cell differentiation and malformation of the meibomian gland. Cell Death Dis. 2015, 6, e1726. [Google Scholar] [CrossRef] [Green Version]
- Qu, J.Y.; Xiao, Y.T.; Zhang, Y.Y.; Xie, H.T.; Zhang, M.C. Hedgehog signaling pathway regulates the proliferation and differentiation of rat meibomian gland epithelial cells. Investig. Ophthalmol. Vis. Sci. 2021, 62, 33. [Google Scholar] [CrossRef]
- Khandelwal, P.; Liu, S.; Sullivan, D.A. Androgen regulation of gene expression in human meibomian gland and conjunctival epithelial cells. Mol. Vis. 2012, 18, 1055–1067. [Google Scholar]
- Parfitt, G.J.; Brown, D.J.; Jester, J.V. Transcriptome analysis of aging mouse meibomian glands. Mol. Vis. 2016, 22, 518–527. [Google Scholar]
- Rosen, E.D.; Walkey, C.J.; Puigserver, P.; Spiegelman, B.M. Transcriptional regulation of adipogenesis. Genes Dev. 2000, 14, 1293–1307. [Google Scholar] [CrossRef]
- Yasmeen, R.; Jeyakumar, S.M.; Reichert, B.; Yang, F.; Ziouzenkova, O. The contribution of vitamin a to autocrine regulation of fat depots. Biochim. Biophys. Acta 2012, 1821, 190–197. [Google Scholar] [CrossRef] [Green Version]
- Jester, J.V.; Potma, E.; Brown, D.J. Ppargamma regulates mouse meibocyte differentiation and lipid synthesis. Ocul. Surf. 2016, 14, 484–494. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.W.; Xie, Y.; Nguyen, P.Q.; Bui, V.T.; Huynh, K.; Kang, J.S.; Brown, D.J.; Jester, J.V. Ppargamma regulates meibocyte differentiation and lipid synthesis of cultured human meibomian gland epithelial cells (hmgec). Ocul. Surf. 2018, 16, 463–469. [Google Scholar] [CrossRef]
- Jun, I.; Kim, B.R.; Park, S.Y.; Lee, H.; Kim, J.; Kim, E.K.; Seo, K.Y.; Kim, T.I. Interleukin-4 stimulates lipogenesis in meibocytes by activating the stat6/ppargamma signaling pathway. Ocul. Surf. 2020, 18, 575–582. [Google Scholar] [CrossRef]
- Nesterova, M.; Bossis, I.; Wen, F.; Horvath, A.; Matyakhina, L.; Stratakis, C.A. An immortalized human cell line bearing a prkar1a-inactivating mutation: Effects of overexpression of the wild-type allele and other protein kinase a subunits. J. Clin. Endocrinol. Metab. 2008, 93, 565–571. [Google Scholar] [CrossRef] [Green Version]
- Skalhegg, B.S.; Tasken, K. Specificity in the camp/pka signaling pathway. Differential expression, regulation, and subcellular localization of subunits of pka. Front. Biosci. 2000, 5, D678–D693. [Google Scholar]
- Rogne, M.; Tasken, K. Compartmentalization of camp signaling in adipogenesis, lipogenesis, and lipolysis. Horm. Metab. Res. 2014, 46, 833–840. [Google Scholar] [CrossRef]
- Shim, M.S.; Kim, K.Y.; Ju, W.K. Role of cyclic amp in the eye with glaucoma. BMB Rep. 2017, 50, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Ru, Y.; Huang, Y.; Liu, H.; Du, J.; Meng, Z.; Dou, Z.; Liu, X.; Wei, R.H.; Zhang, Y.; Zhao, S. Alpha-melanocyte-stimulating hormone ameliorates ocular surface dysfunctions and lesions in a scopolamine-induced dry eye model via pka-creb and mek-erk pathways. Sci. Rep. 2015, 5, 18619. [Google Scholar] [CrossRef] [Green Version]
- Arita, R.; Itoh, K.; Maeda, S.; Maeda, K.; Furuta, A.; Tomidokoro, A.; Aihara, M.; Amano, S. Effects of long-term topical anti-glaucoma medications on meibomian glands. Graefes Arch. Clin. Exp. Ophthalmol. 2012, 250, 1181–1185. [Google Scholar] [CrossRef]
- Uzunosmanoglu, E.; Mocan, M.C.; Kocabeyoglu, S.; Karakaya, J.; Irkec, M. Meibomian gland dysfunction in patients receiving long-term glaucoma medications. Cornea 2016, 35, 1112–1116. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, X.; Zhou, D.; Zhao, Y.; Duan, X. A narrative review of ocular surface disease related to anti-glaucomatous medications. Ophthalmol. Ther. 2022. [Google Scholar] [CrossRef]
- Beavo, J.A.; Brunton, L.L. Cyclic nucleotide research—Still expanding after half a century. Nat. Rev. Mol. Cell Biol. 2002, 3, 710–718. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Kam, W.R.; Liu, Y.; Chen, X.; Sullivan, D.A. Influence of pilocarpine and timolol on human meibomian gland epithelial cells. Cornea 2017, 36, 719–724. [Google Scholar] [CrossRef]
- Lee, V.H.; Luo, A.M.; Li, S.Y.; Podder, S.K.; Chang, J.S.; Ohdo, S.; Grass, G.M. Pharmacokinetic basis for nonadditivity of intraocular pressure lowering in timolol combinations. Investig. Ophthalmol. Vis. Sci. 1991, 32, 2948–2957. [Google Scholar]
- Zhang, H.; Kong, Q.; Wang, J.; Jiang, Y.; Hua, H. Complex roles of camp-pka-creb signaling in cancer. Exp. Hematol. Oncol. 2020, 9, 32. [Google Scholar] [CrossRef] [PubMed]
- Montminy, M.R.; Bilezikjian, L.M. Binding of a nuclear protein to the cyclic-amp response element of the somatostatin gene. Nature 1987, 328, 175–178. [Google Scholar] [CrossRef]
- Xiao, X.; Li, B.X.; Mitton, B.; Ikeda, A.; Sakamoto, K.M. Targeting creb for cancer therapy: Friend or foe. Curr. Cancer Drug Targets 2010, 10, 384–391. [Google Scholar] [CrossRef]
- Chin, K.V.; Yang, W.L.; Ravatn, R.; Kita, T.; Reitman, E.; Vettori, D.; Cvijic, M.E.; Shin, M.; Iacono, L. Reinventing the wheel of cyclic amp: Novel mechanisms of camp signaling. Ann. N. Y. Acad. Sci. 2002, 968, 49–64. [Google Scholar] [CrossRef]
- Fox, K.E.; Fankell, D.M.; Erickson, P.F.; Majka, S.M.; Crossno, J.T., Jr.; Klemm, D.J. Depletion of camp-response element-binding protein/atf1 inhibits adipogenic conversion of 3t3-l1 cells ectopically expressing ccaat/enhancer-binding protein (c/ebp) alpha, c/ebp beta, or ppar gamma 2. J. Biol. Chem. 2006, 281, 40341–40353. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.W.; Klemm, D.J.; Vinson, C.; Lane, M.D. Role of creb in transcriptional regulation of ccaat/enhancer-binding protein beta gene during adipogenesis. J. Biol. Chem. 2004, 279, 4471–4478. [Google Scholar] [CrossRef] [Green Version]
- Brasaemle, D.L.; Barber, T.; Kimmel, A.R.; Londos, C. Post-translational regulation of perilipin expression. Stabilization by stored intracellular neutral lipids. J. Biol. Chem. 1997, 272, 9378–9387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmen, G.Y.; Victor, S.M. Signalling mechanisms regulating lipolysis. Cell Signal 2006, 18, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Pidoux, G.; Witczak, O.; Jarnaess, E.; Myrvold, L.; Urlaub, H.; Stokka, A.J.; Kuntziger, T.; Tasken, K. Optic atrophy 1 is an a-kinase anchoring protein on lipid droplets that mediates adrenergic control of lipolysis. EMBO J. 2011, 30, 4371–4386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Hatton, M.P.; Khandelwal, P.; Sullivan, D.A. Culture, immortalization, and characterization of human meibomian gland epithelial cells. Investig. Ophthalmol. Vis. Sci. 2010, 51, 3993–4005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hampel, U.; Garreis, F. The human meibomian gland epithelial cell line as a model to study meibomian gland dysfunction. Exp. Eye Res. 2017, 163, 46–52. [Google Scholar] [CrossRef]
- Ishibashi, T.; Yokoi, N.; Kinoshita, S. Comparison of the effects of topical levobunolol and timolol solution on the human ocular surface. Cornea 2003, 22, 709–715. [Google Scholar] [CrossRef]
- Rolle, T.; Spinetta, R.; Nuzzi, R. Long term safety and tolerability of tafluprost 0.0015% vs timolol 0.1% preservative-free in ocular hypertensive and in primary open-angle glaucoma patients: A cross sectional study. BMC Ophthalmol. 2017, 17, 136. [Google Scholar] [CrossRef] [Green Version]
- Trope, G.E.; Liu, G.S.; Basu, P.K. Toxic effects of topically administered betagan, betoptic, and timoptic on regenerating corneal epithelium. J. Ocul. Pharmacol. 1988, 4, 359–366. [Google Scholar] [CrossRef]
- Nam, M.; Kim, S.W. Changes in corneal epithelial thickness induced by topical antiglaucoma medications. J. Clin. Med. 2021, 10, 3464. [Google Scholar] [CrossRef]
- Yuan, X.; Ma, X.; Yang, L.; Zhou, Q.; Li, Y. Β-blocker eye drops affect ocular surface through β2 adrenoceptor of corneal limbal stem cells. BMC Ophthalmol. 2021, 21, 419. [Google Scholar] [CrossRef]
- Ciancaglini, M.; Carpineto, P.; Agnifili, L.; Nubile, M.; Fasanella, V.; Lanzini, M.; Calienno, R.; Mastropasqua, L. An in vivo confocal microscopy and impression cytology analysis of preserved and unpreserved levobunolol-induced conjunctival changes. Eur. J. Ophthalmol. 2008, 18, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Kitazawa, K.; Cho, Y.; Yoshida, M.; Okumura, T.; Sato, A.; Kinoshita, S. Alteration in meibum lipid composition and subjective symptoms due to aging and meibomian gland dysfunction. Ocul. Surf. 2021. [Google Scholar] [CrossRef] [PubMed]
- Jun, I.; Kim, S.; Kim, H.; Kim, S.W.; Ji, Y.W.; Kim, K.P.; Lee, T.G.; Seo, K.Y. Evaluation of meibum lipid composition according to tear interferometric patterns: Rrh: Meibum composition according to interferometric patterns. Am. J. Ophthalmol. 2022, 240, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Jun, I.; Lee, J.S.; Lee, J.H.; Lee, C.S.; Choi, S.I.; Gee, H.Y.; Lee, M.G.; Kim, E.K. Adult-onset vitelliform macular dystrophy caused by best1 p.Ile38ser mutation is a mild form of best vitelliform macular dystrophy. Sci. Rep. 2017, 7, 9146. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′–3′) | Reverse (5′–3′) | Product Size (bp) |
|---|---|---|---|
| GAPDH | GAG TCA ACG GAT TTG GTC GT | GAC AAG CTT CCC GTT CTC AG | 185 |
| PPARγ | TTG CAG TGG GGA TGT CTC AT | TTT CCT GTC AAG ATC GCC CT | 208 |
| SREBP-1 | GAG CTC AAG GAT CTG GTG GT | CCG ACA CCA GAT CCT TCA GA | 175 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jun, I.; Choi, Y.J.; Kim, B.-R.; Seo, K.Y.; Kim, T.-i. Activation of ADRB2/PKA Signaling Pathway Facilitates Lipid Synthesis in Meibocytes, and Beta-Blocker Glaucoma Drug Impedes PKA-Induced Lipid Synthesis by Inhibiting ADRB2. Int. J. Mol. Sci. 2022, 23, 9478. https://doi.org/10.3390/ijms23169478
Jun I, Choi YJ, Kim B-R, Seo KY, Kim T-i. Activation of ADRB2/PKA Signaling Pathway Facilitates Lipid Synthesis in Meibocytes, and Beta-Blocker Glaucoma Drug Impedes PKA-Induced Lipid Synthesis by Inhibiting ADRB2. International Journal of Molecular Sciences. 2022; 23(16):9478. https://doi.org/10.3390/ijms23169478
Chicago/Turabian StyleJun, Ikhyun, Young Joon Choi, Bo-Rahm Kim, Kyoung Yul Seo, and Tae-im Kim. 2022. "Activation of ADRB2/PKA Signaling Pathway Facilitates Lipid Synthesis in Meibocytes, and Beta-Blocker Glaucoma Drug Impedes PKA-Induced Lipid Synthesis by Inhibiting ADRB2" International Journal of Molecular Sciences 23, no. 16: 9478. https://doi.org/10.3390/ijms23169478